
Abstract
In chronic hepatitis C virus (HCV) infection, treatment failure and defective host immune response highly demand improved therapy strategies. Vγ9Vδ2 T-cells represent a good target for HCV immunotherapy, since phosphoantigen (PhAg)-activated Vγ9Vδ2 T-lymphocytes are able to inhibit subgenomic HCV replication by interferon (IFN)-γ release. A profound impairment of IFN-γ production by Vγ9Vδ2 T-cells during chronic HCV infection was previously shown. Interestingly, in vitro IFN-α partially restored Vγ9Vδ2 T-cells responsiveness to PhAg, by stabilizing IFN-γ-mRNA. To verify how in vivo IFN-α/ribavirin (RBV) treatment could affect Vγ9Vδ2 T-cells phenotype and responsiveness to PhAg in HCV-infected patients, 10 subjects underwent a longitudinal study before and after treatment. IFN-α/RBV therapy did not significantly modify Vγ9Vδ2 T-cell numbers and differentiation profile. Interestingly, Vγ9Vδ2 T-cell responsiveness remained unmodified until 3 weeks of therapy, but dropped after 1 month, suggesting that repeated in vivo IFN-α administration in the absence of T-cell receptor (TCR)-mediated signals results in Vγ9Vδ2 T-cell anergy. The present work defines the window of possible application of combined strategies targeting Vγ9Vδ2 T-cells during chronic HCV infection; specifically, the first 3 weeks from the beginning of treatment may represent the optimal time to target Vγ9Vδ2 T-cells in vivo, since their function in terms of IFN-γ production is preserved.
C
Standard therapy for HCV infection is based on a combination of pegylated interferon α (Peg-IFN-α) and ribavirin (RBV), and it is effective only in approximately half of the treated patients. Viral subtypes clearly differ in the treatment response: a sustained virological response (SVR) occurs in ∼80% of patients infected with HCV genotypes 2 or 3, and only in ∼45% of genotypes 1 or 4 (McHutchison and others 1998; Fried and others 2002; Jacobson and others 2007). Recently, a new and promising generation of direct-acting antiviral agents has been approved by the US Food and Drug Administration; the addition of these inhibitors, targeting HCV protease, to standard therapy has been found to substantially improve SVR rates (Jesudian and others 2012).
HCV persistence is mainly due to the failure of the host immune system to effectively and definitively clear the infection by generating a protective cellular immune response. Indeed, marked quantitative and qualitative defects of HCV-specific CD8 T-cells have been described in HCV patients (Neumann-Haefelin and Thimme 2011; Jouan and others 2012; Thimme and others 2012), correlated with innate immune cell impairment, such as in dendritic cells (Neumann-Haefelin and others 2005) and in natural killer cells (Par and others 2002; Oliviero and others 2009; Ahlenstiel and others 2010).
Among innate immune cells, Vγ9Vδ2 T-cells represent a good target for immunotherapy in infectious diseases (Gougeon and others 2002; Poccia and others 2009) for their multifaceted response capability (Poccia and others 2005a), able to exert direct antimicrobial effects (Qin and others 2009; Bonneville and others 2010) and to orchestrate the whole immune response (Scotet and others 2008; Agrati and others 2009; Eberl and others 2009; Nussbaumer and others 2011).
Notably, phosphoantigen (PhAg)-activated Vγ9Vδ2 T-lymphocytes were found able to inhibit subgenomic HCV replication, and this effect was mainly mediated by IFN-γ release (Agrati and others 2006), confirming the antiviral role of IFN-γ in inhibiting HCV replication. Nevertheless, as shown also for other chronic infections (Martini and others 2000), a decrease of Vγ9Vδ2 T-cells was observed during chronic HCV infection, associated to the acquisition of a terminally differentiated/effector phenotype (Par and others 2002). Moreover, a profound impairment of Vγ9Vδ2 T-cells during chronic HCV infection was also shown, resulting in a defective IFN-γ production (Cimini and others 2012). Thus, the identification of some costimulatory signal(s) able to improve Vγ9Vδ2 T-cell functions would allow to restore the Vγ9Vδ2 T-cell-mediated antiviral activity. Type-I IFN is known to be sensed by Vγ9Vδ2 T-cells by inducing an increase of CD69 expression (Kunzmann and others 2004). Interestingly, we previously showed that in vitro IFN-α stimulation was able to partially restore Vγ9Vδ2 T-cells responsiveness to PhAg in HCV-infected patients by stabilizing IFN-γ mRNA (Cimini and others 2012). The higher IFN-γ production by Vγ9Vδ2 T-cells after in vitro PhAg/IFN-α combination may represent an interesting approach to improve the antiviral activity of standard treatment. Nevertheless, data on in vivo effects of IFN-α administration on Vγ9Vδ2 T-cell activity are still missing.
The aim of this work was to verify whether in vivo IFN-α/RBV administration during standard treatment could restore Vγ9Vδ2 T-cells functionality in HCV-infected patients.
Ten chronically HCV-infected patients (HCV: 5 males and 5 females, mean age: 46.9±10.6 years) were enrolled at the INMI L. Spallanzani for a longitudinal study. This study was approved by the Ethics Committee of the Institute, and all individuals provided written informed consent. Patients' clinical features are described in Table 1. Standard therapy consisted on Peg-IFN-α-2a (180 μg/week, Pegasys; Schering-Plough) administered subcutaneously once weekly, and RBV (1 g/day, Copegus; Roche) administered orally.
SVR, sustained virological response; NR, nonresponder; GOT, glutamate oxaloacetate transaminase; GPT, glutamic pyruvic transaminase.
Plasma HCV-RNA levels were assayed by Abbott Real-Time HCV assay (Abbott Laboratories) and HCV genotype was determined by the Abbott RealTime HCV Genotype II Amplification Reagent kit (Abbott Laboratories). Peripheral blood mononuclear cells (PBMCs) from HCV-infected patients were isolated by lympholyte (Cedarlane; Horbay). Phenotype analysis of Vγ9Vδ2 T-cells was performed by using the following monoclonal antibodies: anti-Vδ2 FITC (clone IMMU389; Beckman Coulter Immunotech), anti-CD3 PerCP (clone SK7; BD Pharmingen), anti-CD27 APC (clone L128; BD Biosciences), and anti-CD45RA CY-Chrome (clone HI100; BD Biosciences). Briefly, PBMCs (1×106 cells/mL) were incubated with mAbs cocktail for 10 min at 4°C, washed twice, and fixed with paraformaldeyde (Sigma). Sample acquisition and data analysis were performed by a FACSCalibur flow cytometer (Becton Dickinson) by using Cell Quest software. Statistical analysis was performed by GraphPad Prism software by using the Wilcoxon test, and differences were considered significant when the P value was less than 0.05.
Figure 1A shows the HCV-RNA plasma levels of patients during the study: before starting HCV therapy (T0) and after 1 (T1w), 2 (T2w), 3 (T3w) weeks, and after 1 (T1m), 3 (T3m), 6 (T6m) months after treatment. As shown in Table 1, standard therapy was effective in 8 out of 10 patients in terms of an early virological response (EVR) and in 7 out of 10 patients in terms of SVR.
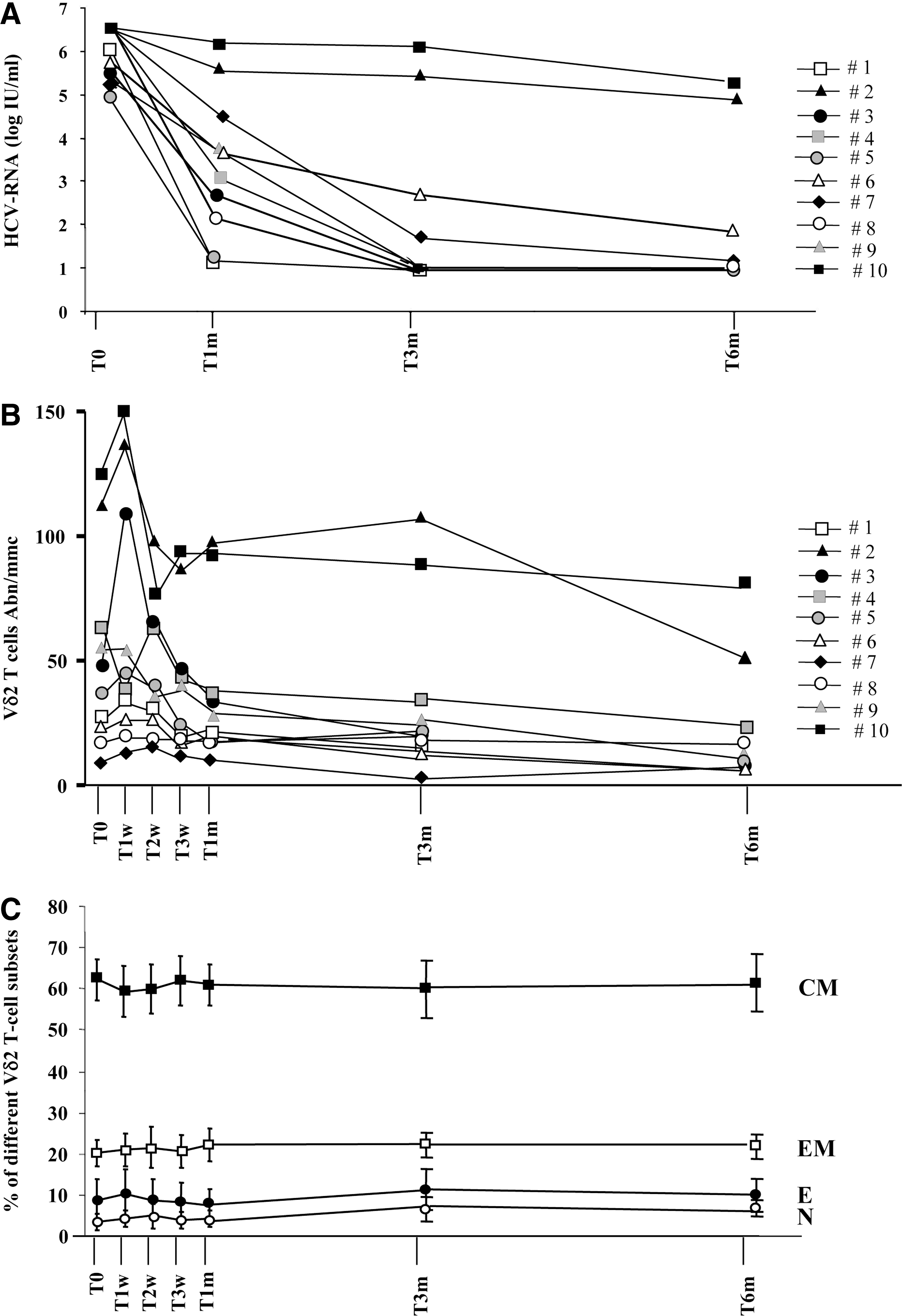
In vivo interferon (IFN)-α/ribavirin (RBV) effect on hepatitis C virus (HCV)-RNA and Vγ9Vδ2 T-cell phenotype.
Moreover, the effect of in vivo IFN-α/RBV treatment on Vγ9Vδ2 T-cells absolute number and phenotype were evaluated by flow cytometry at the same time points (Fig. 1B, C). Differentiation profile was defined by monitoring CD27 and CD45RA expression on the Vγ9Vδ2 T-cell surface. This approach is able to identify the frequency of naïve (CD27+CD45RA+), central memory (CD27+CD45RA−), effector memory (CD27−CD45RA−), and effector (CD27−CD45RA+) cells. Results showed a large variability in the Vγ9Vδ2 T-cell absolute number (Fig. 1B) and a transient increase was shown in some patients during the first week after treatment. Nevertheless, no significant differences were found, probably due to the small sample size. Similar results were obtained by analyzing the frequency of Vγ9Vδ2 T-cells (data not shown). Accordingly, Vγ9Vδ2 T-cell differentiation profile (Fig. 1C), were not significantly modified during IFN-α/RBV treatment. The EVR and SVR status were not correlated with Vγ9Vδ2 T-cells number and phenotype (data not shown).
To verify whether in vivo IFN-α/RBV administration could improve Vγ9Vδ2 T-cells response capability, a functional analysis was performed before and during standard treatment. Specifically, Vγ9Vδ2 T-cells were stimulated for 18 h with a synthetic PhAg (IPH1201, 200 nM; Innate-Pharma) known to specifically activate Vγ9Vδ2 T-cells (Tanaka and others 1995). IFN-γ production was evaluated by the ELISA test (Thermo Scientific). As shown in Fig. 2A, Vγ9Vδ2 T-cell responsiveness remained unmodified until 3 weeks of therapy, but dropped after 1 month (T1m), suggesting that after in vivo repeated IFN-α/RBV administrations, Vγ9Vδ2 T-cells lose their responsiveness to a subsequent in vitro PhAg stimulation. This observation represents a new important finding in designing innovative immunomodulating strategies targeting Vγ9Vδ2 T-cells by defining the small time window in which these cells are able to respond (T0–T3w). Nevertheless, a role of IFN-α in inducing Vγ9Vδ2 T-cell anergy seems in contrast with our recently published results about the adjuvant activity of IFN-α on Vγ9Vδ2 T-cell responsiveness in vitro (Cimini and others 2012). Thus, in vitro and in vivo effect of IFN-α on Vγ9Vδ2 T-cell reactivity was completely different. To explain this discrepancy, it should be considered that in this work, Vγ9Vδ2 T-cells received repeated in vivo IFN-α stimulations in the absence of PhAg. It is well known that T-cell responsiveness may be affected by repeated costimulation signals in the absence of a TCR-mediated antigenic signal (Lenschow and Bluestone 1993). We therefore suggest that the loss of Vγ9Vδ2 T-cell response to PhAg observed in our study can be at least partially due to a long time in vivo IFN-α/RBV costimulation without direct antigenic stimulation. To verify this hypothesis, PBMCs from HCV-infected patients during IFN-α/RBV therapy were stimulated in vitro with PhAg or with a combination of PhAg/rIFNα. We found that when used together with PhAg, IFN-α was able to boost IFN-γ production by Vγ9Vδ2 T-cells in all patients (Table 2), confirming previously published results (Cimini and others 2012). In Fig. 2B and C, 2 representative HCV-infected patients presenting different basal activation capabilities of Vγ9Vδ2 T-cells (PtA: 187.5 pg/1,000 Vδ2; PtB: 9.8 pg/1,000 Vδ2) are shown. Cytometric experiments performed on selected patients showed that IFN-γ produced after PhAg and PhAg/IFN-α stimulation was released only by Vγ9Vδ2 T-cells (data not shown). Thus, IFN-α may act as an adjuvant costimulation only in the presence of TCR-mediated stimulation. In contrast, when used in vivo without TCR-mediated stimulation, as during standard therapy, induced a reduction of Vγ9Vδ2 T-cell function.
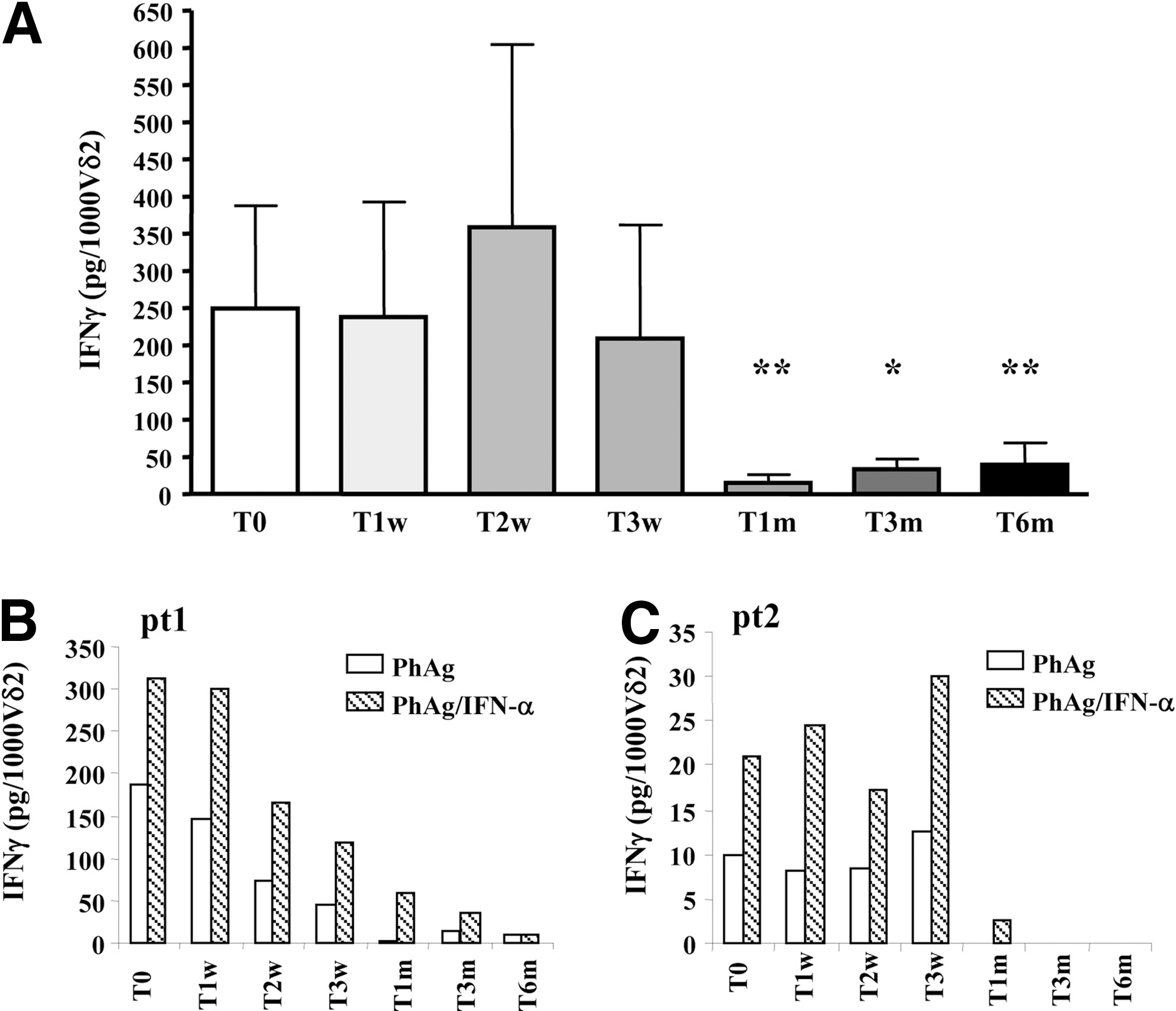
In vivo IFN-α/RBV treatment induces Vγ9Vδ2 T-cell anergy.
Results are expressed as adjuvant index obtained by dividing the amount of IFN-γ released after PhAg+IFN-α stimulation and those released after single PhAg stimulation. Results >1 represent a positive effect of IFN-α on Vγ9Vδ2 T-cell reactivity.
IFN, interferon; PhAg, phosphoantigen.
In recent years, the availability of new direct antiviral drugs for HCV infection does not exclude IFN-α/RBV use, to avoid the emergence of drug resistance (Reesink and others 2006; Sarrazin and Zeuzem 2010). Moreover, other new combined immunomodulating approaches may contribute to optimize the antiviral response by inducing antiviral cytokines, such as IFN-γ. In this context, Vγ9Vδ2 T-cells may represent a good target of immunomodulating strategies, for their ability to be easily activated in vivo by PhAgs (Sicard and others 2005; Bonneville and Scotet 2006; Dieli and others 2007; Poccia and others 2009) without HLA restriction (Morita and others 1995) and to orchestrate a complex network of antiviral and immunomodulating activities (Conti and others 2005; Poccia and others 2005b; Agrati and others 2009). We previously showed that IFN-α is able to improve Vγ9Vδ2 T-cell responsiveness in vitro in HCV patients by enhancing IFN-γ production (Cimini and others 2012). This fact, together with the finding that IFN-α can act synergistically with IFN-γ to inhibit HCV replication (Levy and others 1990; Larkin and others 2003; Jia and others 2007), strengthen the rational for testing combined standard antiviral and immunostimulating therapeutical strategies. The main result of this work was to demonstrate that in vivo IFN-α/RBV treatment was able to induce Vγ9Vδ2 T-cell anergy after 1 month from the beginning of therapy. We cannot rule out a possible direct involvement of RBV in inducing Vγ9Vδ2 T-cell anergy. Nevertheless, available data on RBV effect on the immune system show that it is able to polarize a Th1 immune response (Shiina and others 2004) suggesting a main role of IFN-α in this effect. Thus, the present work clearly defines the window of possible application of combined strategies targeting Vγ9Vδ2 T-cells during chronic HCV infection. Specifically, the first 3 weeks from the beginning of standard IFN-α/RBV treatment represent an optimal time to target Vγ9Vδ2 T-cells, since their function in terms of IFN-γ production is preserved. A future study on nonhuman HCV-infected primates treated with a combination of standard IFN-α/RBV therapy and PhAg during this window time is mandatory to verify both immunological and antiviral effectiveness of this strategy.
Footnotes
Acknowledgments
This work was supported by the Italian Ministry of Health (Current IRCCS Research Program). We thankfully dedicate this work to the memory of Fabrizio Poccia, a prominent researcher in the field of innate immunity and γδ T-cells, and an enthusiastic visionary on innate cell-based immunotherapy for infectious disease treatment, who died at the age of 39.
Author Disclosure Statement
No competing financial interests exist