
Abstract
Inflammatory bowel diseases (IBDs) are chronic inflammatory diseases of the gastrointestinal tract, which are clinically present as 1 of the 2 disorders, Crohn's disease (CD) or ulcerative colitis (UC) (Rogler 2004). The immune dysregulation in the intestine plays a critical role in the pathogenesis of IBD, involving a wide range of molecules, including cytokines. The aim of this work was to study the involvement of T-helper 17 (Th17) subset in the bowel disease pathogenesis by the nitric oxide (NO) pathway in Algerian patients with IBD. We investigated the correlation between the proinflammatory cytokines [(interleukin (IL)-17, IL-23, and IL-6] and NO production in 2 groups of patients. We analyzed the expression of messenger RNAs (mRNAs) encoding Th17 cytokines, cytokine receptors, and NO synthase 2 (NOS2) in plasma of the patients. In the same way, the expression of p-signal transducer and activator of transcription 3 (STAT3) and NOS2 was measured by immunofluorescence and immunohistochemistry. We also studied NO modulation by proinflammatory cytokines (IL-17A, IL-6, tumor necrosis factor α, or IL-1β) in the presence or absence of all-trans retinoic acid (At RA) in peripheral blood mononuclear cells (PBMCs), monocytes, and in colonic mucosa cultures. Analysis of cytokines, cytokine receptors, and NOS2 transcripts revealed that the levels of mRNA transcripts of the indicated genes are elevated in all IBD groups. Our study shows a significant positive correlation between the NO and IL-17A, IL-23, and IL-6 levels in plasma of the patients with IBD. Interestingly, the correlation is significantly higher in patients with active CD. Our study shows that both p-STAT3 and inducible NOS expression was upregulated in PBMCs and colonic mucosa, especially in patients with active CD. At RA downregulates NO production in the presence of proinflammatory cytokines for the 2 groups of patients. Collectively, our study indicates that the IL-23/IL-17A axis plays a pivotal role in IBD pathogenesis through the NO pathway.
Introduction
C
In particular, cytokines produced by T-lymphocytes appear to initiate and perpetuate chronic intestinal inflammation (Breese and others 1993; Pender and others 1998). Interestingly, cytokine production by lamina propria (LP) CD4+ T lymphocytes differs between CD and UC. Classically, the effector T-cells in CD produce a T-helper 1 (Th1) cell-type cytokine profile with elevated amounts of interferon (IFN)-γ and interleukin (IL)-12 (Neurath and others 2002; Neurath 2008; Rafa and others 2010), and these cells express augmented amounts of the Th1 cell-associated transcription factor T-bet (Neurath and others 2002). In contrast, the cytokine profile of LP T-lymphocytes in UC is characterized by the increased production of the Th2 cell-type cytokines IL-4 and IL-13, and the latter cytokine has been implicated in the generation of barrier dysfunction and ulcer formation in this disease (Heller and others 2005).
With the discovery of Th17 cells, many questions about the physiopathology of certain chronic inflammatory diseases can now be better understood, since the mechanism of such immune responses did not fit under the Th1 or Th2 paradigm (Cua and others 2003; Mesquita and others 2009).
Th17 cells are characterized by the master transcription factor RORγt, the surface markers IL-23R, and CCR6, and by high production of the proinflammatory cytokines IL-17A, IL-17F, IL-21, IL-22, and tumor necrosis factor α (TNF-α), and the chemokine CCL20 (Weaver and others 2007; Brand 2009). These proinflammatory cytokines are capable of promoting local tissue destruction (Mayer 2010).
It has been reported that the Th17 response plays key roles in the pathology of organ-specific autoimmunity and chronic inflammatory diseases (Tesmer and others 2008), such as rheumatoid arthritis, systemic sclerosis, systemic lupus erythematosus, Helicobacter pylori-associated gastritis, and renal allograft rejection (Kim and others 2007; Zhu and others 2008).
Th17 cells are activated by the combination of IL-6 and transforming growth factor (TGF-β), stimulating the transcription factor RORγt (Zhou and others 2007). IL-23 is produced by antigen presenting cells and promotes the maintenance of Th17 cells (Mancini and others 2008; Wang and others 2011). Both IL-6 and IL-23 activate signal transducer and activator of transcription 3 (STAT3), which is required for the development of the Th17-cell lineage, as it enables the expression of the transcription factor RORγt (Langrish and others 2004; Chen and others 2006b; Kan and others 2008; Durant and others 2010). STAT3 is a transcription factor that serves critical functions in development, cell growth, and homeostasis in a variety of tissues (Hirano and others 2000; Levy and Lee 2002). It plays critical roles in the pathogenesis of autoimmunity. Despite recent data linking STAT3 with IBD, exactly how it contributes to chronic intestinal inflammation is not known. STAT3 was critical in modulating the balance of Th17 and regulatory T (Treg) cells, as well as in promoting CD4+ T-cell proliferation (Durant and others 2010).
Many functions of retinoid have been identified in the regulation of immune responses (Kim 2008)
Generation and secretion of a high NO concentration by infiltrating cells may lead to perpetuation of local tissue damage (Kolios and others 2004). Some data report that proinflammatory cytokines induce NOS2 expression and activity in colonic epithelial cells (Huang and others 1998). In our previous studies, we have shown the IFN-γ involvement by the NO pathway in IBD pathogenesis (Rafa and others 2010). In the current study, we focused our attention on understanding the implication of the Th17 subset in IBD pathogenesis. In this way, we investigate the involvement of both the IL-17/NO pathways in this pathogenesis mechanism in 2 groups of Algerian patients reached with IBD (patients with CD and UC).
Patients and Methods
Patients
Forty patients with CD (14 men and 26 women; mean age 37.02±10.81 years; range 22–53 years) and 20 patients with UC (7 men and 13 women; mean age 33.16±8.31 years; range 24–47 years) were enrolled in this study. CD and UC were diagnosed by standard endoscopic and histological examination in the Department of Gastroenterology, Maillot Hospital, Algiers, Algeria. The disease activity of IBD was assessed using clinical parameters (endoscopic and histological criteria). The patients in active stage were tested before the beginning of therapy. Healthy controls (n=18) were obtained from adult volunteer donors. Each patient has given a written informed consent for the study required by the ethics committee of the National Agency of Research Development in Health (ANDRS), which supported our project.
Plasma collection
Blood samples collected from healthy donors and patients were centrifuged at 2,000 rpm for 10 min to obtain plasma. All plasma samples were stored at −45°C until messenger RNA (mRNA) extraction and cytokine (IL-17, IL-23, and IL-6) and NO determination.
Peripheral blood mononuclear cells
Preparation and culture
Peripheral blood mononuclear cells (PBMCs) were separated by density-gradient centrifugation on Ficoll-Hypaque (1.077 mg/mL; Sigma). The mononuclear fraction was collected and washed twice with sterile phosphate-buffered saline (PBS, pH 7.5) (2,800 rpm, 5 min). Cell viability was checked using 0.2% Trypan blue dye exclusion and was always >98%. Freshly isolated PBMCs were resuspended at a final concentration of 106 viable cells/mL in an RPMI 1640 culture medium supplemented with penicillin (100 U/mL), streptomycin (100 mg/mL), and 5% heat-inactivated fetal bovine serum (FBS; Sigma). The cultures were stimulated by human recombinant IL-17A at different concentrations (4, 8, 10, and 25 pg/mL), IL-6 (15 pg/mL), TNF-α (15 pg/mL), or IL-1β (25 pg/mL) in the presence or absence of At RA (10−7 M), N-monomethyl-
Monocyte isolation and culture
Human peripheral blood monocytes were isolated from PBMCs of patients with IBD and healthy donors as previously described. Adherent cell populations were obtained by incubation of PBMCs (109 cells/mL) in 24-well culture plates for 1 h at 37°C in the DMEM culture medium supplemented with 10% heat-inactivated FBS (Sigma). After trypsinization, the cell viability was checked using 0.2% Trypan blue dye exclusion and was always >97%. Freshly isolated monocytes were resuspended at a final concentration of 106 viable cells/mL in the DMEM culture medium supplemented with penicillin (100 U/mL), streptomycin (100 mg/mL), and 10% heat-inactivated FBS (Sigma). The cultures were stimulated by human recombinant IL-17A (25 pg/mL), IL-6 (15 pg/mL), TNF-α (15 pg/mL), or IL-1β (25 pg/mL) in the presence or absence of At RA (10−7 M). In all cases, unstimulated cultures were incubated as negative controls. The cultures were incubated for 24 h in a humidified atmosphere with 5% CO2 at 37°C. At the end of the culture period, the medium of each culture was harvested for NO (nitrite+nitrate) determination.
Colonic biopsies and culture
Multiple colonic biopsies were taken from patients who underwent colonoscopy in the department of gastroenterology, Maillot Hospital (Algiers, Algeria). The study included biopsies from inflamed mucosa of patients with UC (n=11) and with CD (n=16) in active stage. Biopsies from normal mucosa (n=6) were also used as negative controls. Colonic biopsies were immediately placed in the transport medium Hanks' balanced salt solution, pH 7.4, supplemented with antibiotics. The biopsies were stimulated by human recombinant IL-17A (25 pg/mL) in presence or absence of At RA (10−7 M). The cultures were incubated at 37°C in an atmosphere of 5% CO2 up to 24 h. Supernatants were then collected for NO (nitrite+nitrate) measurement. The total protein per well was estimated using the Bradford method, and the nitrite+nitrate contents of each well were expressed as μM/mg of total protein (Kolios and others 2000).
RNA extraction and real-time quantitative polymerase chain reaction
RNAs were isolated with a QIAGEN RNeasy kit (Qiagen). After reverse transcription into cDNA with a Reverse Transcription Kit (Bio-Rad), quantitative polymerase chain reaction (qPCR) was then performed on a MyiQ single-color real time-PCR detection system with SYBR Green Super mix (Bio-Rad), and gene-specific primers are summarized in Table 1. We normalized the gene expression amount to β-actin, HPRT, and GAPDH housekeeping gene and represented data as fold differences by the 2−ΔΔCt method, where ΔCt=Ct target gene−Ct MG, and ΔΔCt=ΔCt patients−ΔCt healthy donor. Fold changes were calculated using the comparative Ct method.
NO, nitric oxide; IL, interleukin; NOS2, NO synthase 2.
Histological analysis
Multiple colonic biopsies were taken from the patients with IBD (active CD, n=13; active UC, n=11) and were fixed for 24 h in a buffered formaldehyde solution (10% in PBS) at room temperature. The biopsies were dehydrated by graded ethanol and embedded in paraffin (solidification point 60°C–62°C). Tissue sections (thickness 3 μm) were deparaffinized with toluene and stained with hematoxylin and eosin. Digital images were captured with a digital camera at ×100 and ×400 resolution.
Immunohistochemistry
Sections (3 μm) were cut from paraffin-embedded tissues and mounted on positively charged Super frost slides. Tissues were deparaffinized and rehydrated through graded alcohols. All sections were incubated in 3% hydrogen peroxide (10 min) to blockade the endogenous peroxidase activity. Nonspecific binding was blocked by incubation (2 h) in PBS containing 5% skim milk. The sections were subsequently incubated overnight at 4°C in monoclonal mouse anti-p-STAT3 (Invitrogen-Life Technologies; diluted 1:100 in PBS containing 5% skim milk). Binding of the primary anti-p-STAT3 was detected with biotinylated goat anti-rabbit immunoglobulin horseradish peroxidase-conjugated streptavidin (1:500). Immunoreactive complexes were detected using the DAB system (Invitrogen-Life Technologies). The slides were counterstained briefly in hematoxylin (Sigma Aldrich), and mounted in Eukit (Sigma Aldrich). The slides were observed using a standard microscope (Zeiss), and pictures were taken using a digital camera at ×200 and ×400 resolution.
Western blot
The biopsies were lysed with a phosphate buffer containing 1% Triton X-100 and 1% protease inhibitor mixture (Sigma Aldrich). The protein concentrations were determined using a Bio-Rad Protein Assay (BioRad) according to manufacturer's instructions. SDS-PAGE was performed according to the Laemmeli procedure using gradient precast gels (4%–12% gradient, Bis–Tris; Invitrogen). Proteins were transferred onto a PVDF membrane (Millipore) and probed with a primary Ab [anti-p-STAT-3, 1/1,000; anti-inducible (iNOS), 1/1,000; Invitrogen-Life Technologies] and the rabbit anti-β-actin as an internal control. The membranes were incubated with peroxidase-conjugated secondary antibodies (anti-rabbit, 1:10,000; GE Healthcare) and washed again with a blocking buffer. Specific protein signals were visualized using the Western Lightning H Plus-ECL, Enhanced Chemiluminescence Substrate kit (PerkinElmer).
Immunofluorescence
PBMCs (106 cells/mL) of patients with IBD were fixed in paraformaldehyde (4%) for 30 min at room temperature. After blocking nonspecific binding with 5% skim milk and then permeabilized with 0.1% Triton X-100, a rabbit monoclonal antibody (anti-p-STAT-3, 1/100; anti-iNOS, 1/100; Invitrogen-Life Technologies) was used as a primary antibody for iNOS (NOS2) and pSTAT3 detection. Goat anti-rabbit IgG-FITC (fluorescein isothiocyanate) was used as a secondary antibody. Slides were covered with Kaiser's glycerin-PBS and observed using a standard microscope (Zeiss), and pictures were taken using a digital camera at ×100 resolution.
Enzyme-linked immunosorbent assay for human IL-17, IL-23, and IL-6
Plasma IL-17, IL-23, and IL-6 levels were determined by using an enzyme-linked immunosorbent assay (ELISA) kit according to the manufacturer's instructions (Invitrogen-Life Technologies). The absorbance was read on an ELISA reader at 450 nm (LABSYSTEM). These assays detected only human cytokines. The results are expressed as picograms per milliliter relating to a standardized dose curve of the relevant recombinant cytokine (IL-17A, IL-23, or IL-6).
NO assay
NO production was assessed by determination of the end products of NO oxidation. After reduction of nitrate (NO3−) by nitrate reductase-containing Pseudomonas oleovorans (ATCC 8062), the total nitrite [nitrite (NO2−) nitrate (NO3−)] was quantified by spectrophotometry (at 543 nm) in samples (plasma, supernatants of colonic mucosa, monocyte culture, and PBMC culture) after a reaction with the Griess reagent as described by Touil-Boukoffa and others (1998).
Statistical analysis
All results were expressed as mean±standard deviation. Minitab 15 was used for data analysis. The Student's t-test was used for comparison between different groups. Differences were considered to be statistically significant at P<0.05.
Results
Mediator Th17 cytokine/NOS2 mRNA expression in patients with IBD
To assess the mediator Th17 profile in Algerian patients with IBD, we analyzed the expression of mRNAs encoding Th17 cytokines (IL-17, IL-23, IL-6, and IL-1β) and some cytokine/chemokine receptors (IL-17R, IL-6R, and CCR6) in plasma from patients with UC and CD. The NOS2 mRNA expression was also analyzed by qPCR in these patients.
As shown in Fig. 1, analysis of cytokines and cytokines/chemokine receptors transcripts reveals that the mRNA transcript levels of the indicated genes are elevated in all IBD groups with active stage. Moreover, IL-17, IL-23, and IL-17R mRNA expression is higher in plasma of patients with active CD than in plasma of patients with active UC (∼3-fold, P<0.01). An ∼2- to 3-fold increase in IL-6 and IL-6R transcripts was found in patients with CD as compared with patients with UC (P<0.05 and P<0.01, respectively). A similar observation was noted in patients with active CD, who presented significantly higher IL-1β mRNA levels compared with patients with active UC (∼2-fold P<0.01). In contrast, no statistically significant difference in CCR6 mRNA expression was detected between the patients with active UC and active CD (P>0.05). The patients with active CD also had significantly greater NOS2 (iNOS) transcripts than the patients with active UC (∼2-fold P<0.05) (Fig. 1).
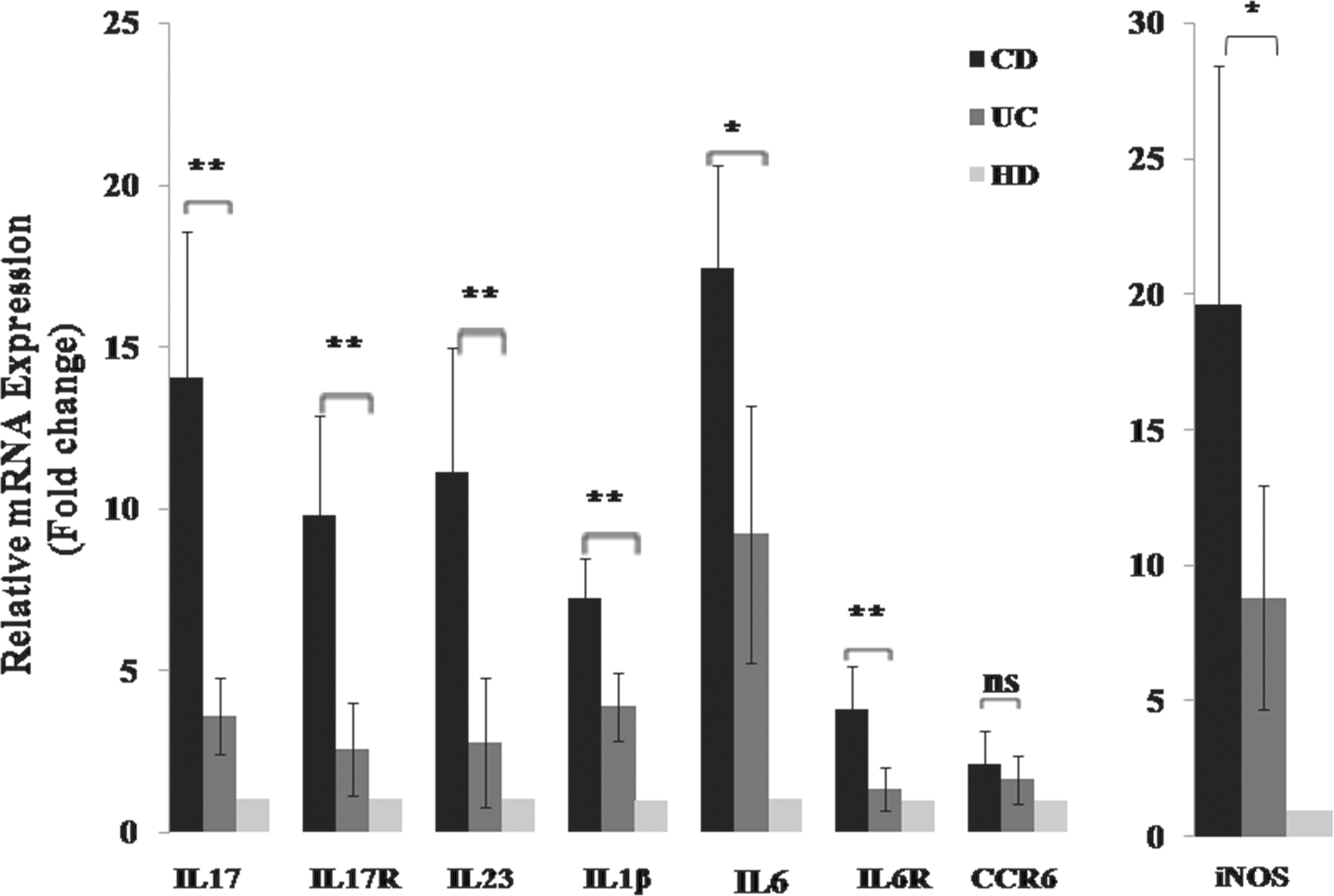
Messenger RNA (mRNA) transcripts of the indicated genes were quantitated in the sera of IBD patients in the active stage. Expression was normalized against β-actin, HPRT, and GAPDH housekeeping gene, and relative expression was represented data as fold differences by the 2−ΔΔCt method, where ΔCt=Ct target gene−Ct MG, and ΔΔCt=ΔCt patients−ΔCt healthy donor. Fold changes were calculated using the comparative Ct method. Data represent mean±standard deviation. Significant difference between 2 groups of patients (active UC/active CD) is indicated (**P<0.01, *P<0.05). ns, no significant difference, P>0.05. IBD, inflammatory bowel disease; CD, Crohn's disease; UC, ulcerative colitis.
The relationship between NO and proinflammatory cytokine levels (IL-17A, IL-23, and IL-6) in plasma of patients with IBD
As shown in Table 2, an analysis of NO and circulating cytokine production reveals that the levels of NO, IL-17, IL-23, and IL-6 are elevated in all IBD groups with active stage in comparison to the healthy controls. Moreover, the NO and cytokine (IL-17, IL-23, and IL-6) levels are higher in the sera of patients with active CD (65.20±9.35 μM, 77.48±3.83 pg/mL, 57.41±11.70 pg/mL, 183.76±19.32 pg/mL respectively) than in the sera of patients with active UC (49.91±5.64 μM, 57.21±6.42 pg/mL, 38.06±4.94 pg/mL, and 143.88±9.76 pg/mL, respectively) (P<0.05) (Table 2).
CD active phase, n=32; UC active phase, n=28; HD, n=15.
Values represent the mean±standard deviation.
Significance compared with HD: IL-17A, IL-6, and NO.
Significant difference between 2 groups of patients (active UC/active CD) is indicated (# P<0.01).
IL-23: no significant difference between 2 groups of patients (active UC/active CD) is indicated (P>0.05).
HD, healthy donors; CD, Crohn's disease; UC, ulcerative colitis; ns, no significant difference.
As shown in Fig. 2A and B the correlation between the NO levels and Th17 cytokine (IL-17A, IL-23, and IL-6) levels in plasma of patients with active CD and patients with active UC, respectively. A significant positive correlation was found in the NO levels and IL-17A levels in all cohorts of patients (Fig. 2A, B). We also showed the linear positive correlation between the NO levels and IL-23 levels in plasma of all patients (Fig. 2A, B). A similar observation was noted in patients with IBD, who presented a positive correlation between the NO levels and IL-6 levels in plasma (Fig. 2A, B). However, the correlation was higher with active CD than in that with active UC (CD: IL-17/NO. R2=0.80. P<0.01, versus UC: IL-17/NO. R2=0.60. P<0.05/CD: IL-6/NO. R2=0.79. P<0.01, versus UC: IL-6/NO. R2=0.69. P<0.05) (P<0.01), but no significant difference was found in correlation between the IL-23 and NO levels in plasma of patients with active CD and patients with active UC (IL-23/NO. R2=0.77. P<0.01, IL-23/NO. R2=0.72. P<0.01) (P>0.05) (Fig. 2A, B).
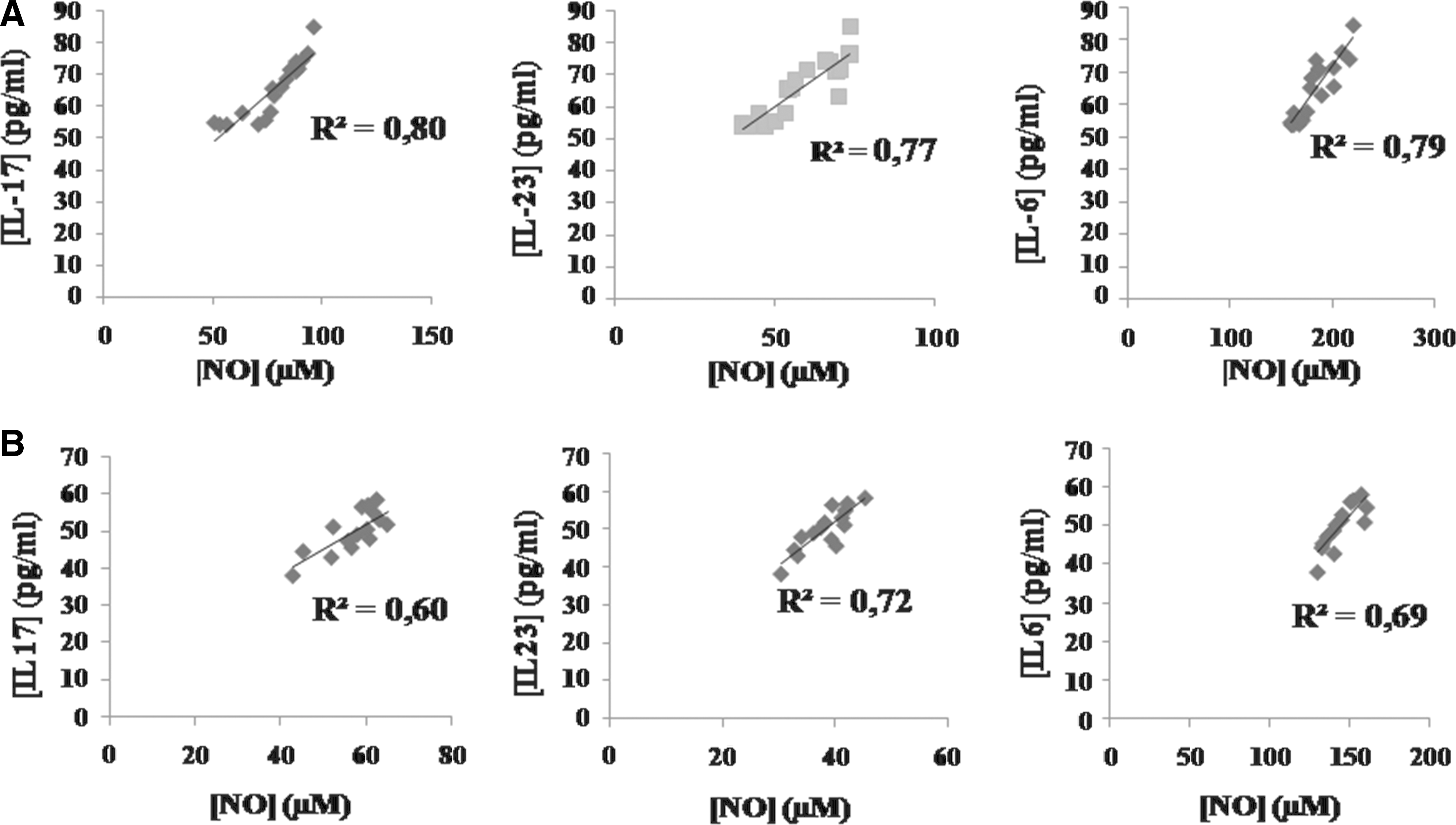
Correlation between the serum NO levels and the serum IL-17, IL-23, and IL-6 levels in Algerian patients with
Histological analysis of colonic mucosa
The histological study revealed a profound colonic inflammation characterized by crypt destruction. Inflammatory cell infiltration into mucosa in patients with CD (Fig. 3C, D) was also noticed compared to colonic mucosa control showing a normal structure (Fig. 3A, B). A similar observation was noticed in colonic mucosa of patients with UC (Fig. 3E, F).
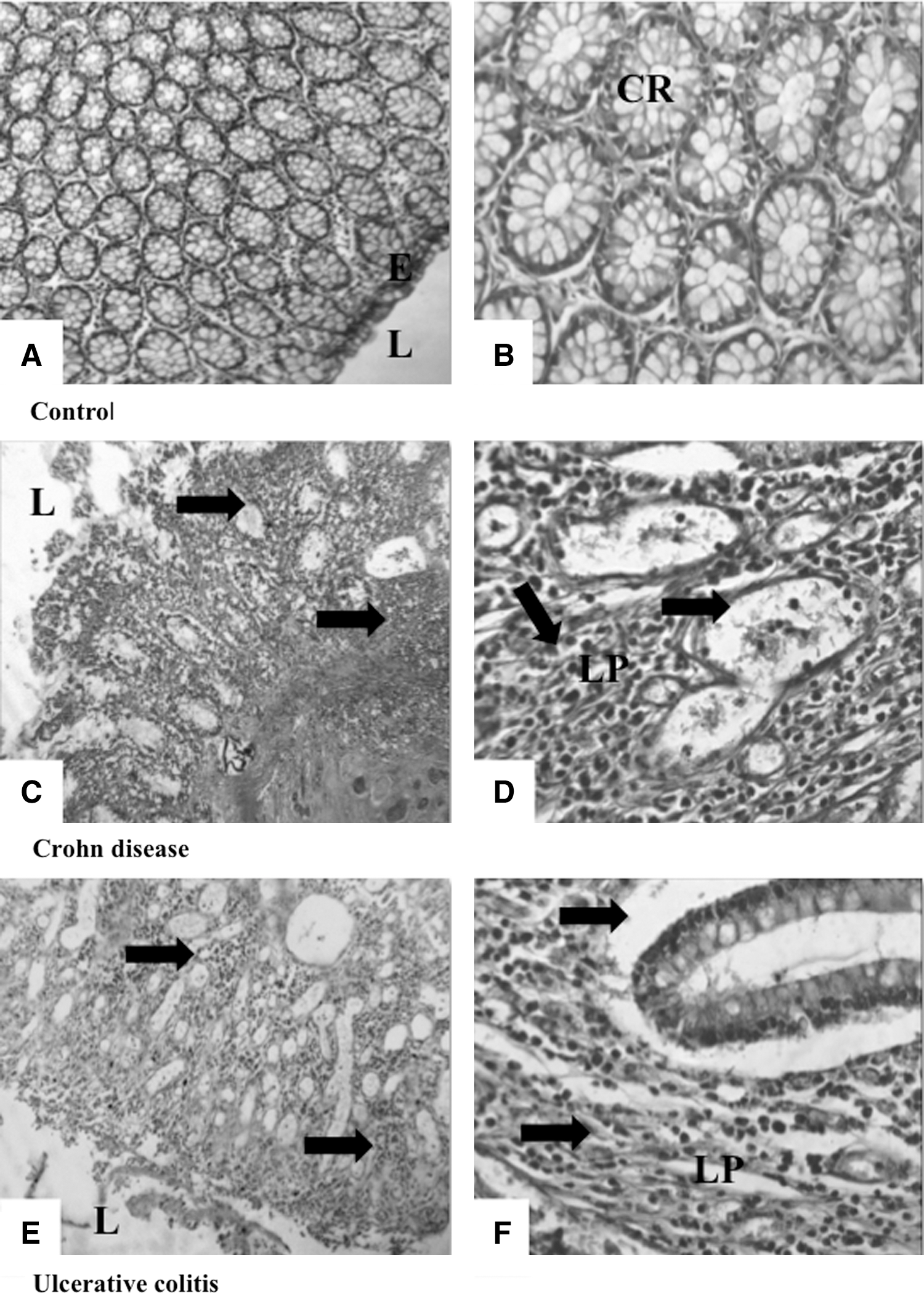
Representative photomicrographs (×100 and ×400) of hematoxylin-and-eosin-stained colonic mucosa from control
p-STAT3 and iNOS (NOS2) expression are the key regulators of Th17 cell implication in IBD pathogenesis
To investigate the importance of STAT3 in Th17-cell differentiation and its implication in Th17-cell-mediated IBD pathology, p-STAT3 protein expression in colonic mucosa was analyzed by Western blotting and immunohistochemistry. p-STAT3 expression was also analyzed in PBMCs by immunofluorescent staining. In addition, NOS2 (iNOS) protein expression was analyzed in colonic biopsies by Western blotting and in PBMCs by immunofluorescent staining.
In our study, we observed with interest that p-STAT3 expression was upregulated in colonic mucosa (Fig. 4, Fig. 5) and PBMCs (Fig. 8) of patients with active CD in comparison with controls. The similar observation was noted in colonic mucosa and PBMCs from patients with active UC.
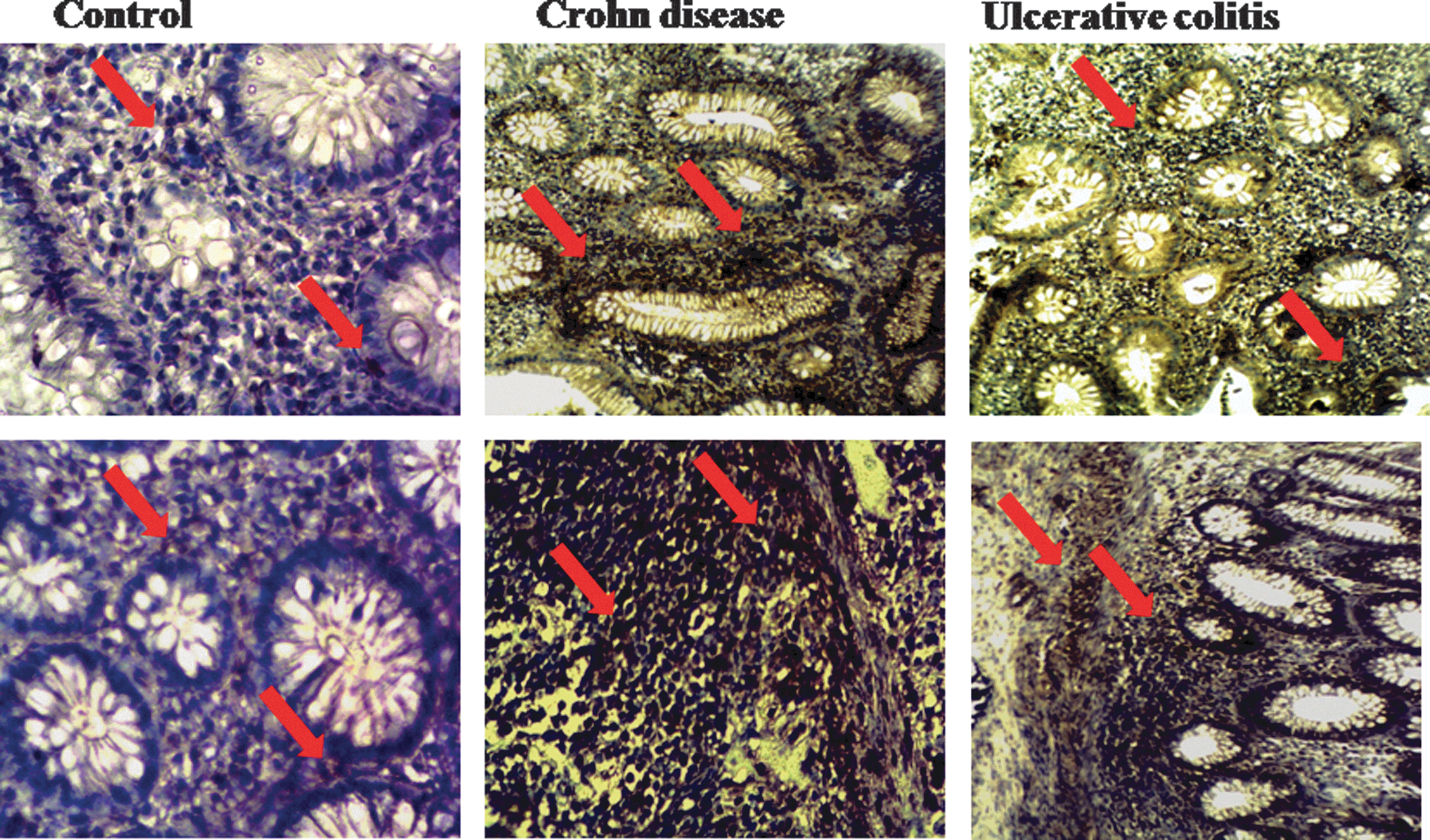
Immunohistochemical expression of p-STAT3. Control colonic mucosa shows a low expression of p-STAT3. This expression was profoundly increased in colonic mucosa of patients with Crohn's disease and Ulcerative colitis (×400). Arrows show P-STAT3 expression. STAT3, signal transducer and activator of transcription 3.

The expression of p-STAT3 and NOS2 in colonic mucosa of an patient with IBD detected by Western blot. Representative Western blot analysis of the total biopsy homogenate from colonic mucosa of UC and CD patients with active stage. An NOS2- and p-STAT3-specific polyclonal antibody recognized a protein whose molecular weight was ∼130 and 90 kDa, respectively. C, control colonic mucosa; NOS2, NO synthase 2.
Moreover, p-STAT3 expression is higher in colonic mucosa and PBMCs of patients with active CD than in colonic mucosa and PBMCs of patients with active UC (Figs. 4 and 8).
In addition, Western blotting analysis showed that NOS2 expression was also upregulated in colonic mucosa of all cohorts of patients with IBD in comparison with the controls (Fig. 5). We also noted that NOS2 expression was upregulated in PBMCs of patients with IBD by an immunofluorescence analysis (Fig. 6) compared with the controls (Fig. 6), especially in patients with active CD (Fig. 6).
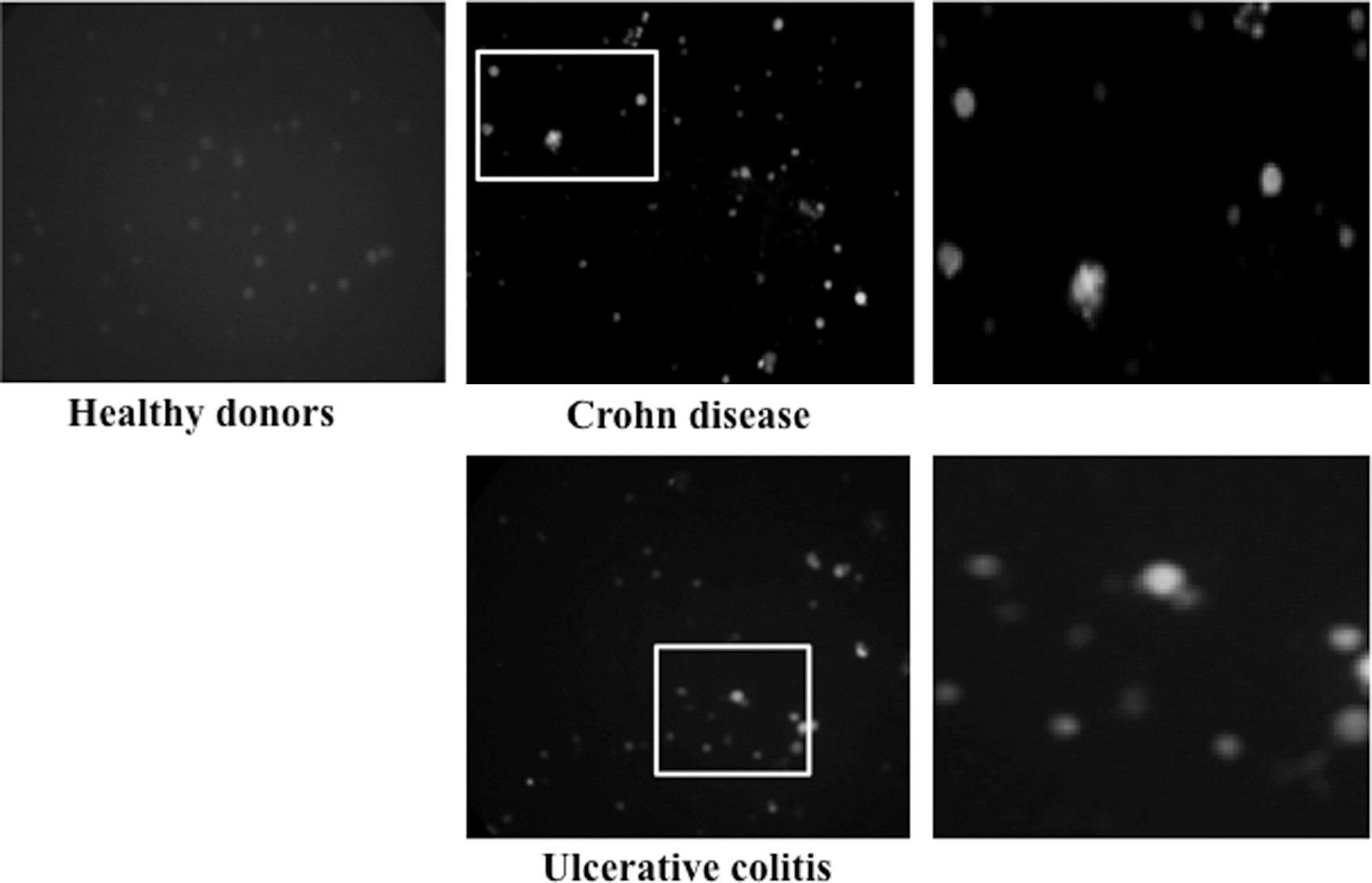
The expression of NOS2 in PBMCs of an patient with IBD detected by immunofluorescent staining. This expression was profoundly increased in PBMCs of patients with CD and UC. Healthy donors (×100). Insets shown enlarged at right. PBMCs, peripheral blood mononuclear cells.
At RA modulates cytokine-inducible NO production by PBMCs, monocytes, and colonic mucosa of patients with IBD
Considering the high levels of NO and IL-17A assessed in the plasma of patients with IBD, we decided to explore the effect of IL-17A at different concentrations on NO production in PBMC cultures. p-STAT3 protein expression was also analyzed in PBMCs stimulated with IL-17A (25 pg/mL). The effect of IL-17A was also explored in the absence or presence of At RA in PBMC cultures, monocyte cultures, and colonic mucosa cultures.
As shown in Fig. 7, stimulation with IL-17A at different concentrations increases NO production by PBMCs from patients with active CD compared with unstimulated cells (P<0.001). These findings were also observed in the culture with PBMCs of patients with UC (P<0.001) (Fig. 7). This increase occurred over a wide range of IL-17A concentrations (from 4 to 25 pg/mL) and occurred in a dose-dependent manner from 4 to 25 pg/mL (Fig. 7).
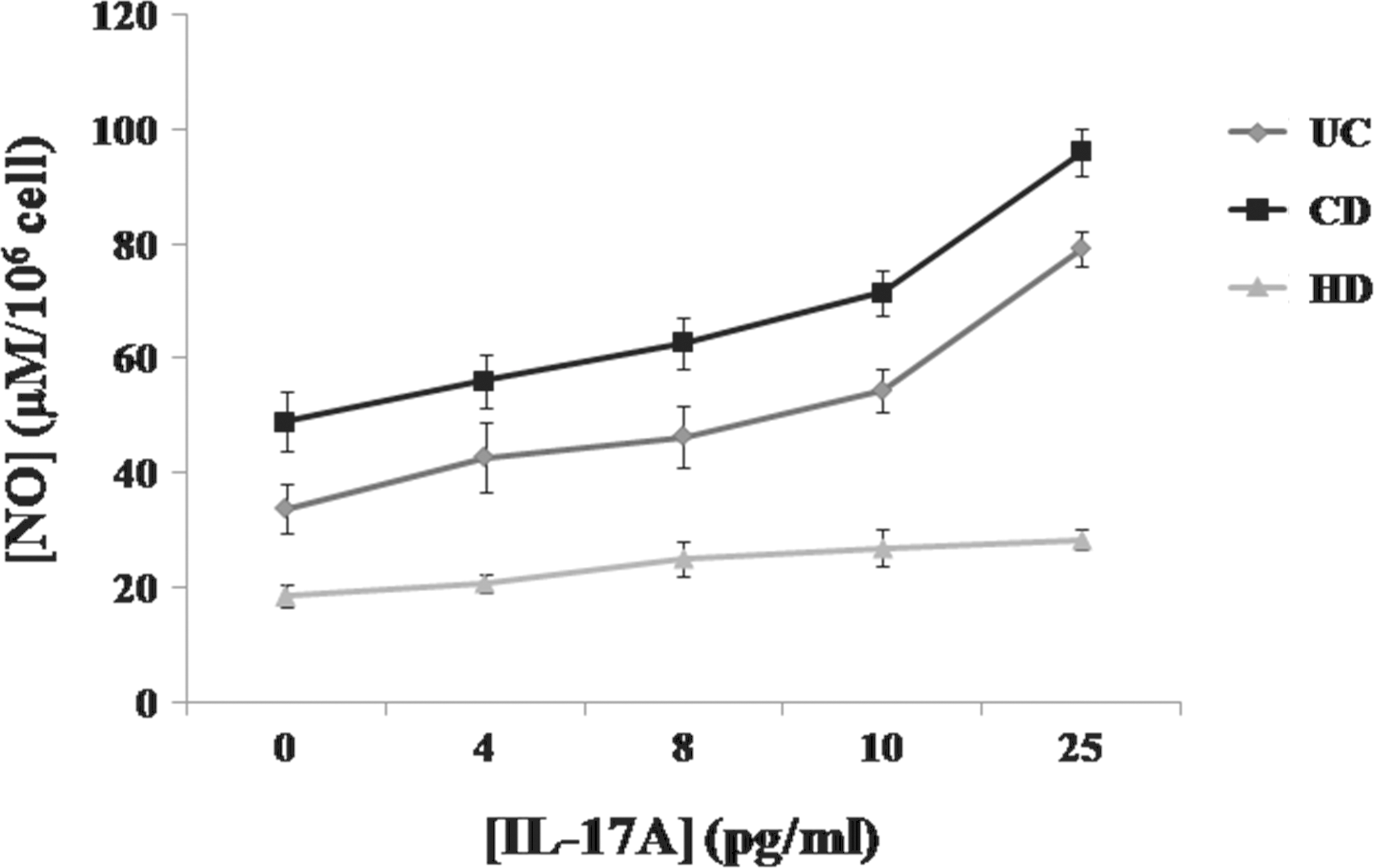
Effect of IL-17A at different concentrations (4, 8, 10, and 25 pg/mL) on NO production by PBMCs of patients with IBD in the active stage of the disease. Values represent mean±standard deviation. Significance compared with control: ***P<0.001. HD, healthy donor.
In addition, the immunofluorescence analysis showed that p-STAT3 expression was also upregulated in PBMCs stimulated with IL-17A (25 pg/mL) compared with unstimulated PBMCs (Fig. 8).
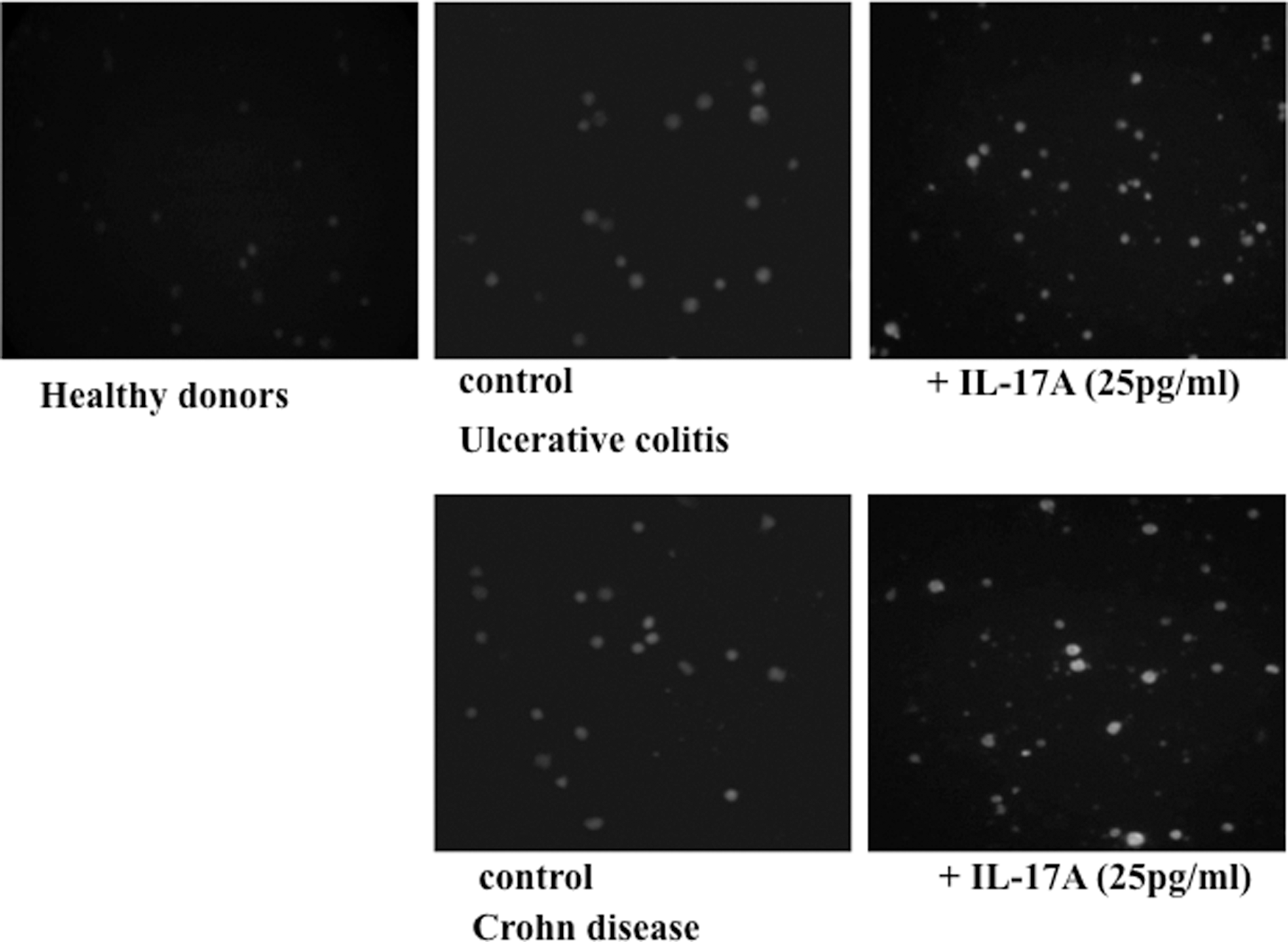
The expression of p-STAT3 in PBMCs stimulated with IL-17A (25 pg/mL) of patients with IBD detected by immunofluorescent staining. p-STAT3 expression was upregulated in PBMCs of active CD and active UC patients compared with healthy donors. This expression was also upregulated in PBMCs stimulated with IL-17A (25 pg/mL) compared with unstimulated PBMCs of patients with IBD (×100).
In our study, the mitogen stimulatory effect on NO production was also investigated in PBMC cultures. We report in our work that PHA stimulation increases NO production by PBMCs from patients with active CD and patients with UC compared with unstimulated cells (99.17±4.11 μM versus 49.05±5.2 μM and 84.67±6.17 μM versus 33.81±4.34 μM, respectively) (P<0.001). In contrast, if we consider the control conditions, we observe that the presence of L-NMMA caused a significant decrease in NO production in active CD patients' PBMCs (30.66±3.28 μM versus 49.05±5.2 μM, P<0.001) (Fig. 9). These findings were also reported in the culture using PBMCs from patients with UC (18.25±3.21 μM versus 33.81±4.34 μM, P<0.001). We also noticed that L-NMMA inhibited NO production in PBMC cultures stimulated by IL-17A in patients with CD and UC (45.73% and 32%, respectively, # P<0.001) (Fig. 9).
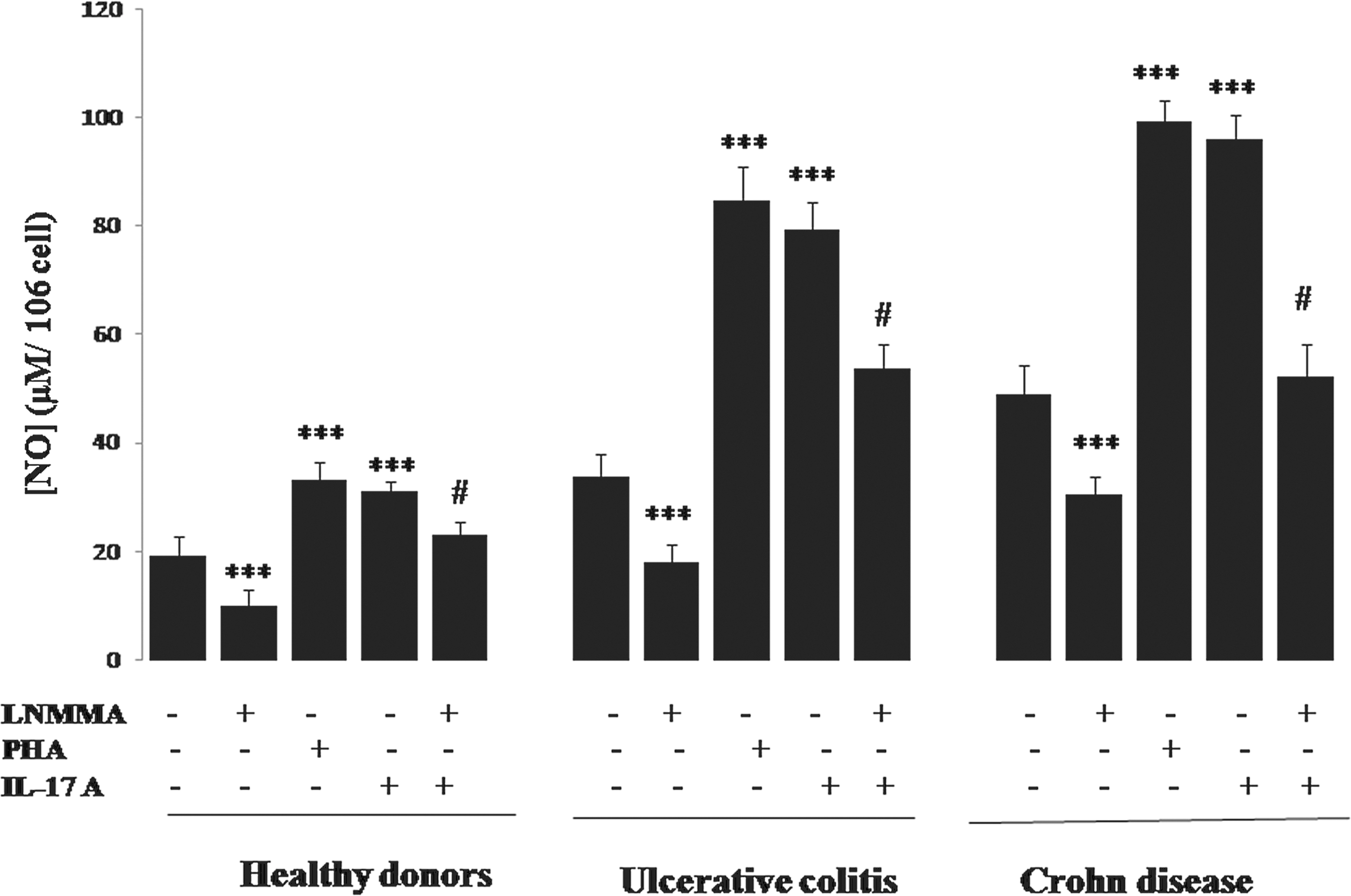
Effect of IL-17A, PHA, and L-NMMA on NO production by human mononuclear cells (PBMCs) of patients with IBD in the active stage of the disease. Values represent mean±standard deviation. Significance compared with control: ***P<0.001. L-NMMA inhibited NO production in PBMC cultures stimulated by IL-17A in CD and UC patients (45.73% and 32%, respectively, #
P<0.001). L-NMMA, N-monomethyl-
Activated PBMCs have been implicated in the course of IBD, as they are able to produce NO and many proinflammatory cytokines (IL-17A, IL-6, TNF-α, and IL-1β). We suggested that these molecules may contribute to disease progression. Here, we examined the ability of At RA to modulate NO production by PBMCs and monocytes in response to IL-17A, IL-6, TNF-α, or IL-1β. In the absence of proinflammatory cytokine stimulation, PBMCs (Fig. 10) and monocytes (Fig. 11) of all group of patients produced high levels of NO in comparison with healthy donors (P<0.001).
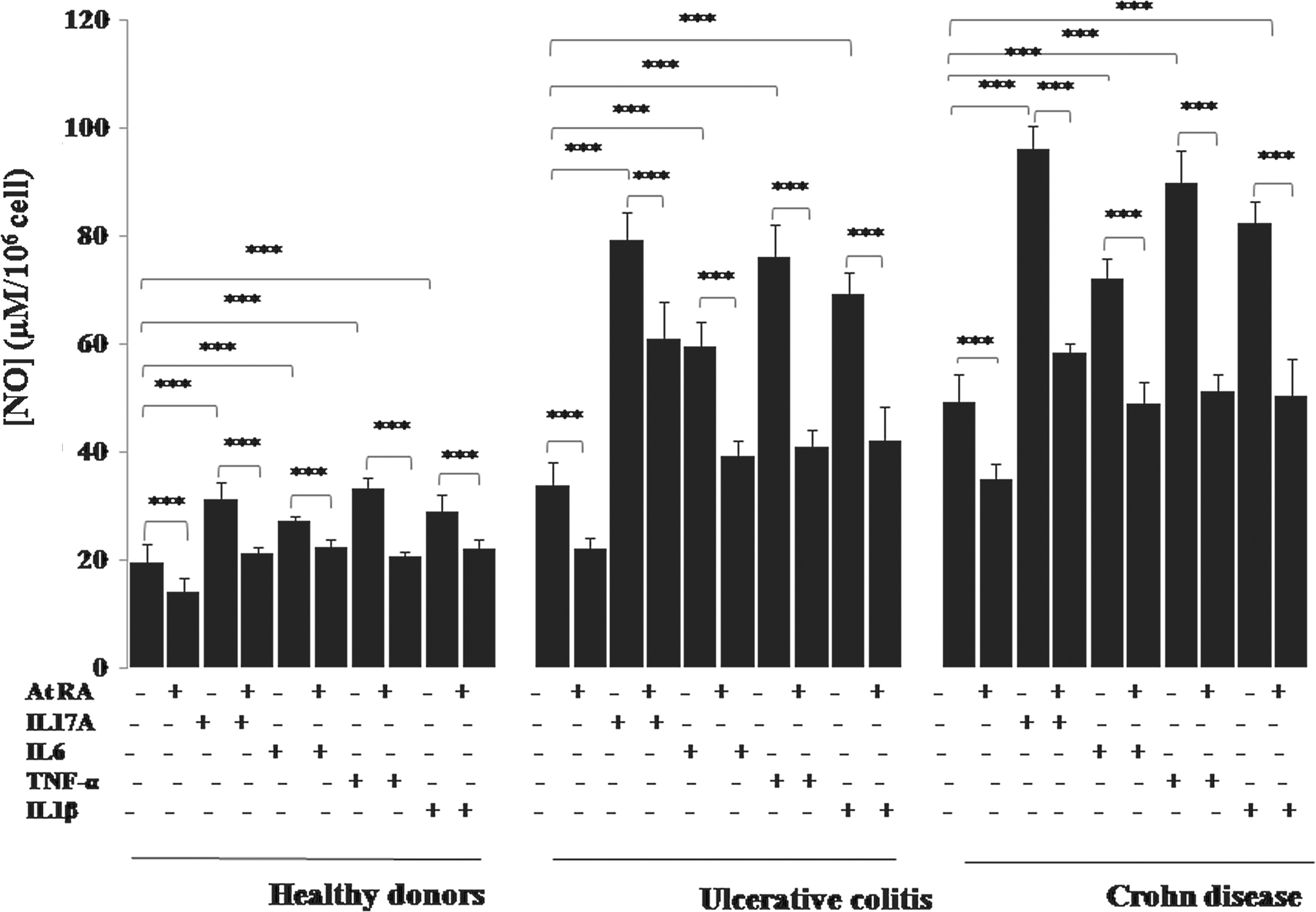
Effect of IL-17A, IL-6, TNF-α, and IL-1β in the presence or absence of At RA on NO production by human mononuclear cells (PBMCs) of patients with IBD in the active stage of the disease. Values represent mean±standard deviation. Significance compared with control: ***P<0.001. At RA, all-trans retinoic acid; TNF-α, tumor necrosis factor-α.
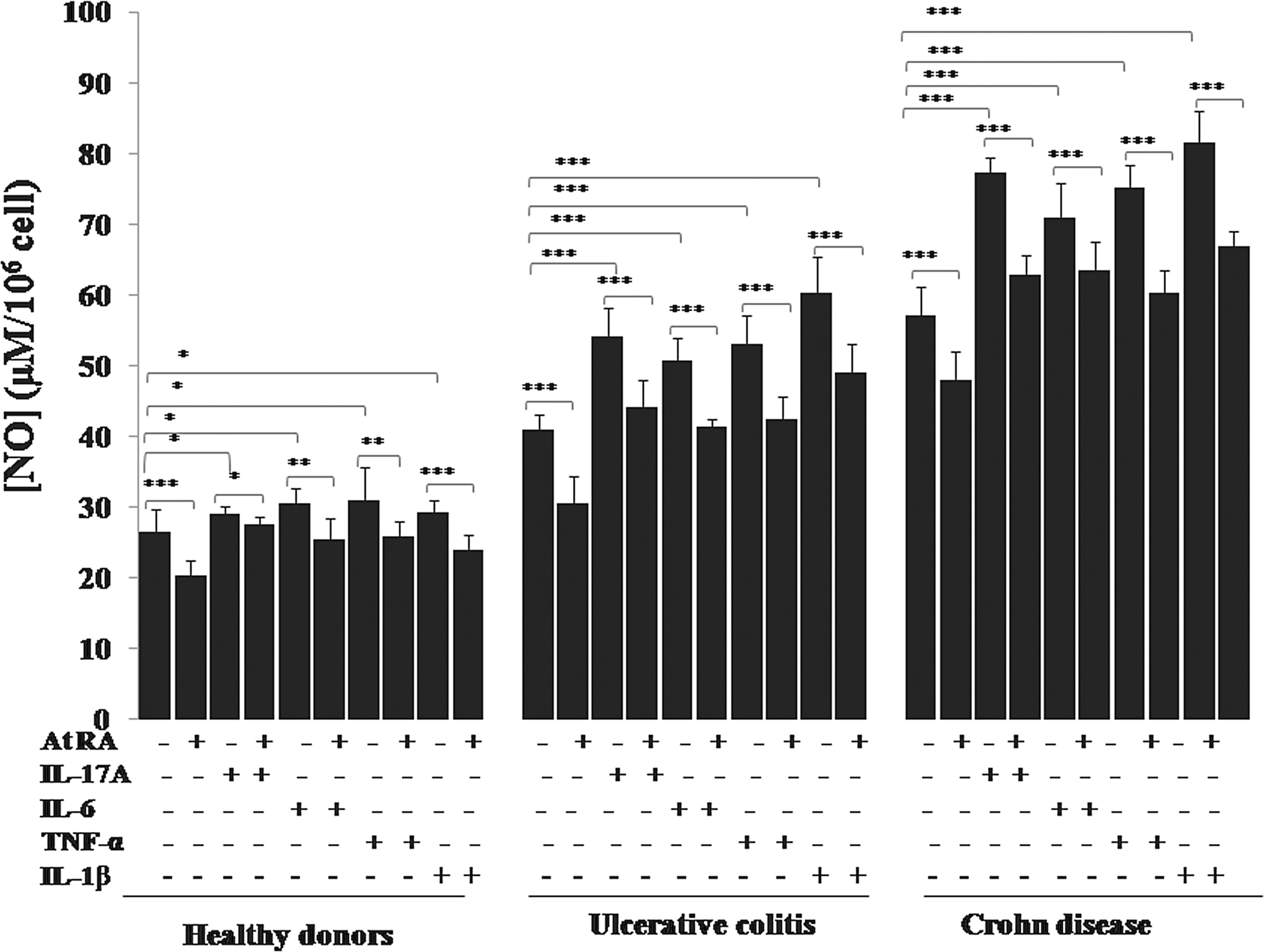
Effect of IL-17A, IL-6, TNF-α, and IL-1β in the presence or absence of At RA on NO production by monocytes of patients with IBD in the active stage of the disease. Values represent mean±standard deviation. Significance compared with control: *P<0.05, **P<0.01, ***P<0.001.
Interestingly, we report in our work a significant increase in NO production by PBMCs (Fig. 10) and monocytes (Fig. 11) in response to IL-17A, IL-6, TNF-α, or IL-1β stimulation in patients with UC and patients with CD in active stage (P<0.001). The similar effect of IL-17A was noticed in colonic mucosa cultures (P<0.001) (Fig. 12).
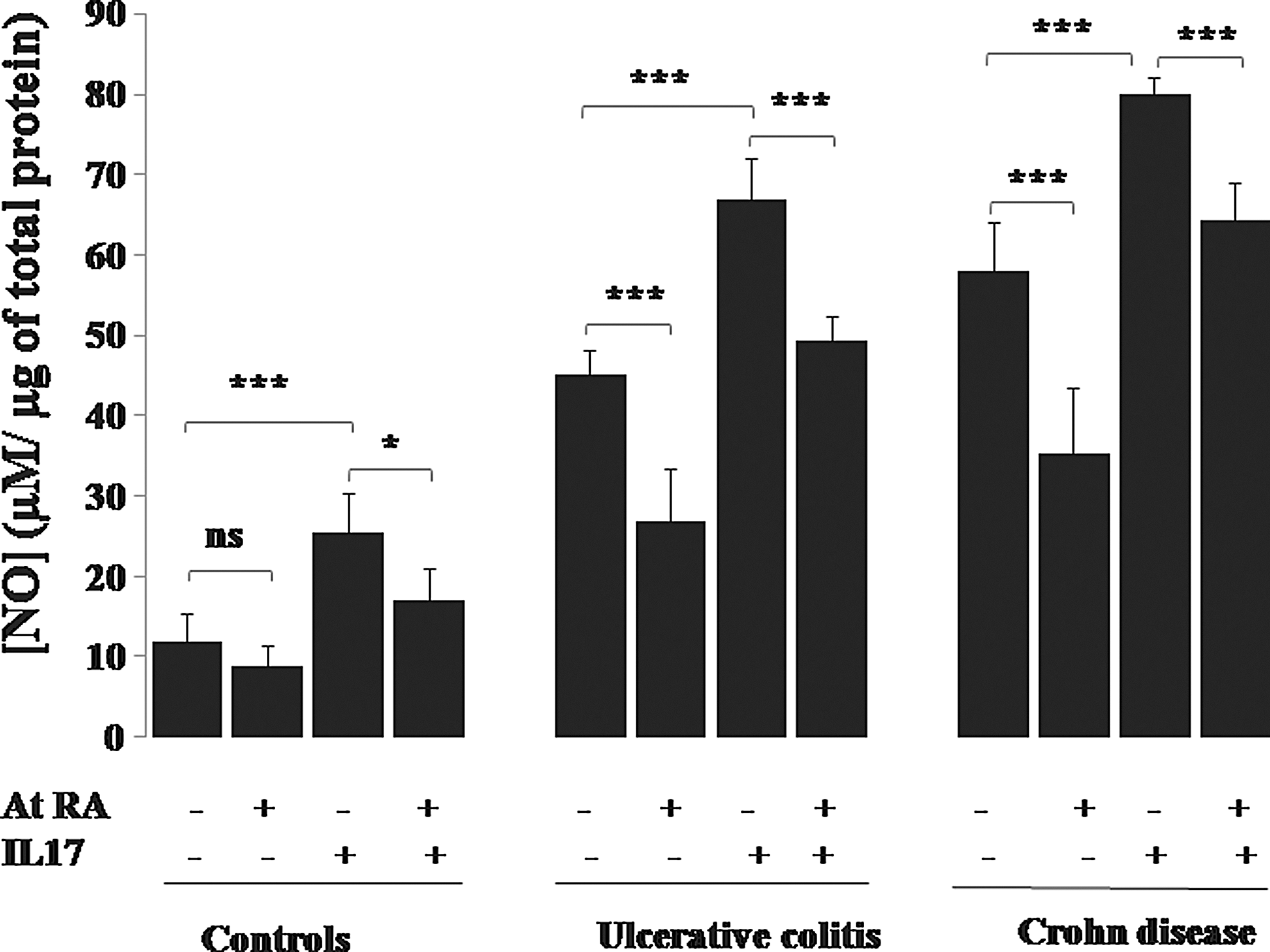
Effect of IL-17A in the presence or absence of At RA on NO production by colonic mucosa of patients with IBD in the active stage of the disease. Values represent mean±standard deviation. Significance compared with control: *P<0.05, ***P<0.001. ns, nonsignificant difference, P>0.05.
Our study showed that At RA inhibited IL-17A induction of NO production in PBMC cultures (Fig. 10), monocyte cultures (Fig. 11), and colonic mucosa cultures (Fig. 12) in all group of patients (P<0.001). Figure 10 illustrates the significant downregulation of NO production by UC and CD patients' PBMCs pretreated with At RA in response to TNF-α, IL-6, or IL-1β compared with the controls (P<0.001). The same profile was observed with monocytes pretreated with At RA from the patients with CD and patients with UC (P<0.001) (Fig. 11).
Discussion
Dysregulation, imbalanced production, or aberrant expression of cytokines is believed to be involved in the immunopathogenesis of inflammatory and autoimmune diseases (Leonard and O'Shea 1998; Lovato and others 2003). Many studies addressing the pathogenesis of IBD have focused on altered immune responses. A growing number of studies have reported an increase in sera and mucosal concentrations of proinflammatory cytokines and chemokines, including IL-1β, TNF-α, IL-6, IL-8, IL-16, IL-18 (Garside 2001; Middel and others 2001), IL-12, and IFN-γ (Rafa and others 2010)
In our study, we evaluated the expression of mRNAs for some Th17 cytokines (IL-17A, IL-23, IL-1β, IL-6), cytokines receptors (IL-17R, IL-6R, CCR6) and NOS2 in sera of patients with IBD and controls. We have observed with interest significant levels of mRNA transcripts of the indicated genes in all IBD groups with active stage. Our findings are in agreement with previous data showing the overexpression of Th17 signature cytokine genes in the inflamed intestinal mucosa in IBD (Geremia and others 2011). The study by Fujino and others (2003) showed that IL-17 expression is upregulated in the inflamed mucosa and in the serum of patients with IBD (Fujino and others 2003). An elevation in transcripts encoding IL-17A and IL-6 is detected in the biopsies from individuals with active CD (Holtta and others 2008). Previous studies indicated the key roles of TNF-α, IL-1β, and IL-6 in several diseases, including IBD (Steinman 2010).
Two independent studies demonstrated Th17 cells implication in human peripheral blood and in the gut from healthy individuals and patients with CD. These 2 studies showed that these T cells are characterized by the expression of CCR6 (Acosta-Rodriguez and others 2007; Annunziato and others 2007).
In our study, we observe with interest the positive correlation between Th17 cytokines (IL-17A, IL-23, and IL-6) and NO in plasma of patients with IBD. These results suggest that Th1 cytokines (Rafa and others 2010) and Th17 cytokines are both involved in upregulation of NOS2 induction in patients with IBD.
IL-17A is a member of the IL-17 family, which encompasses the potent inducers of inflammation, promoting cellular infiltration and the production of several proinflammatory cytokines and chemokines (Mesquita and others 2009), including IL-6, granulocyte/macrophage colony-stimulating factor, leukemia inhibitory factor, and intercellular adhesion molecule (Fossiez and others 1996; Awane and others 1999). In addition, IL-17 enhances the proinflammatory responses induced by IL-1β and TNF-α (Chabaud and others 2001; Katz and others 2001). IL-17 binds to and signals through IL-17 receptor A (IL-17RA), a member of the IL-17R family (Mesquita and others 2009)
Production of IL-6 during inflammation and its activation of STAT3 result in upregulation of RORγt and skewing of the developmental program toward the Th17 lineage (Takatori and other 2008; Korn and others 2009). Thus, the STAT3 pathway is situated at the juncture where Th17 and iTreg developmental programs diverge (Egwuagu 2009). The role of STAT3 in IBD has been most well documented by recent studies obtained from human IBD studies (Mudter and others 2005; Sugimoto 2008) and experimental IBD models (Siegmund and others 2002).
Our Western blotting, immunohistochemistry, and immunofluorescence analysis showed the upregulation of p-STAT3 in colonic mucosa and PBMCs of patients with IBD. This finding is in agreement with previous data showing that the patients with active UC had significantly more IL-6- and p-STAT3-positive epithelial cells than controls (Li and others 2010). Other studies showed that RORγt is induced in developing Th17 cells in the intestine through STAT3 activation (Sawa and others 2011)
The study by Durant and others (2010) showed that STAT3 bound to multiple genes involved in Th17-cell differentiation, cell activation, proliferation, and survival, regulating both expression and epigenetic modifications.
Our results show that histologic examination of biopsies obtained from patients with active disease reveal a severe mucosal degeneration, crypt loss, and destruction of epithelium in addition to cellular infiltrates. These observations correlate with the high NOS2 and p-STAT3 expression in colonic mucosa of patients with IBD with active stage. Mudter and other (2005) showed that increased pSTAT3 directly correlated with the histological degree of inflammation of tissues. Several studies demonstrated an overexpression of iNOS by inflamed epithelium and the proinflammatory cells in CD and UC patients (Singer and others 1996; Southey and others 1997). It can be argued that NO is clearly implicated in the mucosal injury observed during IBD pathogenesis. These results suggest that the histological degree of inflammation may be mainly induced by Th17 cytokines; these induce NOS2 upregulation in inflamed colonic mucosa.
The induction of iNOS by IL-17 was preceded by phosphorylation of p38 mitogen-activated protein kinase (MAPK), and inhibition of p38 MAPK activation completely abolished IL-17-stimulated NO release (Miljkovic and others 2005). IL-17 induced NO production in osteoblastic cells by a nuclear factor-κB (NF-κB)-dependent mechanism. This effect was associated with elevated mRNA levels of the NF-κB isoforms RelA and p50. (Van Bezooijen and others 2001).
In the last part of our study, we aimed to investigate the immunoregulatory effect of At RA on NO production in PBMCs, monocytes, and colonic mucosa cultures stimulated with some proinflammatory cytokines. Our current studies demonstrated that At RA inhibited NO production by PBMCs and monocytes stimulated with any one of the cytokines (IL-17A, IL-6, IL-1β, or TNF-α) and also inhibited NO production by colonic mucosa stimulated with IL-17A.
RA could modulate NOS2 gene expression by direct mechanisms mediated by RARα binding to the putative RARE in the promoter or the NOS2 gene (Sirsjo and others 2000). At RA could also regulate directly this expression by a protein–protein interaction with some transcription factors such as AP-1 (Schule and others 1991) and/or NF-κB (NA and others 1999), both reported to be involved in the modulation of NOS2 gene expression (Xie and others 1993)
Taken together, our study indicates in Algerian patients that the IL-23/IL-17 axis is actively implicated in IBD disease, and it correlates with the NO pathway, particularly in CD disease. At RA presents as a promising usefulness tool in IBD.
Footnotes
Acknowledgments
We thank the technical and surgical staff of the Department of Gastroenterology, Maillot Hospital, Algiers, Algeria, for providing blood and colonic biopsies. We thank all voluntary participants in this study. This work was supported by a grant from the ANDRS (National Agency for Scientific Development and Research in Health) Project No.03/06/01/10/011).
Author Disclosure Statement
No competing financial interests exist.