
Abstract
Interleukin (IL-35) is a newly identified heterodimeric cytokine belonging to the IL-12 family. It contains Epstein-Barr virus-induced gene 3 subunit and IL-27 p35 subunit. Although its receptor and signaling pathway are not clear, we presumed that its receptor is composed by two chains that might be similar to those receptors of IL-12, IL-23, and IL-27. We also believe that the signal transducer activator of transcription family members is involved in its signaling pathway. It was reported that IL-35 could suppress Teff cell proliferation and Th17 development. It was considered to have a potential therapeutic effect against immune diseases. In our perspective, the finding of IL-35 is of great significance, since it can regulate T cells, which is an important therapeutic target of immunological disorders. IL-35 would promote the development of different kinds of vaccines, even vaccine for special cancer, and be promising to cure autoimmune and inflammatory diseases.
Introduction
Structure
IL-35 is a newly identified heterodimeric cytokine, containing Epstein-Barr virus-induced gene 3 (EBI3) and IL-27 p35 subunits (Stern and others 1990; Schoenhaut and others 1992; Devergne and others 1997). It belongs to the IL-12 cytokine family that includes other three cytokines IL-12, IL-23, and IL-27 (Xiao-Qing and others 2010; Zhao 2010). IL-12 plays an important role in naïve CD4+ T-cell differentiation and the production of interferon-γ (IFN-γ) (Devergne and others 1997; Yoon and others 2000; Zhang and Wang 2008). IL-12p70 consists of p40 and p35 subunits, and its receptor includes IL-12Rβ1 and IL-12Rβ2 chains. IL-23 is synthesized by p19 and p40 subunits, and IL-12Rβ1 and IL-23R consist of its receptor. IL-27 contains p28 and EBI3 subunits, and its receptor is composed by WSX1 and gp130 (Zhao 2010). Similarly, IL-35 comprises EBI3 and p35 subunits. Although the receptor of IL-35 is not clear yet, we supposed that it also consists of two chains named IL-35Rα and IL-35Rβ. One chain mainly serves as a ligand, and the other mainly acts as a signal transmitter. One subunit of IL-35, EBI3, is widely expressed in EBV-transformed B-lymphocytes and tissues, such as tonsil and spleen (Devergne and others 1996, 1997). It is also expressed in human atherosclerosis, which is a complex immune response involving both inflammation and autoimmune processes (Kempe and others 2009). EBI3 may exert a critical role in the immune system. Yang and others (2008) suggested that EBI3 could negatively regulate IL-17, IL-22, and Th17 transcription factor RORγt and exert protective immunity against inflammation. Tong and others (2010) performed a delayed-type hypersensitivity (DTH) animal model using EBI-3-deficient (EBI-3−/−) mice. Their results showed that EBI-3−/− mice got a severe inflammatory response of DTH than that in control wild-type mice. These data suggested that EBI-3- or EBI-3-related cytokines may be potential targets for immune-associated illnesses. The subunit of IL-12p35 could lead to the progression of Herpes stromal keratitis (HSK) in mice, which is IL-12p40 independent (Frank and others 2010). The two subunits of IL-35 do have their own ability to regulate immunity and the process of inflammation. When they combine together to form the heterodimer, the p35 subunit may act as a ligand, and the other subunit EBI3 may mainly exert its immunological function.
Production
It has been reported that the source of IL-35 is different from that of IL-12, IL-23, and IL-27. The later three cytokines are all primarily secreted by antigen-presenting cells (APCs), including macrophages, monocytes, and dendritic cells (DCs). They are activated by the combination of pathogen-specific patterns (Ma and others 1996; Pflanz and others 2002; Hibbert and others 2003). On the contrary, only forkhead box protein 3 (Foxp3)+ regulatory T-cells could produce bioactive IL-35 (Collison and others 2007). Meanwhile, its subunits EBI3 and p35 are also found to be coexpressed in peripheral γδT-cells, CD8+ T-cells, and placental trophoblasts (Devergne and others 1997). Whether these cells could produce bioactive IL-35 still remains to be determined. According to the fact that APCs could secrete both IL-12 and IL-27, containing one subunit of IL-35, respectively, APCs may also produce IL-35 in certain situations. Seyerl and others (2010) demonstrated that human rhinoviruses can activate DCs, which then can produce and release IL-35 into human peripheral blood. Further, they showed that the induction of IL-35 was Foxp3 independent. Blocking receptors of CD274 (B7-H1) and CD169 (sialoadhesin) on DCs could prevent the induction of IL-35, which indicated that the induction of IL-35-induced Treg cells by human rhinoviruses depends on CD274 and CD169 receptors on DCs. Differences of stimuli, kinetics of stimulation, and the relative affinities of two chains are all factors affecting the production of cytokines (Collison and Vignali 2008).
Signaling Pathways
As we mentioned above, IL-12, IL-23, IL-27, and IL-35 are all composed by two subunits. One of the two mainly serves as a ligand, and the other should be a signal transmitter. IL-12 combines to its receptor complex, followed by activation of the Janus-associated kinase 2 (Jak2), tyrosine kinase 2 (Tyk2) and a series of the signal transducer activator of transcription (STAT) family members, such as STAT1, STAT3, STAT4, and STAT5. STAT4 was considered to play a critical role in this signal transmission. Some researchers found that these kinases and signal transducers were essential for IL-12 signaling (Bacon and others 1995; Gollob and others 1998; Mikus and others 2001; zbek and others 2005). The signaling pathway of IL-23 is similar to that of IL-12 (Presky and others 1996; Parham and others 2002). Jak2, Tyk2, and STAT family also are involved in IL-23 signaling, and STAT3 is believed to be a critical transcription factor (Lankford and Frucht 2003; Minegishi and others 2006; Mathur and others 2007; Tokumasa and others 2007; Yang and others 2007). IL-27 acts through different pathways in different cells. In naïve CD4+ T-cells, IL-27 signals via the activation of Jak1, Jak2, Tyk2, and STAT1–5 family (Hibbert and others 2003; Lucas and others 2003; Takeda and others 2003; Kamiya and others 2004). However, IL-27 mainly activates STAT1 and STAT3 in monocytes or macrophages (Pflanz and others 2004; H lscher and others 2005). In addition, in activated T-cells, STAT2 was the only factor to be activated (Yoshimura and others 2006).
So far, the signaling pathway of IL-35 is not clear yet. We could infer that the subunit EBI3 would combine with the gp130 chain as IL-27 does. The other subunit p35 would bond with the IL-12Rβ2 chain. From this perspective, the gp130 and IL-12Rβ2 chain would constitute the receptor of IL-35 (see Fig. 1). Meanwhile, the latest research performed by Collison and colleagues (2012) confirmed that IL-35 signaled through a unique heterodimer of receptor chains IL-12Rβ2 and gp130 or homodimers of each chain. Further, they believed that transcription factors STAT1 and STAT4 constitute a unique heterodimer bonding to distinct promoter regions of the p35 and EBI3 genes.
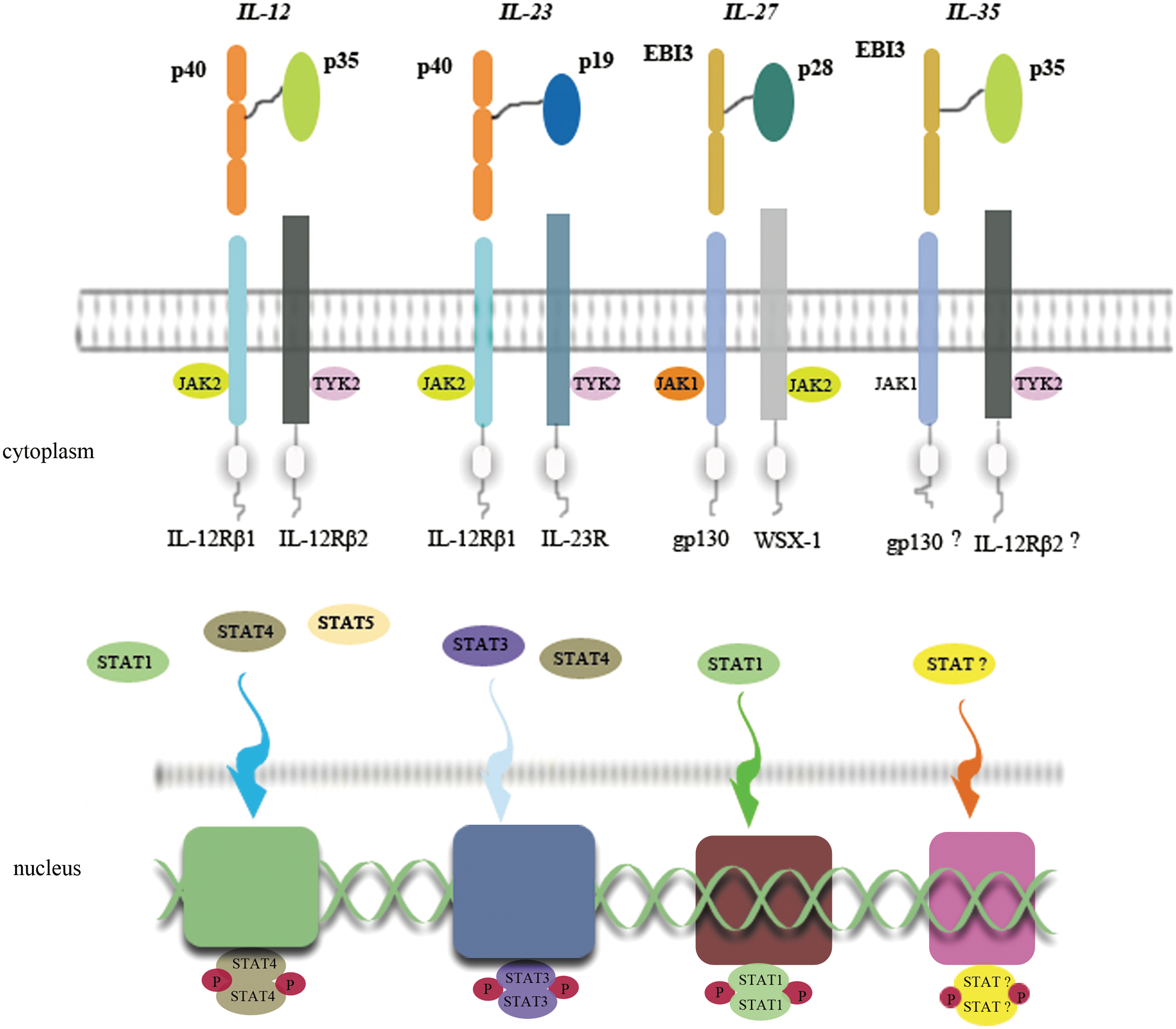
Signal pathways of IL-12 family members. IL-12 contains p40 and p35 subunits, which combine to IL-12Rβ1 and IL-12Rβ2, respectively, on the cell membrane. Then, JAK2 and TYK2 are activated. Phosphorylated STATs (especially STAT4) themselves form a dimer that translocates into the nucleus, and combines to a nucleotide, influencing the transcription. IL-23 and IL-27 do the similar way as IL-12 does. Such, we assume that IL-35 also has a similar signal pathway as the formers so. Two subunits of IL-35 EBI3 and p35 might attach to gp130 and IL-12Rβ2, respectively, activate JAK1 and TYK2, and translocate some kind of STAT into the nucleus. IL, interleukin; JAK2, Janus-associated kinase; TYK2, tyrosine kinase; STAT, signal transducer activator of transcription; EBI3, Epstein-Barr virus-induced gene 3.
Biological Function
The IL-12 family members have different dominant functions. IL-12p70 can induce the production of IFN-γ, promote the differentiation of T-helper 1 (Th1) cells (Trinchieri 2003), antagonize IL-4 secretion, and block Th2 cell responses, which can inhibit Th2-mediated diseases such as allergy and asthma (Sur and others 2000; Wills-Karp 2001). IL-23 also can induce IFN-γ production and favor the Th1 response and differentiation, though it is not absolutely necessary (Aggarwal and others 2003; Cua and others 2003; Murphy and others 2003; Kastelein and others 2007). According to previous reports, IL-27 can downregulate T-helper cell response, inhibit Th1-driven infections (Hamano and others 2003; Villarino and others 2003; Kastelein and others 2007), and block the development of Th17 (Neufert and others 2007). The newly found factor IL-35 can directly suppress Teff cell proliferation (Collison and others 2007), suppress Th17 development (Niedbala and others 2007), and mediate the proliferation of CD4+CD25+ T-cells and IL-10 generation. It is a brand new potential therapeutic cytokine for inflammation, autoimmunity, and other immunological disorders (Vojdani and Lambert 2009; Castellani and others 2010). Devergne and others (2001) showed that the two subunits of IL-35 were highly coexpressed in placental trophoblasts, which implied that IL-35 may have some immunological function at the fetomaternal border. Interestingly, IL-35 is functionally conserved between mouse and human, suggesting that mouse IL-35 would be a great replacement of human IL-35 in disease research (Li and others 2012).
IL-35 and Treg Cells
Treg cells can be classified as natural Treg cells and inducible Treg cells. Natural Treg cells are originated from the thymus (Shevach 2006). CD25 and transcription factor FOXP3 are generally expressed on natural Treg cells. Inducible ones are generated from T-cells under certain stimuli, such as IL-10 and transforming growth factor-β (TGF-β) (Shevach 2006; Tato and Cua 2008). IL-10 and TGF-β could induce their corresponding Treg cells: IL-10-iTR cells and TGF-β-iTR cells (see Fig. 2). Both of these iTR cells owned potential suppressive function, whatever in vitro or in vivo, (Groux and others 1997; Chen and others 2003; Grazia Roncarolo and others 2006) playing a critical role in maintaining immune balance. Collison and colleagues (2010) showed that IL-35 could induce naïve human or mouse T-cells to become regulatory T-cells, and they called them iTR 35 cells. These iTR 35 cells had a similar suppressive function as IL-10-iTR cells and TGF-β-iTR cells do. They also found that iTR 35 cells did not express or require the transcription factor Foxp3. Meanwhile, these iTR 35 cells were relatively stable in vivo. Ebi3−/− and Il12a−/− Treg cells had less regulatory activity and failed to control homeostatic proliferation and to cure inflammatory bowel disease in vivo (Collison and others 2007). The immune-suppressive function of Treg cells was primarily believed through a cell-contact-dependent manner (Takahashi and others 1998; Thornton and Shevach 1998; Tang and others 2005). Collison and others (2009) showed that not only Treg-cell-mediated suppression did depend on cell contact, but also some suppressive cytokines were also involved, especially IL-10 and IL-35.
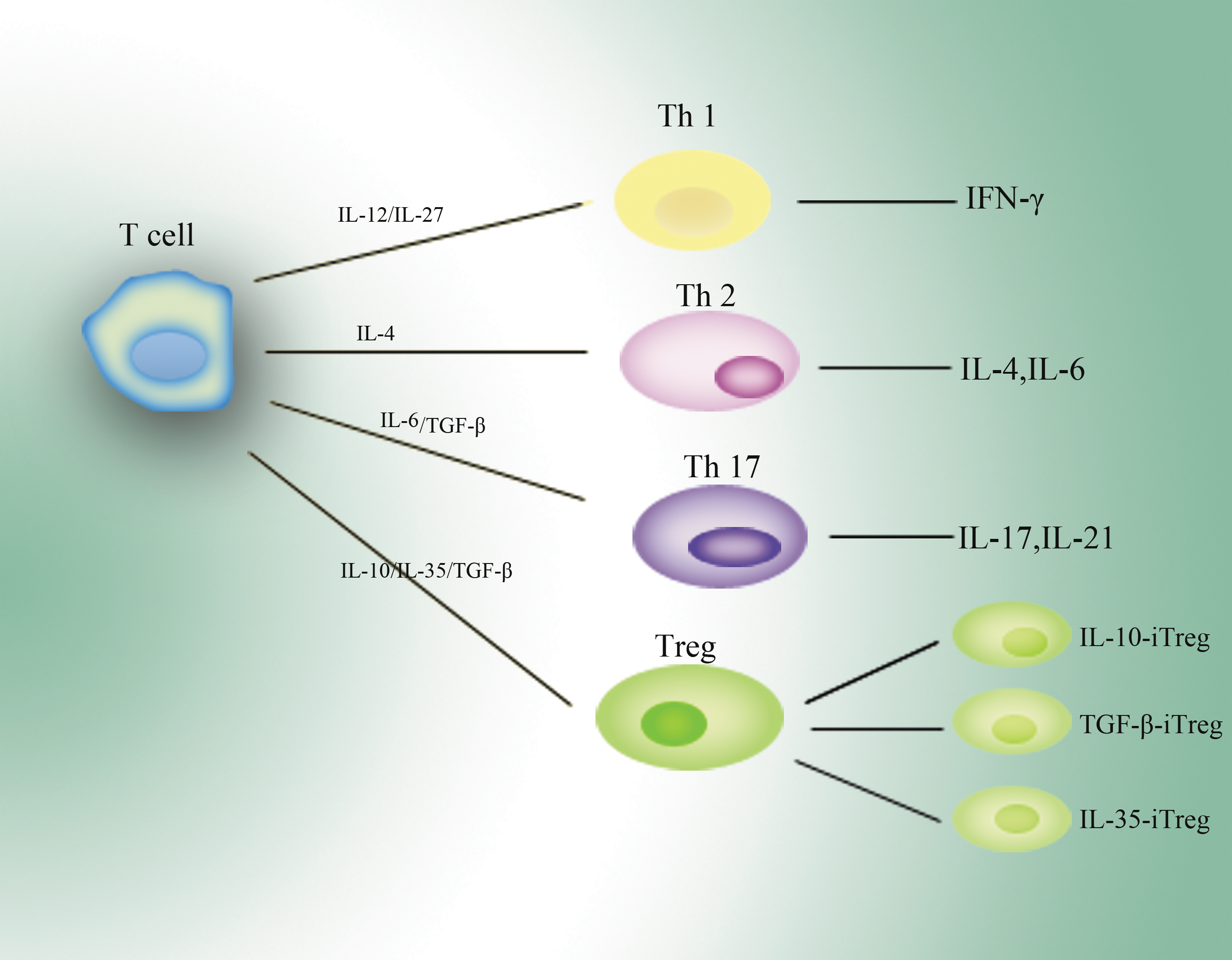
T-cells can be developed into different kinds of functional cells under various stimuli. IL-12 and IL-27 lead T-cells to become Th1 cells that secrete IFN-γ mainly. Similarly, Th2 cells and Th17 cells are developed from primary T-cells. IL-10, IL-35, and TGF-β can induce their specific Treg cells from primary T-cells. Th1, T-helper 1; IFN-γ, interferon-γ; TGF-β, transforming growth factor-β.
Chronic hepatitis C patients had a relatively high level of Treg cells that play an important role in maintaining immunoregulatory balance, and the inhibitory function of Treg cells depends on secretion of IL-10 and IL-35 (Langhans and others 2010). Treg cells were assumed to have therapeutic effects for rheumatoid arthritis (Boissier and others 2009), and IL-10, TGF-β, and IL-35 might play a great role in this process. IL-35 and Treg cells mutually upregulate each other. IL-35 can induce iTR 35 cells, and Treg cells can further produce IL-35.
Recently, Pillai and coauthors demonstrated that IL-10/IL-35 double-deficient Treg cells could maintain their suppressive function via increasing cathepsin E (Ctse) and other endogenous factors both in vitro and in vivo. Their results indicated that multiple regulatory pathways involved in suppressive function of Treg cells, and IL-35 was dispensable, though it was a great contributory factor.
Treg cells, especially its markers as CD25 and Foxp3, are necessary for the induction and maintenance of tolerance. Treg-derived cytokines as IL-10 and TGF-β are confirmed to have the feature to mediate infectious tolerance. Although the infectious tolerance feature of IL-35 has not been verified yet, we believe that it also exerts tolerant function during the process (Gravano and Vignali 2012). More recently, researchers revealed that IL-35, rather than IL-10 or TGF-β, was required in Treg-mediated suppression, which contributes to infectious tolerance (Chaturvedi and others 2011). The digger should go deeper to unearth the truth.
IL-35 and Inflammation
IL-35 involves in inflammatory diseases in the nervous system, alimentary system, bone and joint system, and respiratory system. Zandian and others (2011) established an HSV-IL-2-induced demyelination mouse model. They used different kinds of viruses expressing various cytokines or plasmid DNA to treat those mice. They found that IFN-γ exacerbated HSV-IL-2-induced demyelination, whereas IL-35 could suppress it. Their results indicated that IL-35 had an inhibitory effect against demyelination by preventing the development of autoaggressive T-cells. Liu and colleagues (2012) performed a study of IL-35 on patients with chronic hepatitis B (CHB). They showed that IL-35 could be detected in the peripheral blood of patients with CHB, whereas it was undetectable in healthy individuals. Their results suggested that IL-35 may play some kind of role during HBV infection. HBV might be a factor stimulating CD4+ T-cells to produce IL-35. Thus, IL-35 could provide a brand new perspective for the treatment of CHB (Devergne and others 1996). The subunit p35 is considered to be a critical role in restraining liver fibrosis (Tsuda and others 2012). Wirtz and others (2011) recently confirmed that IL-35, rather than IL-27, could significantly suppress Th1- and Th17-cells' proliferation, reduce the development of experimental colitis, and protect the intestine from immune responses in mice. Collison and coauthors (2010) showed that IL-35-induced Treg cells could contribute to the suppressive milieu, indicating that IL-35 could induce infectious tolerance and might facilitate tumor progression. Although IL-35 was constitutively expressed by mouse CD4+CD25+Foxp3+ regulatory T-cells, and could be detected in human placental extracts and chronic hepatitis patients' peripheral blood, it was not constitutively expressed by human CD4+CD25+Foxp3+ regulatory T-cells (Bardel and others 2008).
Kochetkova and colleagues (2010) used bovine elastin linker (Yamanaka and others 2008) to fuse mouse IL-12p35 subunit and mouse EBI3 together to produce IL-35. They demonstrated that exogenous IL-35 could suppress the activity of CD4+ T-cells, Th1, and Th17 cells. It could induce CD39+CD4+ T-cells and inhibit the inflammation of collagen-induced arthritis. They believed that these IL-35-induced CD39+CD4+ T cells were potent regulatory cells against collagen-induced arthritis. Niedbala and others (2007) also established a collagen-induced arthritis mouse model, and these mice were injected intraperitoneally with IL-35. They found that IL-35 could effectively attenuate collagen-induced arthritis. Further, their results showed that IL-35 could suppress the proliferation of CD4+CD25− effector cells as well as the differentiation of Th17 cells in vitro.
Meanwhile, in a smoke-induced lung inflammation rat model, IL-35 as well as CD4+Foxp3+ Tregs are increased after treatment. It meant that IL-35 could help the respiratory system recover from inflammation (Bai and others 2012; Whitehead and others 2012).
IL-35 and Autoimmune Disease
IL-35 also played a certain role in allergic diseases (Bullens and Ceuppens 2008). It controlled T-cell-driven immune responses. Expression of IL-35 in pancreas attenuated autoimmune diabetes (Bettini and others 2012). The EBI3 subunit played a potential inhibitory role in the development of experimental autoimmune encephalomyelitis (Liu and others 2012). Whitehead and colleagues (2011) suggested that IL-35, rather than IL-10 or TGF-β, could suppress IL-17 production and thereby reverse-established IL-17-dependent airway hyper-responsiveness in mice. Further, IL-35 was found to downregulate Blo t 5- (a major house dust mite allergen) induced allergic airway inflammation through the suppression of circulating Blo t 5-specific and total Immunoglobulin E (Huang and others 2011). More and more evidence supported the suppressive function of IL-35 in some allergic diseases.
There were also some controversies on the suppressive function of IL-35. Most researchers found that IL-35 could inhibit the immune response, attenuate inflammatory cascade, and protect organs and tissues. However, Kuo and others found that recombinant IL-35 would further exacerbate in Borrelia-vaccinated and Borrelia-infected mice, while the anti-IL-17 antibody-treated mice had little tissue damage. This result might induce further discussion on the function of IL-35 on the immune system. Moreover, to clarify it, we should do more work on the signaling pathways of IL-35.
Conclusion and Perspective
We did an exhaustive review on the newly identified cytokine IL-35. The finding of IL-35 was of great significance, since it could regulate T-cells that were important therapeutic targets of immunological disorders. Although targeting T-regulatory cells is of great potential for the treatment of immune diseases, there is little understanding about suppressive factors. Meanwhile, IL-35 just serves as a suppressive factor in the immune system. The interaction between IL-35 and T-regulatory cells would be a critical process in the immune reaction. Therefore, the regulation of IL-35 might be of great significance in the immune response.
The exact mechanism of IL-35 regulating the T-cells is still unknown. If IL-35 served as a major influential factor of T-regulatory cells, inhibition of IL-35 could amplify the immune response that might benefit lots of people. Currently, a great number of people cannot get a response to hepatitis B vaccine no matter how many times they receive injection. T-cells, especially regulatory T-cells, may have a low immune response within these people. We are perspective that inhibition of IL-35 could elevate the vitality of T cells, which would make the antibody positive. The effect of other vaccines would also be strengthened. Even a vaccine for special cancer would show up soon. It provides a brand new therapeutic way for some immunocompromised diseases, such as AIDS. Autoimmune diseases and inflammation are caused by the disorder of immune regulation, and IL-35 would also provide a completely different way for the treatment. It is a good news for patients who suffer from autoimmune diseases.
In conclusion, IL-35 has a great potential in the immune system, though researches on it just take off. Its particular biological function may promote the development of the vaccine and cure inflammatory diseases. Its signaling pathway and further biological function still need to be clarified.
Footnotes
Acknowledgments
This work was supported by the grants from the National Natural Science Foundation of China (No. 30972946), and the Zhejiang Provincial Natural Science Foundation of China for Distinguished Young Scholars (No. R2110125).
Author Disclosure Statement
No competing financial interests exist.