
Abstract
Vasohibin-1 has been detected in endothelial cells as an intrinsic angiogenesis inhibitor. Both tumor-associated macrophages (TAMs) and transforming growth factor-β (TGF-β)/bone morphogenic protein (BMP) signaling have been reported to promote angiogenesis in cancer. However, whether vasohibin-1 expression is regulated by TGF-β/BMP signaling between TAMs and cancer cells remains unclear. The expression of TGF-β1, TGF-β2, BMP-4, and BMP-7 in TAMs and the expression of vasohibin-1, vascular endothelial growth factor-A (VEGF-A), and VEGF-C in two pancreatic cancer cell lines (a nonmetastatic cell line Panc-1 and a distant metastatic cell line HPAF-II) were measured by real-time reverse transcription-polymerase chain reaction (RT-PCR). The TGF-β receptor 1 and BMP receptor 1 were inhibited by the inhibitor SB-431542 and LDN193189, respectively. Thereafter, vasohibin-1, VEGF-A, and VEGF-C expression was detected by real-time RT-PCR. We found that the expression of TGF-β1, TGF-β2, BMP-4, and BMP-7 was upregulated in TAMs cocultured with pancreatic cancer cells. Vasohibin-1, VEGF-A, and VEGF-C mRNA expression in pancreatic cancer cells was upregulated by TAMs. Vasohibin-1 expression in pancreatic cancer cells cocultured with TAMs was upregulated significantly when TGF-β receptors or BMP receptors were inhibited, but VEGF-C expression was downregulated. Therefore, Vasohibin-1 expression is regulated by the TGF-β/BMP signaling between TAMs and pancreatic cancer cells. These results might shed a new light on the antiangiogenesis therapy in the pancreatic cancer.
Introduction
Chronic inflammation relates to tumor progression (Sgambato and Cittadini 2010). Vasohibin-1 expression has been demonstrated to be regulated by mediators of inflammatory process. Miyake and colleagues (2009) found that vasohibin-1 was expressed in rheumatoid arthritis synovial tissue of rheumatoid arthritis and that it was regulated by inflammatory cytokines. The stimulation of rheumatoid arthritis synovial fibroblasts with TNF-α and IL-1β for 8 h significantly downregulated vasohibin-1 mRNA expression under normoxic conditions. Tumor-associated macrophages (TAMs) are the main inflammatory cells in cancer, and they have emerged as key regulators in this process (Qian and Pollard 2010). However, whether vasohibin-1 expression in pancreatic cancer cells is induced by TAMs has been unknown.
The transforming growth factor-β (TGF-β) is the prototype of a large superfamily of secreted signaling polypeptides [including TGF-β, bone morphogenic proteins (BMPs), and activin/inhibin], with diverse functions in the development and pathogenesis of a variety of diseases (Piek and others 1999). The TGF-β/BMP family as a group of anti-inflammatory cytokines was reported to promote tumor metastasis in cancer (Jakowlew 2006). Overexpression of TGF-β1 in human prostate cancer cells significantly stimulates tumor growth and angiogenesis (Stearns and others 1999). Pertovaara and colleagues (1994) reported that TGF-β induces expression of VEGF, which directly acts on endothelial cells to stimulate cell proliferation and migration. In our preliminary study, we found that TGF-β1, TGF-β2, BMP-4, and BMP-7 expression was elevated in TAMs when cocultured with pancreatic cancer cells. However, the role of TGF-β/BMP signaling between TAMs and cancer cells needs to be clarified. So far, there were no reports about the relationship between the TGF-β/BMP signaling mediated by TAMs and vasohibin-1 expression in pancreatic cancer cells.
In this study, we used real-time reverse transcription-polymerase chain reaction (RT-PCR) to detect TGF-β1, TGF-β2, BMP-4, and BMP-7 expression in TAMs cocultured with pancreatic cancer cell lines and vasohibin-1, VEGF-A, and VEGF-C expression in Panc-1 and HPAF cell lines. We found that TGF-β1, TGF-β2, BMP-4, and BMP-7 expression was increased in TAMs cocultured with pancreatic cancer cells and that vasohibin-1, VEGF-A, and VEGF-C expression was increased by TAMs in pancreatic cancer cells. Moreover, we used the inhibitor SB-431542 and the inhibitor LDN193189 to block the TGF-β receptor 1 and BMP receptor 1, respectively, and found that vasohibin-1 expression was significantly upregulated in both inhibition groups in cancer cells, but the VEGF-C expression was downregulated.
Materials and Methods
Isolation of monocytes and macrophages
Mononuclear cells were isolated from the blood of healthy subjects with density gradient centrifugation (Ficol-Paque; Amersham). There were three layers of liquid after density gradient centrifugation, and mononuclear cells were in the second cloudy layer. This layer was transferred to a clean tube and centrifuged anew. The cells were washed with phosphate buffer saline (PBS)+10% acid citrate dextrose solution two times, and then counted, 1.4×106 cells were placed on Matrigel (BD Biosciences) covered coverslip (Nalge Nunc International Corporation). The isolated cells were grown in a serum-free medium designed for macrophages (a macrophage serum-free medium; Gibco) with the granulocyte-macrophage colony-stimulating factor (GM-CSF, 10 ng/mL; ImmunoTools), antibiotics, and 5% CO2 at 37°C. Monocytes adhered to the Matrigel overnight and differentiated into macrophages due to GM-CSF. Other nonadherent mononuclear cells were then removed from the medium. After 6 days, monocytes were fully differentiated into macrophages after 6 days and were then used for experiments. When cocultured with cancer cells, macrophages developed into TAMs with a special surface marker, CD14+ (Mantovani and others 2002). After coculturing with pancreatic cancer cells, the portion of CD14+ macrophages reached more than 80%, as measured with flow cytometry.
Cancer cell culture
The Panc-1 cell line (CRL-1469) was derived from fragments of a tumor resected from a pancreatic cancer patient. The HPAF-1 cell line (CRL-1997) was derived from the peritoneal ascitic fluid of a Caucasian male with primary pancreatic adenocarcinoma and metastases to the liver, diaphragm, and lymph nodes. The AsPc cell line (CRL-1682) was derived from nude mouse xenografts initiated with cells from the ascites of a patient with pancreatic cancer. The MiaPaCa (CRL-1420) cell line was derived from the tumor tissue of the pancreas. The Gastric cancer cell line Hs-746T was used as a reference in Western blot. The cell lines were purchased from American Type Culture Collection and cultured in the Dulbecco's modified Eagle's medium with 4 mM
Cell sorting by magnetic separation
Cell sorting was performed after pancreatic cancer cells were cultured with macrophages for 24 h. Cell sorting was processed by MACS separator (MACS Miltenyi Biotec), which is based on magnetic separation. After Matrigel-containing cells were dissolved in the cell recovery solution (Cell Recovery Solution; BD Biosciences; at 4°C) and centrifuged, 80 μL of a buffer (degassed PBS with 0.5% bull serum albumin and 2 mM ethylene diamine tetraacetic acid) and 20 μL CD11b MicroBeads (MACS Miltenyi Biotec) were added to the deposit and incubated for 20 min at 4°C. Then, the cells were washed with a buffer and centrifuged. Cells resuspended with a buffer were transferred to the prepared LS column (MACS Miltenyi Biotec). Effluents contained pancreatic cancer cells. The purity of pancreatic cancer cells was 97%, as measured with flow cytometry.
Inhibition of TGF-β receptor 1 or BMP receptor 1
SB-431542 (5 μM; Axon Medchem Company) or LDN193189 (400 nM; Axon Medchem Company) was added to the culture medium to inhibit the TGF-β receptor 1 or BMP receptor 1, respectively. The inhibitors were used for 24 h. After that, the cancer cells were separated by the MACS separator mentioned above. SB-431542 is a small molecule that acts as a specific and selective inhibitor of the TGF-β type 1 receptor, including activin receptor-like kinase (ALK) receptors 4, 5, and 7, but has no effect on BMP signaling. LDN193189 is highly potent small molecule BMP inhibitor inhibiting BMP type 1 receptors ALK2, ALK3, and ALK6.
Real-time RT-PCR
Total RNA was isolated from the sorted cells by membrane binding (Rneasy Mini Kit, 74104; Qiagen). RNA was reverse transcribed into single-stranded cDNA by the reverse transcriptase method (High Capacity cDNA Reverse Transcription Kit, 4368814; Applied Biosystems). The target gene expression was measured by the real-time RT-PCR method (TaqMan gene Expression Assay; Applied Biosystems). Glyceraldehyde-3-phosphate dehydrogenase (GAPDH) was used as endogenic control (TaqMan Endogenous Controls; Applied Biosystems). The PCRs were run in ABI PRISM 7000 sequence detection system. 2−ΔΔCt referred to the fold of the mRNA expression of one sample as compared to the calibration sample. 2−ΔCt was defined as the fold of mRNA expression of the target gene as compared to GAPDH expression in the same sample.
Western blotting
The cells were lysed by adding the radio immunoprecipitation assay lysing buffer (9806S; Cell Signaling Technology) (pH 7.6) with 1× Complete™ protease inhibitor cocktail (Roche) and 1 mM Pefabloc SC (Roche) and by incubating for 1 h at 0°C followed by short vortexing. Thereafter, the samples were concentrated with Microconr centrifugal filter devices (MRCPRT010; Merxk Millipore) by centrifuging at 14,000g for 20 min. Protein concentrations of the samples were determined by colorimetric Bradford assay and equal amounts of cytoplasmic protein extracts (30 μg) were diluted in the Laemmli sample buffer with 5% mercaptoethanol. After incubation for 5 min at 95°C, the samples were resolved in 10% polyacrylamide gels in the Tris-glycine-sodium dodecyl sulfate buffer. The gels were transferred to nitrocellulose membranes and blocked in the Odyssey blocking buffer (927-40000; LI-COR). The blots were then incubated overnight with mouse monocolonal anti-vasohibin-1 (sc-365541; Santa Cruz Biotechnology Inc.) and rabbit GAPDH antibodies (Santa Cruz Biotechnology, Inc.) in the Odyssey blocking buffer+Tween 0.1% (OBBT). The membranes were washed three times in PBS+0.1% Tween and incubated for 1 h at room temperature with goat anti-mouse IgG IRDye 800 [1:15000 (v/v); LI-COR] and with goat anti-rabbit IgG Alexa 680 dye (1:15000; Molecular Probes) in OBBT. After washing, protein bands in the membranes were scanned by the Odyssey infrared imager (LI-COR).
Statistical analysis
Statistical Package for Social Science version 17.0 was used. Data were expressed as means±standard error of the mean. The T-test was used to compare the mRNA expression between different groups. P-values were considered significant at the P<0.05 level.
Results
Vasohibin-1, VEGF-A, VEGF-C mRNA, and protein expression in pancreatic cancer cell lines
Vasohibin-1 and VEGF-C mRNA relative expression (×10−3) was significantly higher in the nonmetastatic cell line Panc-1 (9.0±0.1, 6.1±0.4) than in the metastatic cell line HPAF-II (1.0±0.0, 1.7±0.0; P<0.001, P=0.001). VEGF-A expression was higher in the HPAF-II cell line (45.2±5.9) than in the Panc-1 cell line (8.9±1.2, P=0.014, N=4). Weak vasohibin-1 expression was found in the nonmetastatic Panc-1 and MiaPaCa cell lines, while no expression was found in the metastatic cell lines HPAF and AsPc (Fig. 1).
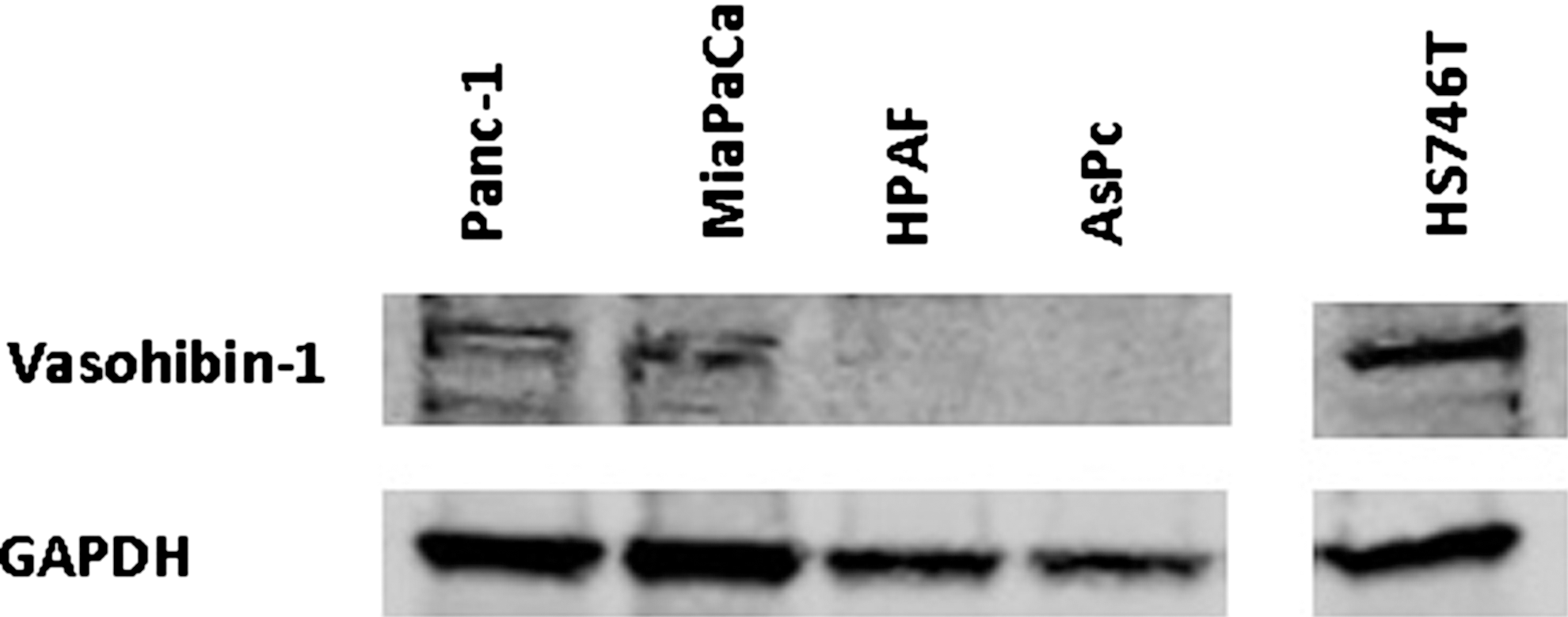
Vasohibin-1 protein level expression. A Western blot of vasohibin-1 protein expression in pancreatic cancer cells. Figure shows weak expression in the nonmetastatic Panc-1 and MiaPaCa cell lines, but no expression in the metastatic HPAF and AsPc cell lines. The Gastric cancer cell line HS745T was used as a reference.
Four cell lines (AsPc, HPAF, MiaPaCa, and Panc-1) were used to calculate the correlation between vasohibin-1 and VEGF-A and VEGF-C expression. There was a positive correlation between vasohibin-1 and VEGF-A expression (r=0.330, P=0.022), and between vasohibin-1 and VEGF-C expression (r=0.799, P<0.001).
Effect of TAMs on Vasohibin-1, VEGF-A, VEGF-C mRNA relative expression levels in pancreatic cancer cell lines
Vasohibin-1, VEGF-A and VEGF-C mRNA expression was all upregulated by TAMs. The upregulation of vasohibin-1 and VEGF-C was significantly higher compared with VEGF-A lower in the metastatic cell line as compared to the nonmetastatic cell line (Table 1).
Significant at the P<0.05 level (N=4).
VEGF, vascular endothelial growth factor.
Vasohibin-1, VEGF-A and VEGF-C expression in pancreatic cancer cells cocultured with TAMs after TGF-β receptor 1 or BMP receptor 1 inhibition
TGF-β1, TGF-β2, BMP-4, and BMP-7 expression was all upregulated in TAMs after coculturing with pancreatic cancer cells (Table 2). TGF-β1, TGF-β2, BMP-4, and BMP-7 expression levels [relative to GAPDH (×10−3) expression] in macrophages grown alone were 1.6±0.1, 33.5±0.3, 2.1±0.0, and 0.8±0.0, respectively.
Significant at the P<0.05 level (N=4).
TGF-β, transforming growth factor; BMP, bone morphogenic protein.
Vasohibin-1 expression was upregulated, but VEGF-C expression was significantly downregulated by both the TGF-β and BMP receptor inhibitors, as compared to the group without an inhibitor. There were no significant changes in VEGF-A expression (Table 3).
Significant at the P<0.05 level compared to the group without inhibitor (N=4).
Discussion
Angiogenesis is the critical step in metastasis of pancreatic cancer (Fisher and Berger 2003). The body contains not only proangiogenic factors, but also antiangiogenic factors, and the balance between those factors regulates the process of neovessel formation (Ribatti 2011). Vasohibin-1 secreted by endothelial cells has been found to play an inhibitory role in VEGF inducible angiogenesis and is considered as an intransitive angiogenesis inhibitor (Nasu and others 2009). The purified vasohibin-1 protein has been shown to inhibit the migration and the network formation of human umbilical vein endothelial cells in vitro (Hosaka and others 2009). Vasohibin-1 has demonstrated its antiangiogenic role in cancer tissues (Kerbel 2004), but most reports are related to its expression in cancer endothelial cells of cancer (Yoshinaga and others 2008; Sato and others 2009). Vasohibin-1 expression in cancer cells has been so far unknown. In this study, we found that not only vasohibin-1 was expressed in pancreatic cancer cells at both the mRNA and protein level, but also the level of its expression depended on the metastatic status of the cell. Vasohibin-1 expression was significantly lower in the distant metastatic cell line than in the nonmetastatic cell line, in contrary to the VEGF-A expression. This suggests that vasohibin-1 might play a more important role in antiangiogenesis in the nonmetastatic cell line than in the distant metastatic cell line.
VEGF-C expression is related to lymphangiogenesis and lympha node metastasis (Wissmann and Detmar 2006). The high VEGF-C expression found in the nonmetastatic cell line in this study might indicate that this cell line has a higher potential for lymphangiogenesis than the distant metastatic cell line. However, Heishi and colleagues (2010) found that vasohibin-1 inhibited the tumor lymphangiogenesis induced by VEGF-C. Accordingly, our nonmetastatic cell line had also a high vasohibin-1 expression probably balancing the lymphangiogenetic power of this cell line. However, this needs further transfectional research to be confirmed.
TAMs can exert protumor effects through the secretion of immunosuppressive cytokines and angiogenic cytokines (Malmberg 2004). Whether vasohibin-1 expression is induced by TAMs remains to be unknown. In this study, we found that vasohibin-1, VEGF-A and VEGF-C expression was all upregulated in both pancreatic cancer cell lines after coculturing with TAMs. This suggests that TAMs enhance not only the expression of proangiogenic factors (VEGF-A, VEGF-C), but also the expression of antiangiogenic factor (vasohibin-1). Moreover, we found that the upregulation of vasohibin-1 expression by TAMs was significantly higher and that of VEGF-A lower in the distant metastatic cancer cell line. It suggests that TAMs might play a more important role in promoting angiogenesis of the nonmetastatic pancreatic cancer cell line.
TGF-β and BMP subfamilies exert promoting effects on the tumor invasion and metastasis (Hong and others 2010). In this study, TGF-β1 and TGF-β2 and BMP-4 and BMP-7 expression in TAMs were all elevated after coculturing with pancreatic cancer cells. Thus, we speculated that the TGF-β/BMP signaling between TAMs and pancreatic cancer cells might play a positive role in promoting angiogenesis in combination with the upregulation of VEGF-A expression in pancreatic cancer cells induced by TAMs.
Vasohibin-1 is a potential agent in antiangiogenesis therapy. Hosaka and colleagues (2009) examined the function of vasohibin-1 in the mouse by subcutaneous inoculations of Lewis lung carcinoma cells. They found that tumors in vasohibin-1−/−mice contained more immature blood vessels and fewer apoptotic tumor cells than tumors in wild-type mice. In wild-type mice that had been inoculated with Lewis lung carcinoma cells, tail vein injection of adenovirus containing the human vasohibin-1 gene inhibited tumor growth and tumor angiogenesis. In our study, vasohibin-1 expression was upregulated in both the pancreatic cancer cell lines after TGF-β receptor 1 or BMP receptor 1 was blocked. This suggests that inhibition of the TGF-β or BMP inhibitor may suppress angiogenesis through increasing vasohibin-1 expression. However, further in vivo research is needed to confirm this.
In conclusion, vasohibin-1 was expressed in pancreatic cancer cells. Moreover, its expression was regulated by the TGF-β/BMP signaling between TAMs and pancreatic cancer cells. These results may be of importance when developing antiangiogenesis therapy for cancer.
Footnotes
Acknowledgments
This study was supported by grants from the Sigrid Juselius Foundation, Helsinki University Central Hospital Research Funds, and Peking University People's Hospital Funds (RDB 2012-18).
Author Disclosure Statement
No competing financial interests exist.