
Abstract
Melanoma differentiation-associated gene-7/interleukin-24 (MDA-7/IL-24) uniquely takes on multiple anticancer functions, such as direct tumor cell cytotoxicity, immune stimulation, and antiangiogenic activities. MDA-7/IL-24 protein levels depend on proteasome degradation. Western blotting and coimmunoprecipitation analyses verified that the MDA-7/IL-24 protein was ubiquitinated and degraded by the 26S proteasome in Hela cells, which was confirmed by protein accumulation treated with proteasome inhibitor MG132. MDA-7/IL-24 contains 10 lysine sites: K63, K69, K78, K119, K123, K136, K179, K189, K203, and K206. Site-directed mutagenesis in these sites reveals that lysine 123 is the major internal lysine involvement of MDA-7/IL-24 ubiquitination. Our results further demonstrated that the mutation of lysine 123 to arginine led to the inhibition of ubiquitin-mediated degradation and the recovery of MDA-7/IL-24 protein level. At the same time, the apoptosis-inducing activity of K123R mutant was obviously stronger than wild-type MDA-7/IL-24 detected by 4′-6-diamidino-2-phenylindole (DAPI) assay and annexin V analysis. In addition, K123R mutant tilted the balance between antiapoptotic protein Bcl-2 and proapoptotic protein Bax, which subsequently induced the cleavage of caspase signaling cascade and initiated cell apoptosis. Together, lysine 123 is mainly implicated in the ubiquitination and degradation of MDA-7/IL-24. Inhibition of degradation and ubiquitination of MDA-7/IL-24 through mutation of lysine 123 result in enhanced stability of MDA-7/IL-24 and exhibits persistent tumor suppression activity compared with the wild type.
Introduction
T
Steady-state protein levels are regulated by not only ribosome synthesis but also proteasome degradation. Protein degradation speed has been primarily determined by either lysosomal destruction or ubiquitin-proteosome pathway. Recent studies demonstrated that proteasome inhibitor MG132 but not lysosome or protease inhibitors leads to MDA-7/IL-24 protein accumulation, which means that MDA-7/IL-24 is ubiquitinated and degraded by the 26S proteasome (Gopalan and others 2008). Ubiquitin is marked to target proteins through C-terminal carboxyl group of an Ub and ɛ-amino group of lysine residues within the substrate protein. This process is catalyzed by 3 enzymes: an ATP-dependent Ub activating enzyme (E1), an Ub-conjugating enzyme (E2), and an Ub ligase (E3) (Hershko and Ciechanover 1998). Since tumor suppressor proteins are involved with ubiquitination and degradation, inhibition of these proapoptotic factors' degradation through ubiquitin modification will be vital in anticancer strategies.
There are 10 lysine sites in the primary structure of MDA-7/IL-24: K63, K69, K78, K119, K123, K136, K179, K189, K203, and K206. Site-directed mutagenesis in these sites reveals that lysine 123 is the major internal lysine site participating in ubiquitin degradation of MDA-7/IL-24. K123R mutant remains higher level of protein compared with that of wild type. In addition, K123R mutant further takes part in the activation of cascade pathway through adjusting the Bcl-2/Bax balance from antiapoptosis to proapoptosis, which suggested that extension of MDA-7/IL-24 half-life via mutation of K123 facilitates cancer cell apoptosis. Consequently, the ubiquitination of MDA-7/IL-24 on lysine 123 may serve the function of signaling protein degradation by the proteasome and in regulation of the cancer-specific killing.
Materials and Methods
Cells and reagents
The human Henrietta Lacks (Hela) cell line was obtained from Shanghai Cell Collection. Cells were cultured in Dulbecco's modified Eagle's medium containing 10% fetal bovine serum, 2 mM
Preparation of the IL-24 mutant plasmids
The plasmid containing the full cDNA coding region of mda-7/IL-24 gene was a gift from Dr. Xin-Yuan Liu (Institute of Biochemistry and Cell Biology, Chinese Academy of Science, Shanghai, China). Mutant plasmids of mda-7/IL-24 in which K63, K69, K78, K119, K123, K136, K179, K189, K203, and K206 individually replaced with arginine were prepared using QuikChange mutagenesis via PCR according to manual. The primers for construction of these mutant enzymes were listed in Table 1. The resulting mutant genes were completely sequenced to be sure that no other mutations were present, and none was found.
Mutated codons were underlined.
Western blotting and immunoprecipitation
Western blotting analysis was performed as previously reported (Zheng and others 2005). Briefly, after transfected with the wild type or the different mutant MDA-7/IL-24 plasmids for the different time points, cells were collected by lysis buffer with cocktail protease inhibitor mixture for 20 min on ice. After centrifugation at 14,000 g for 15 min at 4°C, the supernatants were collected and determined for protein content using the Bradford method (Bio-Rad Laboratories). Proteins (80 μg) were separated by SDS-PAGE (10%) and transferred into nitrocellulose membranes. The membranes were blocked in 5% fat-free powder milk dissolved in phosphate-buffered saline (PBS) with 0.1% Tween20 for 2 h and sequentially were incubated with the primary antibody for overnight at 4°C. After incubation with alkaline phosphatase–conjugated goat anti-rabbit IgG or anti-mouse IgG, the membranes were developed using NBT/BCIP color substrate (Promega). For immunoprecipitation, cells (1×106) were seeded in 15-cm tissue culture dishes and treated with MG132. To reduce the affection of nonspecifically associated protein, cell lysates were then preincubated for 1 h with 20 μL protein A agarose beads and centrifuged at 14,000 g for 10 min at 4°C. The specific antibody (2 μg) was added to the supernatant and incubated with gentle rocking overnight at 4°C. After the addition of protein A agarose beads for 2 h, samples were triple washed with HEPES buffer and eluted by SDS-PAGE loading buffer and then heated at 100°C for 5 min. The complexes were separated on a 10% SDS-PAGE gel and subjected to Western blotting as mentioned previously.
Measurement of apoptosis by annexin V analysis
The annexin V–binding assay was conducted according to the recommendations of the manufacturer for apoptosis. In brief, the cells transfected with the wild type or the different mutant MDA-7/IL-24 plasmids were planted in 24-well plates; 48 h posttransfection, cells were treated twice with PBS and incubated with annexin V–FITC. Apoptotic cells bound with fluorochrome-labeled annexin V were randomly selected and visualized using fluorescent microscopy. The apoptotic rate is equal to the ratio of number of apoptotic cells to total number of cells.
4′-6-Diamidino-2-phenylindole staining
Nuclear condensation and fragmentation that stands for cell apoptosis were detected by 4′-6-diamidino-2-phenylindole (DAPI) staining. Hela cells were transfected with the wild type and the different mutant MDA-7/IL-24 plasmids for 48 h, and then fixed in 100 μL of 4% polyoxymethylene prior to washing with PBS. The washed cells were fixed and stained with 1 mg/mL DAPI for 15 min at room temperature in the dark. The cells were observed under the fluorescence microscope.
Cell viability assay
Cell survival rate was evaluated by a standard MTT assay [3-(4,5-dimethylthazol-2-yl)-2,5-diphenyltetrazolium bromide]. Hela cells (1.0×104 per well) were seeded in 96-well plates and transfected with MDA-7/IL-24 wild-type plasmid and 10 MDA-7/IL-24 mutant plasmids separately. Cells were incubated in 0.2 mL (final volume) for the different time points at 37°C. Then, 20 μL of MTT solution (5 mg/mL in PBS) was added to each well. About 150 μL of DMSO was added after 4 h of incubation at 37°C. The absorbance of this colored solution can be quantified by measuring at 570 nm wavelength by a spectrophotometer (Bio-Tek Instruments, Inc.). The percentage cell viability was calculated through dividing the number of viable cells by the total number of cells and multiplying by 100.
Statistical analysis
Values are expressed as the mean of 3 independent experiments and standard deviation. The Student's t-test or one-way analysis of the variance followed by the Duncan's new multiple range method or Newman–Keuls test was used to statistically compare the differences among multiple groups. P values<0.05 were considered significant.
Results
MDA-7/IL-24 is ubiquitinated and rapidly degraded by the 26S proteasome in situ
We first determined the expression of MDA-7/IL-24 protein by conducting Western blotting analysis at the different time points (Fig. 1A). Our data showed a marked reduction in MDA-7/IL-24. More than 30% of the protein level was decreased at 48 h after transfection, and MDA-7/IL-24 expression continued to decrease over time, with 65% reduction detected at 72 h compared with the protein level at 24 h. To investigate whether MDA-7/IL-24 degraded through ubiquitination, we analyzed MDA-7/IL-24 ubiquitination by immunoprecipitation (IP) (Fig. 1B). We found that the ubiquitination level was consistent with decrease of MDA-7/IL-24 protein expression, suggesting that the significant reduction of MDA-7/IL-24 was probably associated with ubiquitination modification. To confirm it, we further treated Hela cells with proteasome inhibitor MG132. The restoration of MDA-7/IL-24 protein expression by MG132 indicated that degradation of MDA-7/IL-24 was attributed to ubiquitin-mediated 26S proteasome pathway (Fig. 1C).
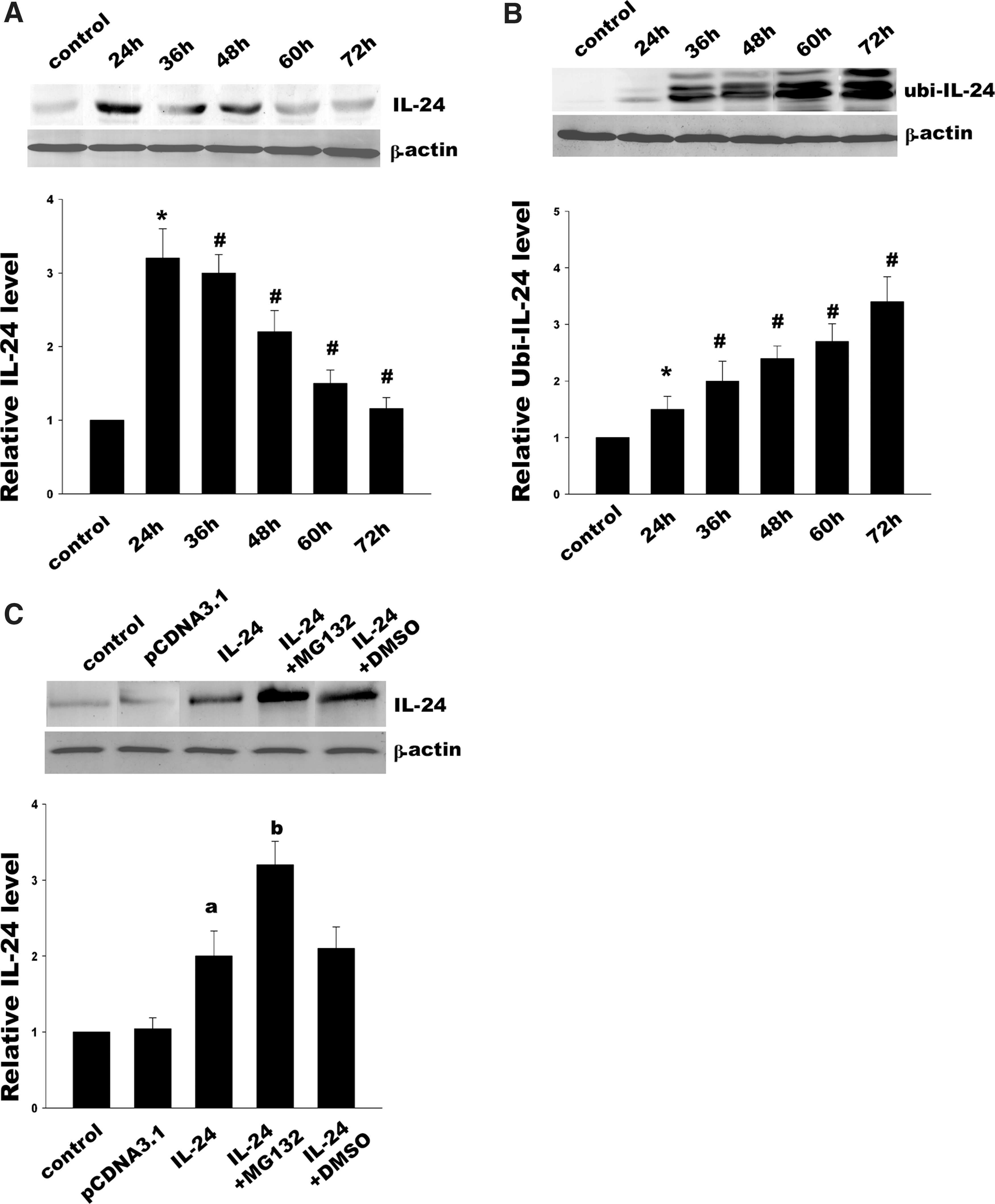
MDA-7/IL-24 expression and ubiquitin-dependent MDA-7/IL-24 degradation.
Implication of lysine 123 in ubiquitination of MDA-7/IL-24
MDA-7/IL-24 contains 10 lysine residues at positions 63, 69, 78, 119, 123, 136, 179, 189, 203, and 206. Site-directed mutagenesis was used to mutate these potential ubiquitination sites to arginine, respectively. Our results demonstrated that mutation of lysine at position 123 led to the accumulation of MDA-7/IL-24 protein, while other mutants had no effect, which means that lysine 123 was implicated in the ubiquitin-dependent degradation of MDA-7/IL-24 (Fig. 2A, B). At the same time, we further investigated ubiquitination of these mutants by IP. As suggested by the results shown in Fig. 2C and 2D, the replacement of the other 9 lysines did not affect ubiquitination of MDA-7/IL-24, except lysine 123. These data strongly suggested that lysine 123 was mainly involved with the ubiquitination and degradation of MDA-7/IL-24.
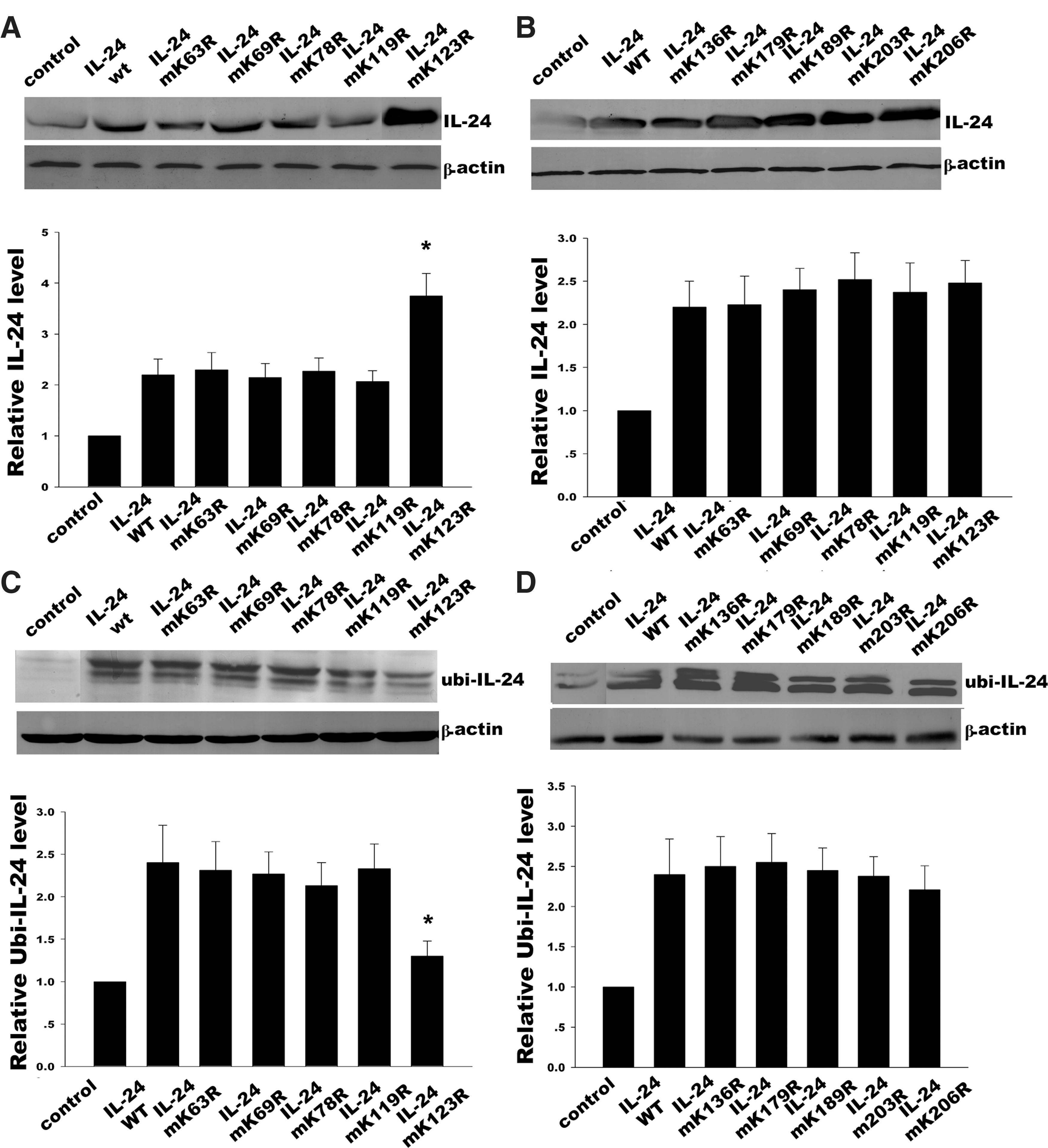
MDA-7/IL-24 mutant expression and ubiquitin-dependent degradation. Ten lysine residues in MDA-7/IL-24 were site-directed mutated to arginine and 10 MDA-7/IL-24 mutants were formed (K63R, K69R, K78R, K119R, K123R, K136R, K179R, K189R, K203R, and K206R).
Inhibition of MDA-7/IL-24 ubiquitination via K123R mutation enhances tumor cell killing
To detect whether MDA-7/IL-24 K123R mutant would affect the cancer cell apoptosis, Hela cell apoptosis was detected by annexin V and then analyzed using flow cytometry (Fig. 3). The results indicated that there were higher levels of annexin V–FITC-positive cells in MDA-7/IL-24 K123R mutant group compared with MDA-7/IL-24 wild-type group at 48 h after transfection. DAPI staining showed the nuclear condensation and fragmentation, which means MDA-7/IL-24 K123R mutant precipitated Hela cell apoptosis. On the other hand, the more significant cytopathic effect of MDA-7/IL-24 K123R mutant on Hela cells was induced in a time-dependent fashion compared with the wild-type IL-24, but the other 9 mutants showed similar growth inhibition to the wild type (Fig. 4).

Effects of MDA-7/IL-24 mutant (K123R) on Hela cell apoptosis.
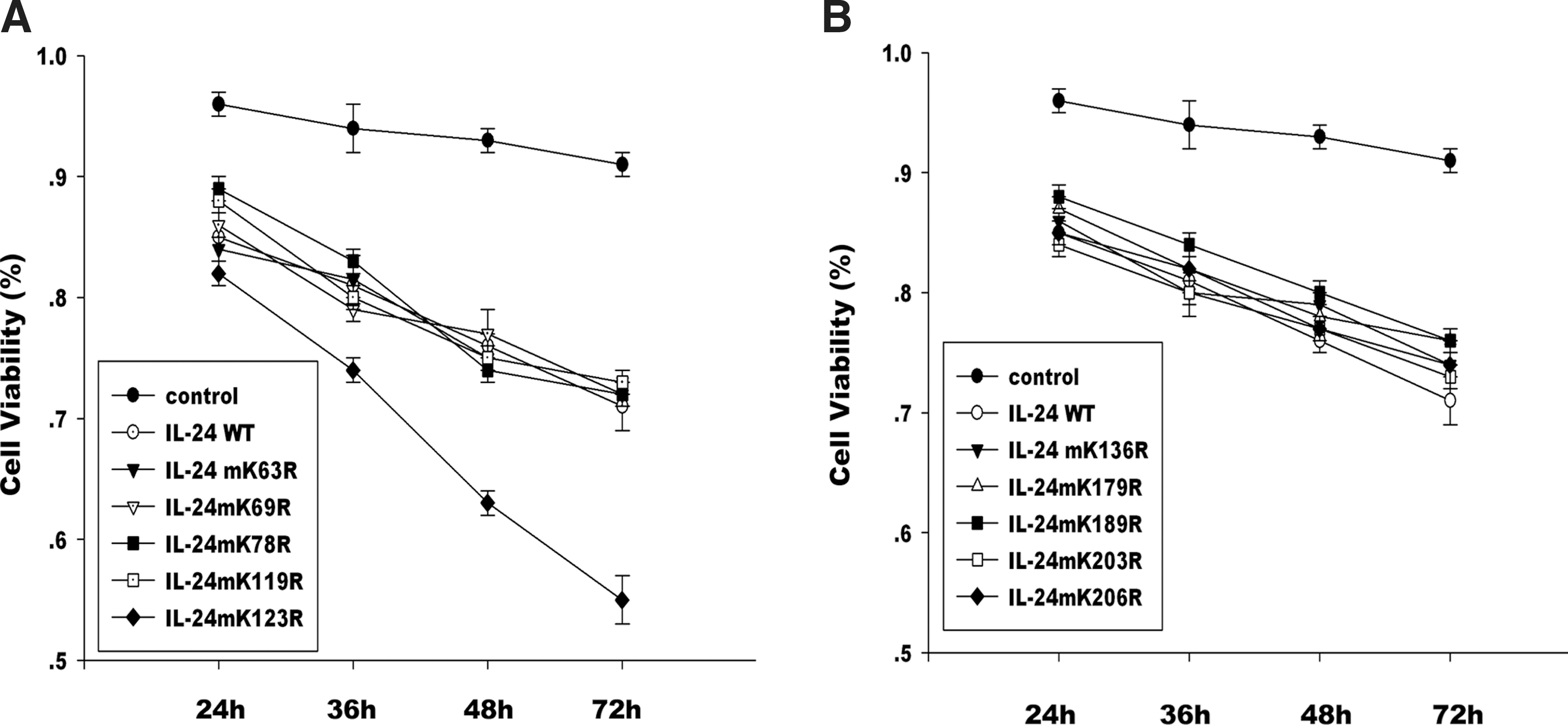
Effects of MDA-7/IL-24 mutants on Hela cell viability.
Inhibition of MDA-7/IL-24 ubiquitination via K123R mutation upregulates the activation of caspase signal pathway
The Bcl-2 family including both antiapoptotic proteins (eg, Bcl-2) and proapoptotic proteins (eg, Bax) plays an important role in MDA-7/IL-24-induced specific tumor apoptosis. The decreased ratio of Bcl-2 to Bax contributed to the activation of downstream caspase cascade. To further understand the underlying mechanism mediated by MDA-7/IL-24 K123R mutant, we analyzed the expression of proapoptotic gene Bax and antiapoptotic gene Bcl-2. Following transfection with MDA-7/IL-24 wild type and MDA-7/IL-24 K123R mutant, respectively (Fig. 5A), we detected that the latter exerted an increased expression of the tumor suppressor protein Bax and decreased expression of the apoptosis-inhibiting protein Bcl-2 compared with the MDA-7/IL-24 wild type. Moreover, MDA-7/IL-24 K123R mutant accelerated the cleavage of caspase-3 and PARP (Fig. 5B). These results suggested that mutation of lysine 123 did not impair the tumor-suppressive function of MDA-7/IL-24. On the contrary, MDA-7/IL-24 K123R mutant promoted the activation of caspase signal pathway through tilting the balance between Bcl-2 and Bax.
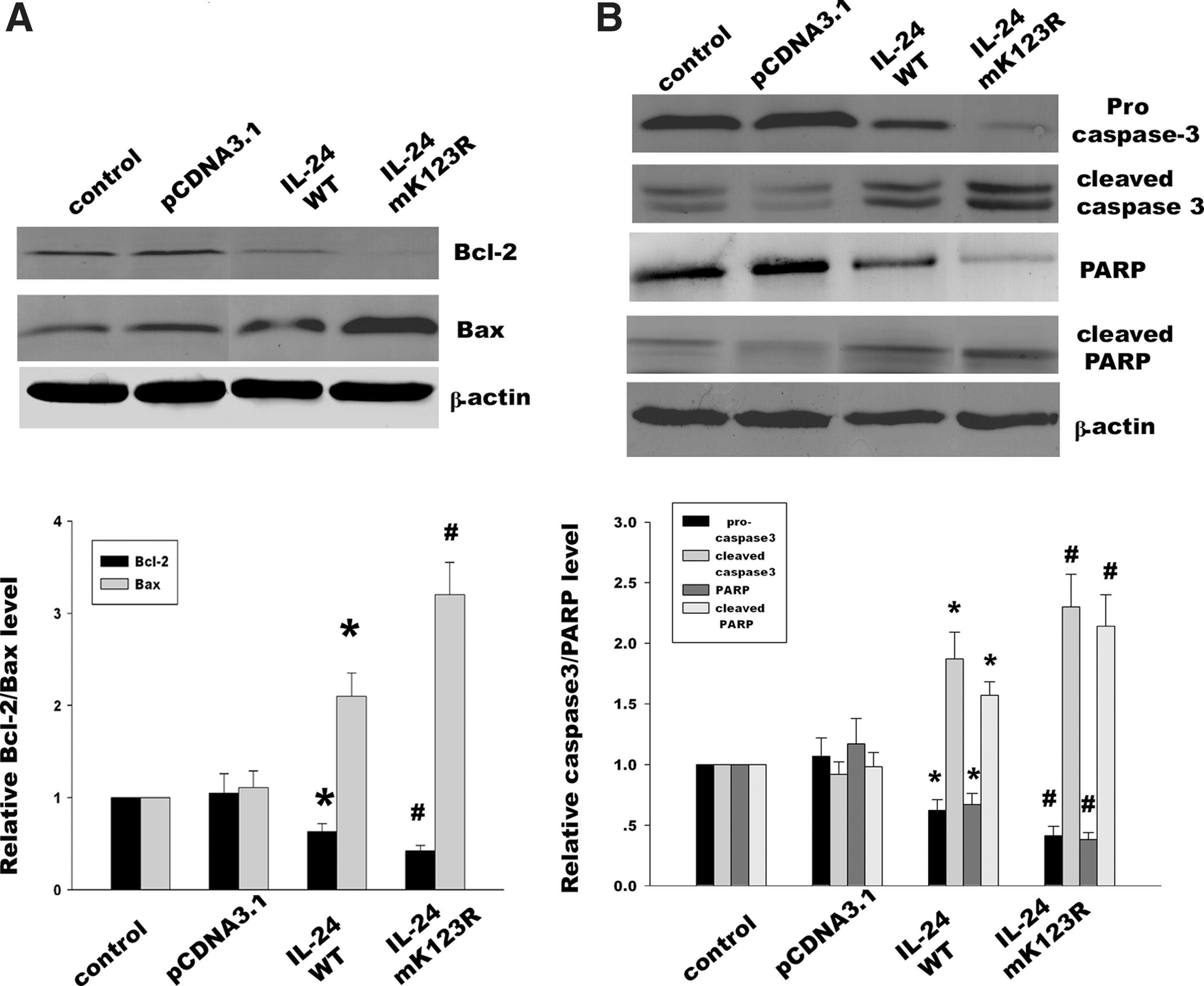
Effects of MDA-7/IL-24 K123R mutant on the ratio of Bcl-2 to Bax and the cleavage of caspase signal pathway.
Discussion
Ubiquitination and degradation of many proteins are essential in cell cycle progression, signal transduction, and cancer development (Lu and Hunter 2010; Muscolini and others 2011; Soond and Chantry 2011). The ubiquitination of the target protein is mediated by a cascade including an ubiquitin-activating enzyme (E1), an ubiquitin conjugating enzyme (E2), and an ubiquitin ligase (E3). Recent studies demonstrated that the proteasome inhibitor MG132 and small interfering RNA (siRNA)–mediated knockdown of ubiquitin resulted in MDA-7/IL-24 protein accumulation and increased tumor cell killing, which were further confirmed by using siRNA to inhibit the catalytic β-5 subunit of the 20S proteasome (Gopalan and others 2008). Therefore, MDA-7/IL-24 was shown to be ubiquitinated and then degraded by the proteasome pathway.
In the present study, we mutated 10 lysine sites in the primary structure of IL-24 to detect the therapeutic approach to inhibit protein ubiquitination and degradation. Using site-directed mutagenesis, we identified that lysine 123 was involved in the lysine-dependent ubiquitination. Substitution of lysine with arginine inhibited the ubiquitination and recovered MDA-7/IL-24 protein level. MDA-7/IL-24 leads to the modulation of the Bcl-2 family genes, and especially downregulates antiapoptotic Bcl-2 while upregulates proapoptotic Bax, which finally induces Hela cell apoptosis. To further detect whether the mutation of lysine 123 would affect the MDA-7/IL-24-mediated Hela cell apoptosis, we used annexin V and DAPI staining to detect the apoptotic rate of Hela cells. Our results demonstrated that the K123R mutant obviously aggravated Hela cell apoptosis compared with the MDA-7/IL-24 wild type. In addition, we found that overexpression of the MDA-7/IL-24 K123R mutant dramatically activated the common caspase cascades, including cleavage of caspase-3 and PARP. However, another challenge of MDA-7/IL-24 ubiquitination is to detect the ubiquitin ligase responsible for MDA-7/IL-24 ubiquitin modification. Future studies should focus on finding the ubiquitin ligase of MDA-7/IL-24 and detecting the regulation of the ubiquitin enzyme, which will further help us to understand the exact mechanism of the MDA-7/IL-24 protein degradation.
As one posttranslational modification, ubiquitin conjugation is always involved with regulation of protein stabilization (Li and others 2002; Gallagher and others 2006; Kessler and Edelmann 2011). Intriguingly, phosphorylation can precipitate ubiquitin-mediated proteolytic degradation, which means that inducible phosphorylation was easily distinguished by ubiquitin ligase E3 responsible for recognizing substrate protein to degradation (Chen and others 1995; Gallagher and others 2006). Some previous reports showed that Skp1-Cul1-F box protein (SCF) complexes as a class of multisubunit E3s specifically recognized phosphor amino acid moieties on the substrate and induced phosphorylated substrates to proteasomal degradation (Kipreos and Pagano 2000; Willems and others 2004). Consequently, whether MDA-7/IL-24 may take place the phosphorylation and this posttranslational modification would regulate ubiquitination through E3 ligase (eg, SCF complex) will be another study in the future.
Although numerous approaches have sought to regulation of MDA-7/IL-24 protein, it remains largely unknown. As a novel tumor suppressor/cytokine, MDA-7/IL-24 is degraded by the ubiquitin-proteasome pathway. Elucidating the exact site of ubiquitination is important, as ubiquitination frequently overlaps with other posttranslational modification. Our results demonstrate that lysine 123 is the major internal lysine of ubiquitination. Preventing ubiquitination at this site by mutation of lysine to arginine leads to stabilization of MDA-7/IL-24 protein and enhances specific tumor cell killing via downregulating the ratio of Bcl-2 to Bax and upregulating the activation of caspase signal pathway. The present study provides basis for future preclinical studies to inhibit ubiquitin-mediated degradation and promote stabilization of tumor-suppressor protein as an anticancer strategy.
Footnotes
Acknowledgments
This project was supported by the National Natural Science Foundation of China (No. 81071854), the Science and Technology Department of Jiangsu Province (Nos. BK2010177 and BK2010179), and Colleges and Universities in Jiangsu Province Natural Science Fund (No. 11KJB320017).
Author Disclosure Statement
The authors have declared that no competing interests exist.