
Abstract
Interferon (IFN)-γ is a potent stimulator of the IL-12 family Th1 cytokines, including IL-12/23p40 and IL-23, responsible for coordinating the innate and adaptive immune responses. Our results show that IFN-γ induced the production of IL-12/23p40 and IL-23p19 mRNA as well as IL-12p40 and IL-23 proteins in primary human monocytes isolated by positive selection through anti-CD14 microbeads. These results were confirmed by IFN-γ stimulation of CD14-activated monocytes resulting in IL-12/23p40 and IL-23 production. We investigated the signaling pathways governing the regulation of IL-23 and its subunits IL-23p40 and IL-23p19 following IFN-γ stimulation. We observed a differential regulation of IL-23p19, IL-12/23p40, and IL-23 following IFN-γ stimulation. IFN-γ-induced IL-23 and IL-12/23p40 expression was positively regulated by the p38 mitogen-activated protein kinases (MAPKs), independent of the Janus kinase (Jak)/signal transducers and activators of transcription (STAT) signaling. In contrast, IL-12 and IL-23 were negatively regulated by the Jak/STAT, phosphatidylinositol 3-kinase (PI3K), and the c-Jun-N-terminal kinase (JNK) MAPKs in IFN-γ-stimulated monocytes. Overall, our results suggest for the first time a differential positive regulation of IL-12p40 and IL-23 by p38 MAPKs independent of the Jak/STAT pathways and negative regulation by the Jak/STAT, JNK, and PI3K pathways in CD14-activated primary human monocytes stimulated with IFN-γ.
Introduction
Stimulation of various Toll-like receptors (TLRs) such as TLR2, TLR4, and TLR7/8 induce IL-23p19 expression or IL-23 production in DCs (Roses and others 2008). Moreover, IFN-γ and lipopolysaccharide (LPS) synergize to induce IL-12 and IL-23 expression in murine and human myeloid-derived DCs and monocytes/macrophages (Liu and others 2003; Gautier and others 2005; Roses and others 2008). CD14 is a glycoprotein that acts as a receptor for LPS either as an anchored protein in the cell membrane or in its soluble form in the serum. Soluble CD14 has been shown to activate monocytic cells in the absence of bacterial components or serum (Landmann and others 1998). Interestingly, IFN-γ has also been shown to negatively regulate LPS-induced IL-23 production in murine macrophages (Sheikh and others 2010). Whether IFN-γ alone can induce the production of IL-23, IL-12p40, or IL-23p19 in human monocytes remains unknown.
IFN-γ mediates its biological effects primarily via the activation of Janus kinase (Jak)/signal transducers and activators of the transcription (STAT) pathway (Gil and others 2001). IFN-γ has also been shown to activate mitogen-activated protein kinases [MAPKs; Valledor and others (2008a)], phosphatidylinositol 3-kinases [PI3Ks; Kaur and others (2008)], and Src kinases (Qing and Stark 2004) in a STAT1-independent manner (Gil and others 2001). The signaling pathways regulating IL-12p40 expression in LPS-stimulated monocytic cells have been identified (Murphy and others 1995; Plevy and others 1997; Becker and others 2001; Utsugi and others 2003; Maruyama and others 2003; Ma and others 2004). However, very little is known about the regulation of IL-23 (Utsugi and others 2006; Carmody and others 2007; Dobreva and others 2008; Roses and others 2008). LPS-induced IL-23p19 was negatively regulated by Rac1 through the NF-κBp65 transactivation-dependent and IκB-independent pathway in phorbol 12-myristate 13-acetate-treated THP-1 macrophages and human DCs without affecting IL-12/23p40 induction (Utsugi and others 2006). IL-23p19 expression was also regulated by c-rel in murine DCs stimulated with TLR-2, -3, -4, -6, and -9 agonists (Carmody and others 2007).
Our results show for the first time that IFN-γ induced the production of IL-12p40 and IL-23 in primary human monocytes isolated by positive selection through the use of anti-CD14 antibody-conjugated microbeads and not in negatively selected monocytes. Subsequently, we studied which signaling pathways are involved in the regulation of IL-12p40 and IL-23 in human monocytes following stimulation with IFN-γ. Our results suggest a differential positive regulation of IL-12p40 and IL-23 by p38 MAPKs, whereas these cytokines were negatively regulated by Jak/STAT, c-Jun N-terminal kinases (JNKs), and PI3K pathways in CD14-activated primary human monocytes in response to stimulation with IFN-γ.
Materials and Methods
Isolation of monocytes, cell culture, and reagents
Peripheral blood mononuclear cells (PBMCs) were isolated from the whole blood of healthy donors by density gradient centrifugation using Ficoll Hypaque (Amersham Biosciences). The monocytes were then isolated by positive selection using CD14 Micro Beads human (Miltenyi Biotec) by using the AutoMACS separator as per the manufacturer's directions. Monocytes were also negatively selected using the Monocyte Isolation Kit II (Miltenyi Biotec) (Saxena and others 2011). Untouched monocytes from PBMCs were isolated indirectly using magnetically labeled biotin-conjugated antibodies against CD3, CD7, CD16, CD56, CD123, and Glycophorin A and anti-biotin MicroBeads. Negatively selected monocytes were then cultured with magnetically labeled biotin-conjugated antibodies against CD14 (Miltenyi Biotec). The mononuclear cells obtained contained <5% CD3+ T cells and CD19+ B cells as determined by flow cytometric analysis. The purified monocytes were rested by keeping them on ice for 30 min and another 2 h at 37°C. Cells were cultured in Iscove's modified Dulbecco's medium (Sigma-Aldrich) supplemented with 10% fetal bovine serum (Invitrogen), 100 units/mL penicillin, 100 μg/mL gentamicin, 10 mm HEPES, and 2 mm glutamine. Monocytes (1×106/mL) were stimulated with IFN-γ (10 ng/mL) for 16 h. To determine the role of signaling pathways, monocytes (1×106/mL) were incubated with increasing concentrations of various pharmacological inhibitors for 2 h before stimulation with IFN-γ. The pharmacological inhibitors specific for Jak/STAT (Jak inhibitor I, 10–100 nM), extracellular signal-regulated kinase (ERK), (PD98059, 5–50 μM), p38 MAPK (SB203580, 5–50 μM), PI3K (LY294002, 5–50 μM) were purchased from Calbiochem-Merck, whereas inhibitors specific for JNK (SP600125, 5–50 μM) were purchased from Biomol and Sigma-Aldrich, respectively. Monocytes exposed to the signaling pathway inhibitors for 24 h were >95% viable by flow cytometry.
Measurement of IL-23p19 and IL-12/23p40 mRNA by relative quantitative real-time polymerase chain reaction
IL-23p19 and IL-12/23p40 mRNA expression were quantified by relative quantitative real-time polymerase chain reaction (RQ-PCR). Total RNA was extracted from human primary monocytes using the RNeasy kit (Qiagen Inc.). The cDNA was generated from 1 μg total RNA by using the Qiagen kit according to the manufacturer's instructions (Applied Biosystems). The expression level of each mRNA was quantitatively analyzed by a 7500 Real-Time PCR System (Applied Biosystems) with the cDNA prepared as described above and TaqMan Universal Master Mix (Applied Biosystems). The primers for IL-23p19 (HS00373324_m1), IL-12p40 (HS00233688_m1), and β-actin (HS99999903_m1) were purchased from Applied Biosystems. The expression levels of IL-23p19 and IL-12/23p40 mRNA quantified by the TaqMan Gene Expression Assay were shown as the ratio of that of β-actin, which was determined with the human β-actin control reagent (Applied Biosystems), by calculation of cycle threshold (Ct) values in amplification plots.
Quantification of IL-23 and IL-12/23p40 proteins by enzyme-linked immunosorbent assay
The IL-12/23p40 protein in the culture supernatants was quantified by enzyme-linked immunosorbent assay (ELISA) using two different monoclonal antibodies that recognize distinct epitopes as described previously (Ma and others 2004). ELISA plates (Thermo Fisher Scientific, Nunc) were coated overnight at 4°C with the anti-human IL-12p40 capture monoclonal antibody (Invitrogen; 1 μg/mL). IL-12/23p40 was detected using the biotinylated anti-IL-12/23p40 monoclonal antibody (Invitrogen; 350 ng/mL). Streptavidin-peroxidase (Jackson ImmunoResearch) was used at a final dilution of 1/1000. The color reaction was developed by o-phenylenediamine (Sigma-Aldrich) and hydrogen peroxide. Recombinant IL-12/23p40 (R&D Systems) was used as a standard. The IL-12p40 capture antibody measures all p40 dimerized with p19 as well as p35.
IL-23 production was quantified using a commercial ELISA kit (eBiosciences). ELISA plates (Thermo Fisher Scientific) were coated overnight at 4°C with the anti-human IL-23p19 capture antibody (eBiosciences) as per the manufacturer's instructions. IL-23 was detected using biotin-conjugated anti-human IL-12p40 (eBiosciences) antibodies. To determine the specificity of IL-23 ELISA, recombinant human IL-12p40 and IL-12p70 (R&D Systems) were used as standard negative controls, with no cross reactivity observed.
The color reaction was developed using the TMB Super Sensitive One Component HRP Microwell Substrate (SurModics In Vitro Diagnostic Products) by using horseradish peroxidase-conjugated avidin provided in the kit.
Cell surface staining of TLR4 and IFNGR1 by flow cytometry
Anti-human mouse antibodies used for flow cytometry were phycoerythrin (PE)-labeled monoclonal anti-IFN-γR1 and anti-TLR4 antibodies (R&D Systems). Monocytes (1×106 cells) were incubated with either anti-human TLR4-PE or anti-human IFNGR1-PE antibodies at 4°C for 30 min, and then washed with phosphate buffered saline as previously described (Gee and others 2002). The cell surface expression of IFN-γR1 and TLR4 was analyzed by flow cytometry using BD FACSCanto software (BD Biosciences) as described previously (Busca and others 2012). The results were expressed as percentage of cells expressing IFNGR1/TLR-4 receptors.
Western blot analysis
Cell lysates were collected and Jak, Akt, ERK, JNK, and MAPK phosphorylation was determined by Western blot analysis as previously described (Ma and others 2004). Cell lysates were prepared by treating the cell pellets at 4°C for 30–45 min with the lysis buffer [50 mM HEPES (pH 7.5), 150 mM sodium chloride, 10% glycerol, 1% Triton X-100, 1.5 mM magnesium chloride, 100 mM sodium fluoride, 100 mM sodium orthovanadate, and 1 mM ethylene glycol tetraacetic acid (pH 7.7)]. Total cell proteins (30 μg) were subjected to 10% polyacrylamide sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE) and the proteins were subsequently transferred onto polyvinylidene difluoride membranes (Bio-Rad). The membranes were probed with either rabbit anti-human-phospho-p38 (Thr180/Tyr182; New England Biolabs), mouse anti-human-phosho-ERK MAPKs (Thr202/Tyr204) (Santa Cruz Biotechnologies, Inc.), mouse anti-human phospho-JNK (Thr183/Tyr185; Santa Cruz Biotechnologies, Inc.), and rabbit anti-human-phospho-Akt (Ser473; Cell Signalling Technology) antibodies followed by detection with horseradish peroxidase-conjugated goat anti-rabbit or goat anti-mouse polyclonal antibodies (Bio-Rad). The membranes were stripped [stripping buffer: 62.5 mM Tris-HCl (pH 6.7), 100 mM 2-mercapto-ethanol, 2% SDS-PAGE, and 710 μL dithiothreitol] for 30 min and reprobed with rabbit polyclonal antibodies specific for the unphosphorylated forms of either ERK, p38, JNK MAPKs (Santa Cruz Biotechnologies, Inc.), or Akt (Cell Signalling Technology). All immunoblots were visualized by enhanced chemiluminescence (Amersham).
Neutralization assay of IFN-γ R1/CD119
Monocytes (1×106) were incubated with goat anti-human IFN-γR1/CD119 (10 μg/mL; IgG; R&D Systems) followed by IFN-γ stimulation. As a control, monocytes were also incubated with equal quantities of an isotype-matched goat IgG antibody (R&D Systems) in the presence of IFN-γ. Cells were cultured for 16 h, following which supernatants were analyzed for IL-23 production by ELISA as described above.
Statistical analysis
Means were compared using the two-tailed Student's t-test. Results are expressed as mean±standard error of mean (SEM).
Ethics statement
Informed consent was given in writing by all participants based on the study protocol, which was approved by the Research Ethics Boards of the Research Institute, Children's Hospital of Eastern Ontario and the Ottawa Hospital Research Institute, University of Ottawa, Ottawa, Ontario, Canada.
Results
IFN-γ induces the expression of IL-12p40 and IL-23 in positively selected, but not in negatively selected human monocytes
Since IFN-γ was shown to prime murine monocytic cells to produce IL-12 in response to LPS (Liu and others 2003), we determined whether monocytes isolated by negative or positive selection respond distinctly to normal physiological stimulants such as IFN-γ by producing IL-23 and its subunits. IFN-γ stimulation of positively selected monocytes induced the expression of IL-23p19 and IL-12/23p40 mRNA (Fig. 1A) as well as IL-23 and IL-12/23p40 proteins (Fig. 1B) compared to unstimulated cells. These positively selected monocytes secreted high basal levels of IL-12 and IL-23 proteins. In contrast, stimulation of negatively selected monocytes with IFN-γ failed to induce IL-12p40 and IL-23 proteins. At the same time, negatively selected monocytes produced relatively low basal levels of IL-12p40 and IL-23 proteins (Fig. 2A, B).
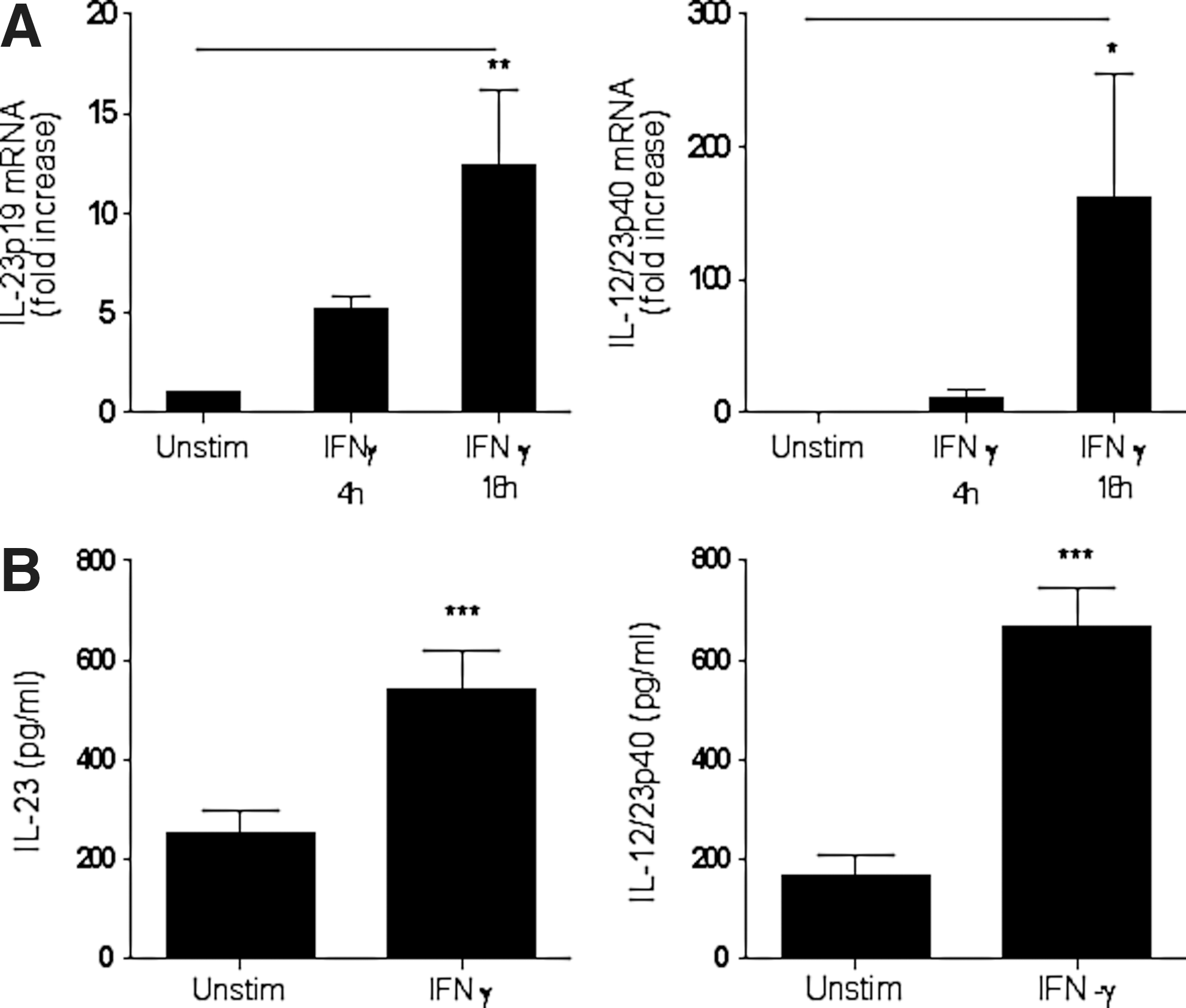
IFN-γ induces IL-23 and IL-12p40 expression in positively selected human monocytes.
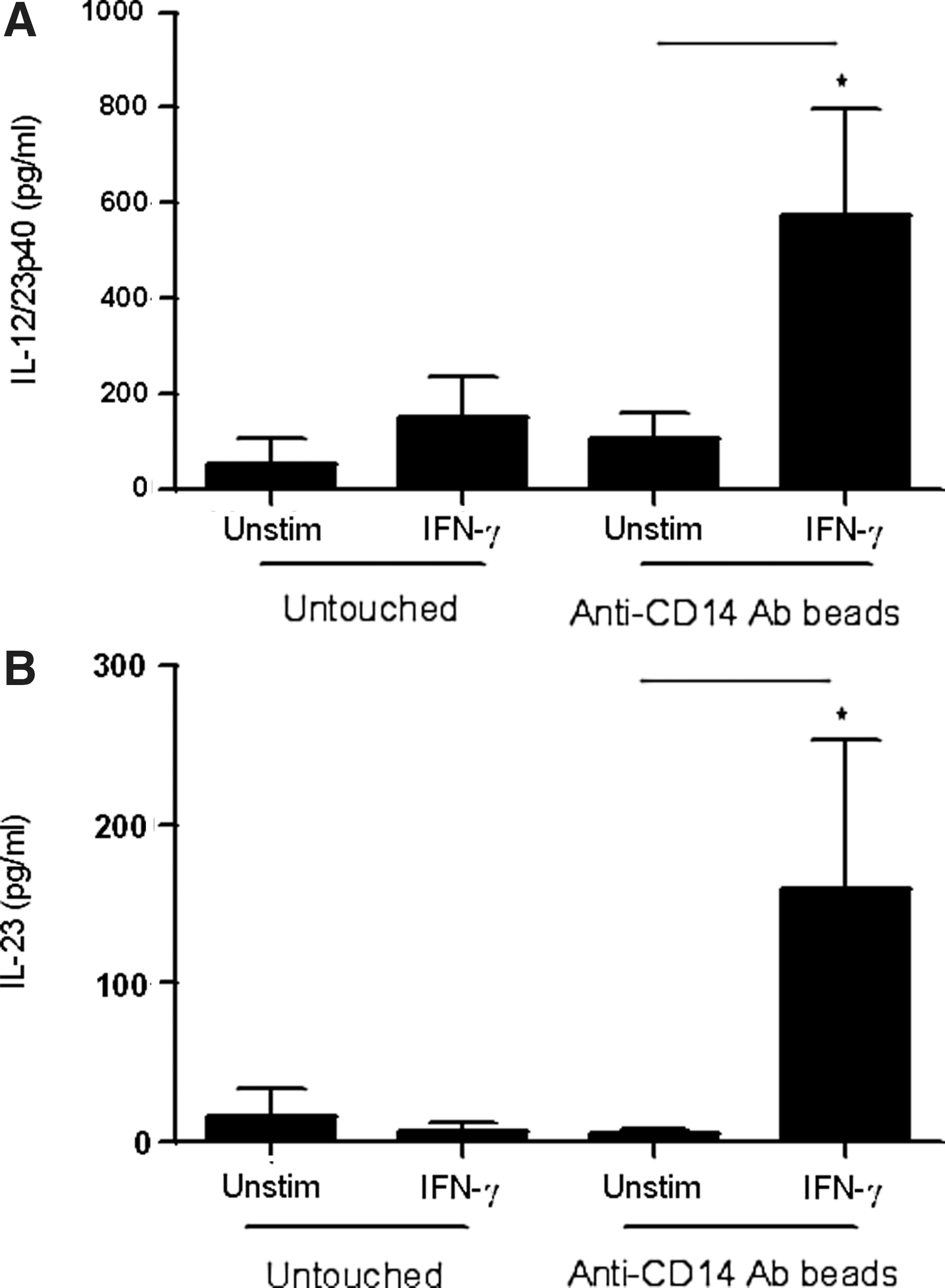
IFN-γ stimulation of CD14-activated monocytes triggers IL-12p40 and IL-23 production. Untouched negatively selected monocytes (1×106/mL) were stimulated with IFN-γ (10 ng/mL, 16 h, n=4). Negatively selected monocytes (1×106/mL) were treated with anti-CD14 antibody-conjugated beads at 4°C for 1 h followed by stimulation with IFN-γ (10 ng/mL) for 16 h (n=4). IL-12/23p40
Since monocytes were isolated by positive selection through the usage of anti-CD14 antibody-coated beads, we hypothesized that IFN-γ-induced production of IL-12p40 and IL-23 proteins was due to stimulation of monocytes through CD14 receptors. Therefore, negatively selected monocytes were cultured with anti-CD14 antibody-coated beads at 4°C for one hour followed by stimulation with IFN-γ. Interestingly, IFN-γ was able to induce significant production of IL-12p40 and IL-23 in these cells (Fig. 2A, B). These results suggest that IFN-γ induces the expression of IL-12p40 and IL-23 mRNA and proteins in cells activated through the CD14 receptors. To understand how CD14 engagement sensitizes positively selected monocytes to IFN-γ, we hypothesized that engagement of CD14 may induce the expression of IFN-γ receptors (IFNGR1/CD119) on positively selected monocytes. Therefore, we measured the expression of IFNGR1 on positively selected monocytes, negatively selected monocytes, and the negatively selected monocytes cultured in vitro with anti-CD14 beads. The results show that basal levels of IFNGR1 were significantly higher in positively selected monocytes compared to negatively selected cells (Fig. 3A). The expression of IFNGR1 on negatively selected cells cultured with anti-CD14 beads was also significantly higher compared to the negatively selected cells. A representative histogram showing IFNGR1 expression in positively selected, negatively selected, and negatively selected cells cultured with anti-CD14 beads is shown (Fig. 3A, right panel). Thus, enhanced expression of IFNGR1 may sensitize positively selected monocytes to produce IL-23 in response to IFN-γ stimulation.
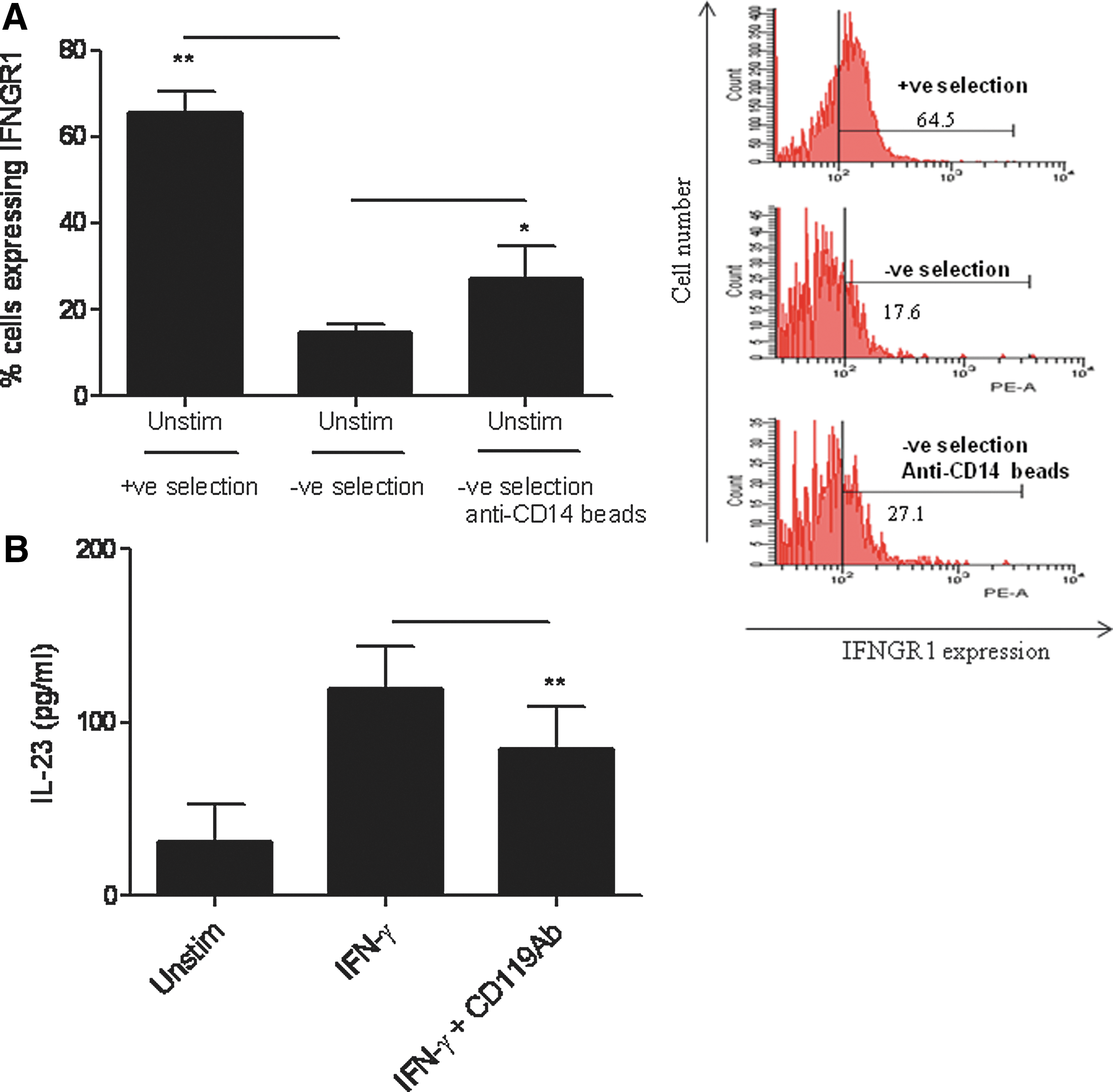
CD14 engagement sensitizes positively selected monocytes to IFN-γ stimulation by upregulating IFN-γ receptor 1 (IFNGR1).
To further demonstrate that the positively selected monocytes respond to IFN-γ stimulation and induce IL-23 production through the upregulation of IFNGR1, we blocked the IFNGR1 by anti-IFNGR1 antibodies (Fig. 3B). Treatment of positively selected monocytes with the neutralizing anti-IFNGR1 antibody significantly decreased IFN-γ-induced IL-23 production compared to the IFN-γ−stimulated cells treated with an isotype-matched control antibody.
Enhanced IFNGR1 expression on positively selected monocytes may also lead to enhanced IFN-γ−mediated effects other than IL-23 induction. Therefore, positively and negatively selected monocytes as well as negatively selected monocytes cultured with anti-CD14 beads were stimulated with IFN-γ for 16 h followed by analyzing the expression of TLR-4 receptors by flow cytometry. The results show that IFN-γ induced significantly higher expression of TLR-4 receptors on positively selected monocytes as well as negatively selected monocytes cultured with anti-CD14 beads compared to the negatively selected monocytes (Fig. 4A). IFN-γ priming of monocytes and antigen-presenting cells (APCs) has been shown to enhance their ability to produce IL-23 in response to a second stimulus such as LPS (Gautier and others 2005). Therefore, we determined whether enhanced TLR-4 expression following IFN-γ priming of positively selected monocytes would result in enhanced IL-23 production following LPS stimulation. Our results show that IFN-γ priming of positively selected monocytes significantly enhanced LPS (TLR-4)-induced IL-23 production compared to the LPS-stimulated cells alone (Fig. 4B). Subsequently, we focused our studies on the mechanism underlying regulation of IL-23 in positively selected monocytes in response to stimulation of IFN-γ-primed cells.
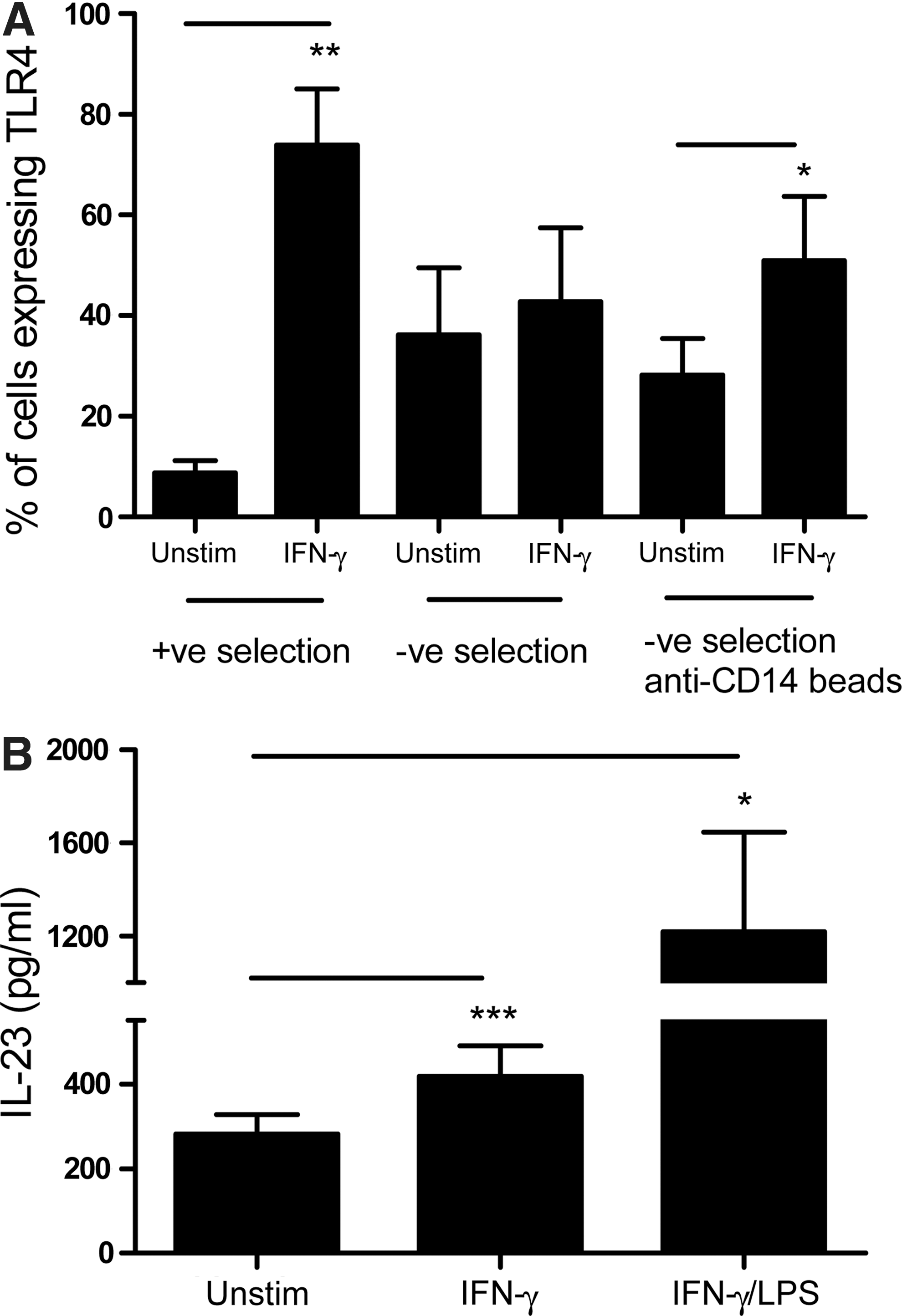
Upregulated IFNGR1 on positively selected monocytes exhibit enhanced TLR-4 receptors following IFN-γ stimulation leading subsequently to an enhanced LPS-induced IL-23 production.
The Jak/STAT pathway negatively regulates IL-12p40 protein and IL-23p19 mRNA expression, but not IL-23 expression following IFN-γ stimulation
IFN-γ mediates its effects primarily through the activation of the Jak/STAT pathway (Gil and others 2001). Therefore, the role of this pathway in regulating IFN-γ-induced IL-23 expression was investigated using the Jak inhibitor-I, which inhibits the Jak/STAT pathway. Monocytes pretreated with the Jak inhibitor-I were stimulated with IFN-γ alone and analyzed for STAT1 phosphorylation followed by measurement of IL-12p40 and IL-23 expression by RQ-PCR and ELISA. Cells pretreated with the Jak inhibitor-I showed inhibition of STAT1 phosphorylation in a dose-dependent manner (Fig. 5A). The unphosphorylated form of the protein was used to demonstrate equal loading control to attribute the increased levels of phosphorylation observed following IFN-γ stimulation. Interestingly, pretreatment with the Jak inhibitor-I significantly increased the expression of IL-23p19 mRNA and IL-12p40 protein production in a dose-dependent manner in cells stimulated with IFN-γ alone (Fig. 5B, C). The Jak inhibitor-I did not significantly enhance IL-23 protein expression in positively selected monocytes following stimulation with IFN-γ. However, a trend toward increased IL-23 production was observed in a dose-dependent manner (Fig. 5D). It may be pointed out that similar concentrations of the Jak inhibitor-I inhibited IL-4-induced CD44 expression in THP-1 cells [data not shown and (Gee and others 2001)].
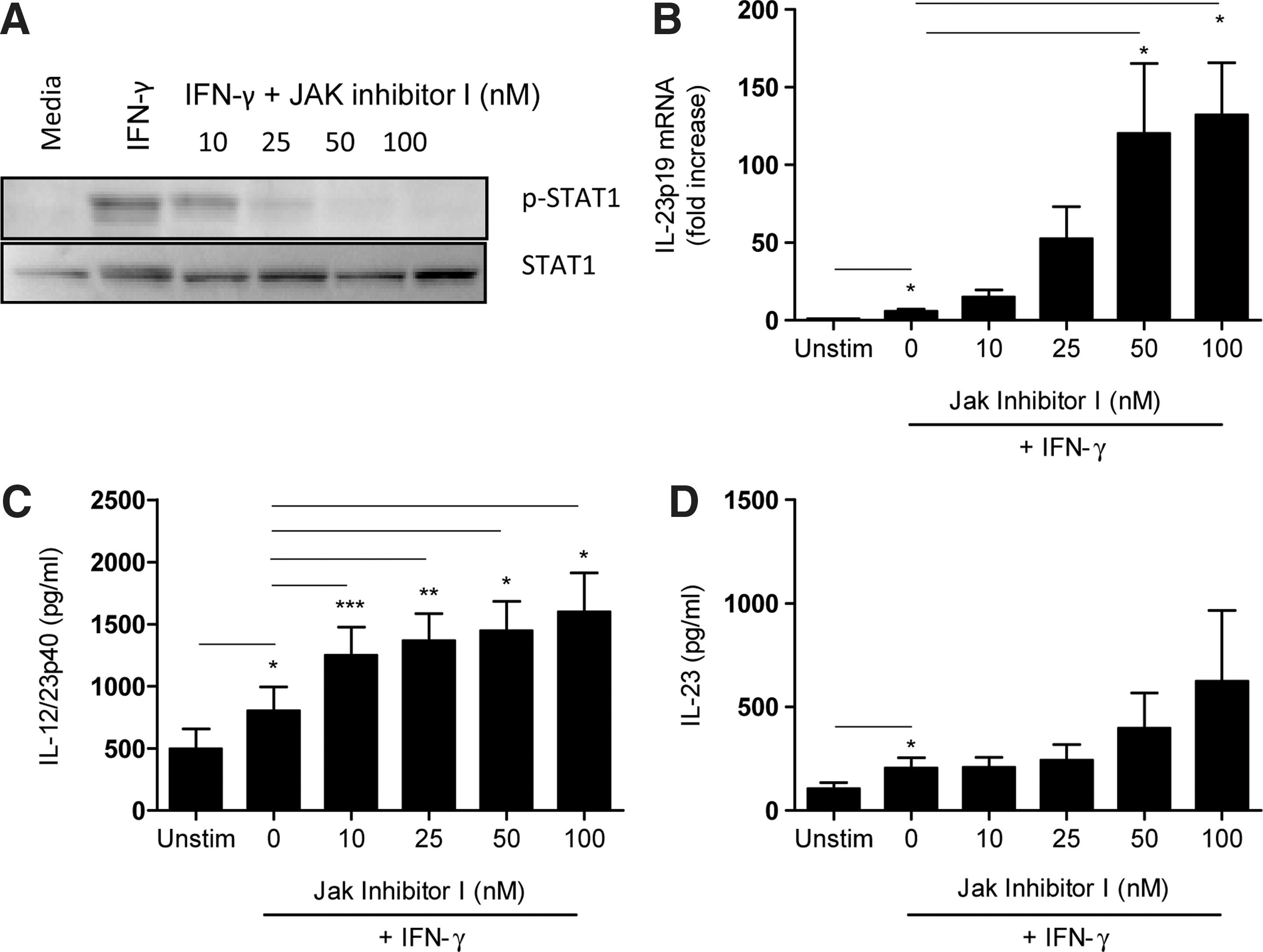
IFN-γ-induced IL-12p40 and IL-23 expression is negatively regulated by the JAK/STAT pathway.
IL-23 expression in IFN-γ-stimulated monocytes is negatively regulated by the PI3K pathway
IL-23 has also been shown to be positively regulated by the PI3K pathway following infection of human monocytic cells with Francisella tularensis and Mycobacterium tuberculosis (Yang and others 2006; Butchar and others 2007). Therefore, we determined the role of PI3K signaling in the regulation of IL-23 expression in IFN-γ-stimulated monocytic cells by using a pharmacological PI3K inhibitor, LY294002. We first demonstrated that pretreatment with LY294002 inhibited IFN-γ-induced Akt phosphorylation in monocytes (Fig. 6A). Inhibition of PI3K in IFN-γ-stimulated monocytes significantly enhanced IL-23p19 mRNA expression (Fig. 6B) as well as IL-12p40 and IL-23 protein production (Fig. 6C, D). These results suggest that IL-12/23p40 and IL-23 expression is negatively regulated by the PI3K pathway in IFN-γ-stimulated primary monocytes isolated by positive selection.
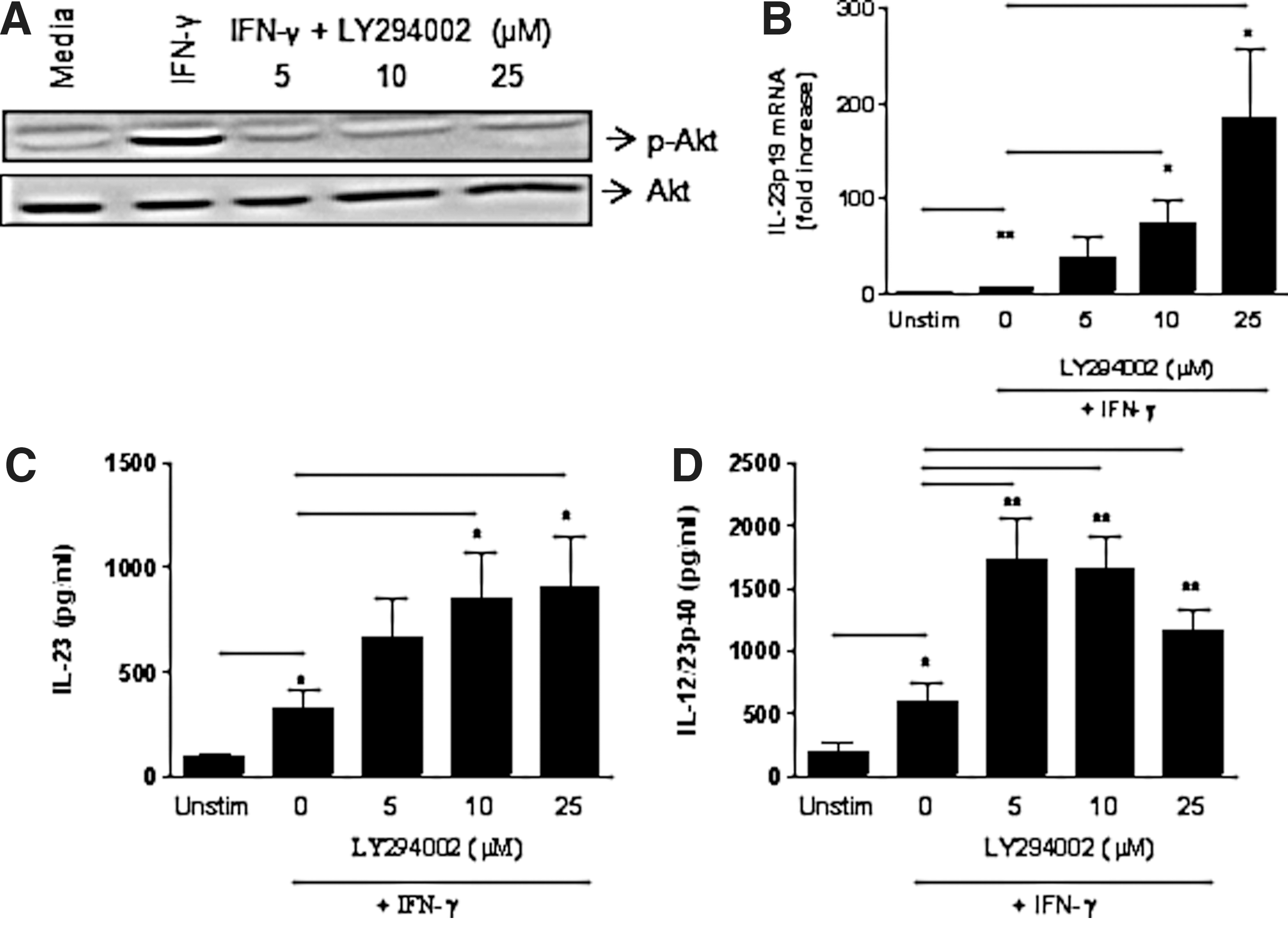
IFN-γ-induced IL-23 expression is negatively regulated by the PI3K pathway.
IL-23 expression in IFN-γ-stimulated monocytes is positively regulated by p38 and negatively regulated by the JNK MAPK pathways
We determined the role of MAPKs in the regulation of IL-23p19, IL-12/23p40, and IL-23 expression in IFN-γ-stimulated monocytes by employing inhibitors specific for p38 (SB203580), ERK (PD98059), and JNK (SP600125) MAPKs (Ma and others 2004; Dobreva and others 2008; Roses and others 2008). JNK, p38, and ERK MAPKs were phosphorylated in positively selected monocytes following stimulation with IFN-γ (Fig. 7). However, monocytes isolated by positive selection exhibited higher phosphorylation of JNK MAPK compared to the cells isolated by negative selection (data not shown). The specificity of different inhibitors was tested before starting the experiments. Cells pretreated with p38 (SB203580), ERK (PD98059), or JNK (SP600125) MAPK inhibitors showed a significant dose-dependent decrease in phosphorylation of p38, ERK, and JNK MAPKs, respectively, following IFN-γ stimulation (Fig. 7). These results show the ability of each inhibitor to block their specific signaling pathways.
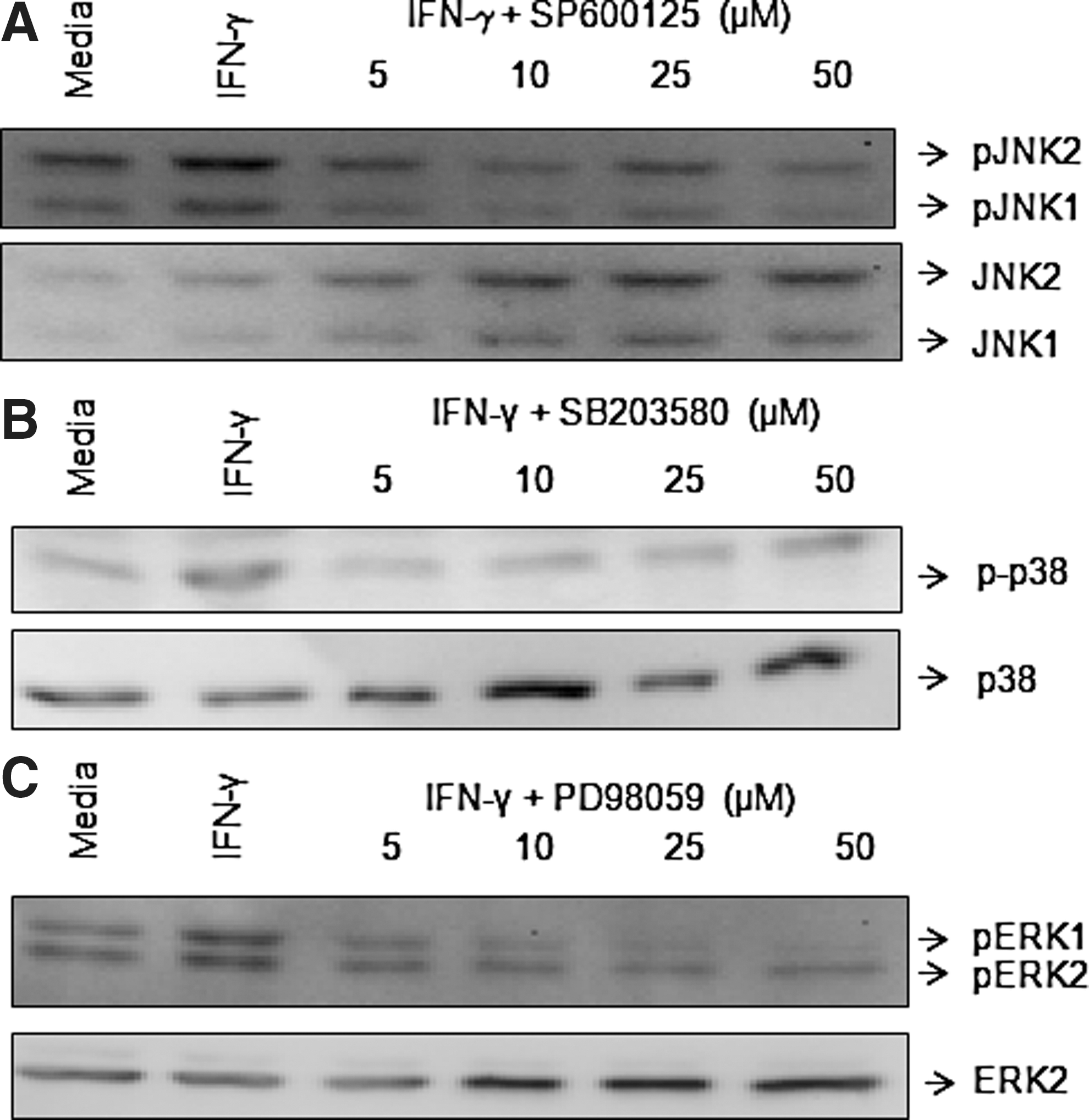
IFN-γ activates ERK, JNK, and p38 MAPKs. Positively selected monocytes (1×106/mL) were treated with increasing concentrations of inhibitor- specific ERK
The inhibition of JNK resulted in a significant increase in IFN-γ-induced expression of IL-23p19 mRNA, and IL-12/23p40 and IL-23 protein levels (Fig. 8A–C, left panels). These results suggest that the JNK pathway negatively regulates IFN-γ-induced IL-12/23p40 and IL-23 production. Interestingly, although inhibition of the p38 MAPK significantly enhanced IFN-γ-induced IL-23p19 mRNA expression (Fig. 8A, middle panel), SB203580 significantly decreased the levels of both IL-12/23p40 and IL-23 protein production (Fig. 8B, C, middle panels). In contrast, inhibition of ERK MAPK in monocytes stimulated with IFN-γ did not affect IL-23p19 mRNA or IL-12p40 and IL-23 protein levels (Fig. 8A–C, right panels). Since the JNK MAPK inhibitor, SP600125, enhanced IFN-γ-induced IL-23 production, we also show that SP600125 did not induce IL-23 production nonspecifically beyond the basal levels in unstimulated positively selected monocytes (Fig. 9).

IFN-γ-induced IL-12p40 and IL-23 expression is positively regulated by p38 MAPK and negatively regulated by the JNK MAPK pathway. Positively selected monocytes (1×106/mL) were treated with increasing concentrations of ERK (PD98059, right panel), JNK (SP600125, left panel), or p38 (SB203580, middle panel) MAPK inhibitors for 2 h followed by stimulation with IFN-γ (10 ng/mL, 16 h) for quantification of IL-23p19 mRNA expression by relative quantitative real-time PCR
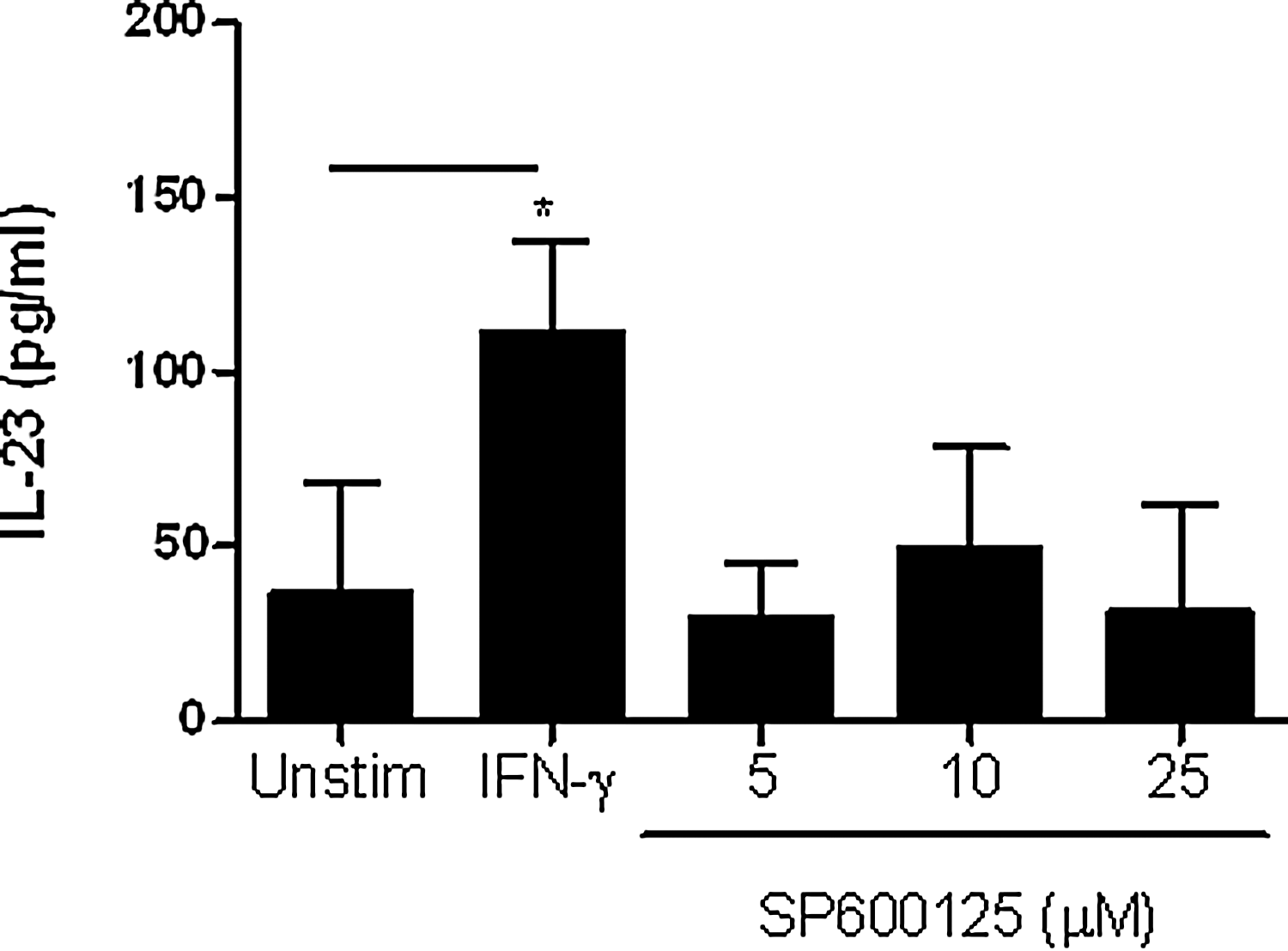
SP600125, a JNK MAPK inhibitor, does not induce IL-23 production. Positively selected monocytes (1×106/mL) were treated with increasing concentrations of SP600125. As a control, these cells were stimulated with IFN-γ (10 ng/mL) for 16 h followed by IL-23 quantification by ELISA. The results are shown as mean±SEM of 6 independent experiments, *p<0.05.
Discussion
Herein, we show that IFN-γ induces the expression of IL-12p40 and IL-23 in primary human monocytes following activation through the CD14 receptor. To understand the mechanism governing IFN-γ-mediated IL-12/23 production, we demonstrated that IFN-γ-induced IL-23 and IL-12/23p40 expression in monocytes is positively regulated by the p38 MAPK. Further, IL-12 and IL-23 expression is modulated by negative signals delivered by the Jak/STAT, JNK MAPK, and PI3K pathways in IFN-γ-stimulated primary monocytes isolated by positive selection.
IFN-γ priming of monocytes and APCs enhances their ability to produce IL-12 and IL-23 in response to a second stimulus such as LPS (Gautier and others 2005). IFN-γ priming thus provides a vital positive feedback mechanism for an enhanced immune response against invading pathogens (Butchar and others 2008; Kryczek and others 2008; Roses and others 2008). Additionally, TLR-2, TLR-4, and TLR7/8 ligands have been shown to induce IL-23 production independent of IL-12 in DCs (Roses and others 2008). Whether IFN-γ alone can enhance IL-12 or IL-23 expression by monocytic cells is not known. Herein, we show that IFN-γ failed to induce either IL-12p40 or IL-23 production in negatively selected monocytes (Fig. 1), monocyte-derived macrophages, and THP-1 cells (data not shown). However, activation of primary monocytes by anti-CD14 antibodies conferred responsiveness to IFN-γ resulting in significant production of IL-12p40 and IL-23 possibly through signals delivered via interactions with the CD14/TLR-4 complex. Previously, it was demonstrated that natural and recombinant soluble CD14 may stimulate monocytes in vitro (Landmann and others 1998). Further, positive selection of monocytes has been shown to influence cytokine production in generated DCs (Elkord and others 2005). We propose that outside of an LPS context, activation of monocytic cells through the CD14 receptor during positive selection significantly impacts cytokine expression and subsequent function of these cells in response to IFN-γ stimulation. These observations also suggest that monocytic cells similarly activated in vivo under certain pathological conditions may produce IL-12 and IL-23 in response to IFN-γ. Our results are supported by a study wherein generation of DCs from monocytes isolated by positive selection inhibited LPS-induced expression of costimulatory molecules and cytokines compared to the DCs generated from monocytes isolated by negative selection (Elkord and others 2005).
We have investigated the possible mechanism by which positively selected monocytes respond to IFN-γ leading to IL-23 production. Positive selection of monocytes through the use of anti-CD14 beads or stimulation of negatively isolated monocytes with anti-CD14 beads exhibited significantly higher levels of IFN-γ receptors. Enhanced IFNGR1 expression on positively selected monocytes may be due to signals delivered following an interaction of CD14 expressed on monocytes with the anti-CD14 antibody-conjugated beads (Landmann and others 1998). Enhanced IFNGR1 expression may also explain the ability of positively selected cells to respond to IFN-γ leading to a higher TLR-4 expression and subsequent enhanced production of IL-23 following LPS stimulation.
We and others have previously identified signaling pathways involved in LPS-mediated IL-12/23p40 expression (Murphy and others 1995; Plevy and others 1997; Becker and others 2001; Maruyama and others 2003; Murphy and others 2003; Dobreva and others 2008; Kaur and others 2008). In this study, we investigated the signaling pathways regulating IFN-γ-induced IL-23 and IL-12/23p40 expression in human monocytes isolated by positive selection. IFN-γ mediates its biological effects by activating the Jak/STAT (Butchar and others 2007) and by STAT1-independent pathways (Faries and others 2001; Yang and others 2006; Butchar and others 2007). Our results suggest that IFN-γ-induced IL-23p19 mRNA and IL-12/23p40 and IL-23 protein expression is negatively regulated by the Jak/STAT, PI3K, and JNK MAPK pathways. The mechanism by which IFN-γ-activated Jak/STAT1, PI3K, and JNK negatively regulate IL-23 or its subunits expression is not clear. However, it appears that IFN-γ-mediated signaling cascades, including Jak/STAT may cross talk with the signals delivered following the engagement of CD14 receptors that may result in the induction of IL-12p40 and IL-23 expression. IFN-γ induces the expression of the suppressor of cytokine-signaling (SOCS) proteins. SOCS1 is a negative regulator of Jak/STAT signaling and is believed to be an inhibitor of IFN-γ-induced IL-12 production in DCs (Hanada and others 2005). It is possible that the SOCS proteins induced by blocking the Jak/STAT pathway following IFN-γ stimulation may negatively regulate IL-23 production in monocytic cells. Although we have used low doses of kinase inhibitors and the increase in IL-23/IL-12 production is dose dependent, interference with ATP binding sites of other kinases cannot be ruled out.
There is some evidence that MAPKs regulate the expression of IL-12/23 subunits (IL-23p19) (Petro 2005; Dobreva and others 2008; Li and others 2008). Importantly, our results show that IFN-γ-induced IL-23 and IL-12/23p40 production is positively regulated by the p38 MAPK and negatively regulated by the PI3K and the JNK pathways. The mechanism by which IFN-γ activates PI3K or MAPKs is not clear (Gil and others 2001; Kaur and others 2008; Valledor and others 2008b). However, IFN receptors have been shown to interact directly with the p85 subunit of PI3K (Uddin and others 2000; Rani and others 2002) independently of JAK-mediated phosphorylation of the receptor (Uddin and others 2000). IFN-γ has been shown to recruit the myeloid differentiation primary response gene 88 (MyD88) leading to p38 MAPK activation (Sun and Ding 2006). Whether IFN-γ activates MAPK and PI3K pathways in positively selected human monocytes via MyD88 recruitment is not clear. Further, the mechanism by which PI3K and JNK MAPKs negatively regulate IL-23 expression needs to be investigated.
The transcription factors NF-κB, SMAD3, and ATF-2 have been implicated in the regulation of LPS-induced IL-23p19 gene expression (Al-Salleeh and Petro 2008). However, the involvement of specific transcription factors in IFN-γ-induced IL-23 expression remains unknown. IFN-γ activates several transcription factors in various cell types, including interferon regulatory factor (IRF)-1, IRF-8, PU.1, C/EBP, CIITA, IRF-9, and c-Jun in a STAT1-dependent manner (Eichbaum and others 1994; Harada and others 1994; Hu and others 2001). IFN-γ has also been shown to activate STAT3, STAT5, NF-κB, and AP-1 in a STAT1-independent manner (Deb and others 2001; Ramana and others 2002; Gough and others 2007). A comprehensive examination of the downstream transcription factors involved in IFN-γ-induced IL-23p19/p40 expression will provide further insight into the regulation of this cytokine. In the present study, it was not possible to delineate the involvement of IFN-γ-activated transcription factors required for IL-23 production as THP-1 cells did not produce IL-23 in response to IFN-γ. Moreover, it is extremely difficult to perform promoter analysis experiments in primary monocytes.
Our results suggest that IL-23p19 and IL-12/23p40 mRNA levels did not always correlate with IL-23 protein production specifically as seen in the p38 MAPK inhibition studies (Fig. 8). It has been reported that IL-23p19 mRNA levels are not associated with an increase in IL-23 production in response to LPS in other model systems (Utsugi and others 2006; Waibler and others 2007; Dobreva and others 2008; Brentano and others 2009). Inhibition of specific p38 and JNK MAPKs as well as PI3K pathways resulting in an increased expression of LPS-induced IL-23p19 has been reported (Petro 2005; Utsugi and others 2006; Dobreva and others 2008). However, inhibition of p38 MAPK was also shown to decrease LPS-induced IL-23 production (Petro 2005; Dobreva and others 2008). These observations suggest that post-transcriptional or post-translational mechanisms may regulate expression of heterodimeric IL-23 protein synthesis and/or secretion (Carra and others 2000). Several possible post-transcriptional and post-translational steps, such as IL-23 subunit mRNA stability, protein translation, intracellular protein degradation, and secretion of a newly synthesized protein, may be regulated by distinct signaling pathways. Interestingly, the p38 MAPK pathway has been shown to be an important post-transcriptional regulator of mRNA stability (Mahtani and others 2001; Rutault and others 2001). It is possible that the decrease in IL-23 protein production, despite an increase in IL-23p19 mRNA that we observed following inhibition of the p38 MAPK may be attributed to the post-transcriptional or post-translational activity.
In summary, we show for the first time that IL-23 and its subunit IL-12/23p40 are regulated by IFN-γ in primary human monocytes via p38 MAPK and independently of the Jak-STAT and PI3K pathways. We also show that the Jak/STAT, PI3K, and JNK MAPK pathways are negative regulators of IL-23 and IL-12/23p40 expression induced by IFN-γ. A schematic model depicting the signaling pathways involved in IFN-γ-induced expression of IL-23 in human monocytic cells is shown in Fig. 10. Elucidating the mechanism of IFN-γ-induced regulation of the IL-12 family cytokine expression, and particularly of IL-23, will further our understanding of immune responses in health and disease.

A model showing the signaling pathways involved in IL-23 induction following IFN-γ stimulation in human monocytic cells. Stimulation of CD14 with either anti-CD14 antibody-coated beads or LPS enhances the expression of IFNGR1 on monocytic cells. IFN-γ stimulation of monocytic cells initiates the oligomerization of its receptor subunit, IFNGR1 and IFNGR2, facilitating JAK activation and recruiting STAT to the receptor. Phosphorylated STAT is translocated to the nucleus. The IFN receptors have been shown to interact directly with the p85 subunit of PI3K, independently of JAK-mediated phosphorylation of the receptor (Uddin and others 2000; Rani and others 2002). IFNGR1 has also been shown to interact directly with MyD88 (Sun and Ding 2006), which may trigger activation of the MKK6/p38 MAPK pathway (Sun and Ding 2006), possibly through Pyk-2 activation (Halfter and others 2005). Further, IFN-γ has also been shown to activate JNK MAPK in macrophages (Valledor and others 2008a, 2008b), although the precise sequence of events leading to JNK MAPK activation is not known. Our results suggest that JAK/STAT, PI3K, and JNK MAPK negatively regulate IL-23 induction, whereas p38 MAPK positively regulates IL-23 induction in response to IFN-γ stimulation through the activation of a yet-identified transcription factor (TF). ↑ indicates enhanced expression, whereas ↓ indicates decreased expression following the blockage of a pathway.
Footnotes
Acknowledgments
This work was supported by grants from the Canadian Institutes of Health Research and the Ontario HIV Treatment Network. A.K. and J.B.A. are supported by a Career Scientist Award from the Ontario HIV Treatment Network. M.B. is funded by the Ontario Graduate Scholarship in Science and Technology. A.R. is funded by a Doctoral Scholarship Award from the Ontario HIV Treatment Network.
M.A.B. performed experiments and helped to write the article; A.A.R.R. performed experiments; J.B.A. designed experiments; A.K. designed experiments, interpreted results, and helped to write the article.
Author Disclosure Statement
The authors declare that no competing financial interests exist. This does not alter our adherence to all the policies on sharing data and materials.