
Abstract
A balanced proinflammatory cytokine response to Plasmodium ssp. infection is crucial to control the disease outcome. To elucidate the effect of cytokines and Plasmodium falciparum-infected erythrocytes on the regulation of interleukin (IL)-6 receptor (IL-6R), ciliary neurotrophic factor alpha (CNTFR-α) and glycoprotein (gp)130 in natural killer (NK) cells in the context of malaria, we assessed their gene expression and surface expression in NK92 cells. P. falciparum alone did not alter gene expression of the investigated receptors in NK92 cells. Analysis revealed a low effect of IL-6 on IL-6R surface expression in NK92 cells. However, at transcriptional level, a downregulation of IL-6R was observed following IL-6 stimulation. Thus, IL-6 might act within a negative feedback loop to terminate signal transduction by downregulating IL-6R expression. Additionally, we observed that IL-6R and CNTFR-α surface expression were regulated by a combination of IL-2, 12, and 18, and gp130 was influenced by interferon-α. Our results show that the IL-6 family receptors in NK92 cells are not directly influenced by P. falciparum. However, cytokines usually derived from accessory cells during malaria episodes may regulate IL-6 receptor signaling pathways. This finding encourages future studies in a more physiological context and with primary cells isolated from humans with and without malaria.
Introduction
N
IL-6 belongs to a family of cytokines that stimulate NK cells. Besides that it has a variety of other functions and is involved in processes like hematopoesis, regulation of immune responses and acute-phase reactions. Its receptor IL-6R is expressed in many tissues on cell types such as bone marrow cells, hepatocytes, peripheral blood mononuclear cells (PBMCs), as well as leukemic blasts (Wognum and others 1993). IL-6 belongs to the helical neuropoetic IL-6-family, which comprises IL-6, IL-11, leukemia inhibitory factor, oncostatin M, ciliary neurotrophic factor (CNTF), cardiotrophin-1, cardiotrophin-like cytokine (Muller-Newen 2003), neuropoietin, and IL-27 (Demyanets and others 2012). These cytokines are involved in the modulation of inflammation, immune responses, heart development, and fertility, among other processes. All members share the same common β-receptor CD130, which associates with the respective α-receptor (IL-6Rα, CNTFR-α, etc.) that does not transduce signals by itself (Muller-Newen 2003). While expression of the associated specific α-receptors is strictly regulated (Hirano and others 1997), glycoprotein (gp)130 is found almost ubiquitously in tissues (Oberg and others 2006). The signal-transducing receptor was described initially in mice as a membrane-bound glycoprotein termed gp130, absent on resting lymphocytes, but present on T-cell blasts (Andersson and others 1978). The receptor consists of a cytoplasmic region, a single transmembrane segment, and an extracellular domain (Muller-Newen 2003). After cytokine binding to its α-receptor, gp130 forms homo- or heterodimers and becomes phosphorylated by noncovalently bound receptor-associated Janus kinases (Jak1, Jak2, or Tyk2) at specific tyrosine residues of the cytoplasmic domain. IL-6-related cytokines preferentially signal via signal transducers and activators of transcription (STAT) 1 and 3 (Chevalier and others 1996), or additionally STAT5 (Hirano and others 1997).
Cytokines play an important role in immune responses to Plasmodium falciparum. In severe malaria, production of proinflammatory cytokines such as IFN-γ, IL-12, IL-6, and tumor necrosis factor (TNF)-α is crucial for activation of immune responses in early stages of the infection, but later on, their levels correlate with disease severity and may contribute to the systemic and organ-related complications (Kwiatkowski 1990; Kwiatkowski and Nowak 1991; Schofield and Grau 2005; Clark and others 2006). Excessive production of TNF and IL-6 can enhance the expression of adhesion molecules on endothelial cells, elevate parasite cytoadhesion, and contribute to host tissue damage (Mackintosh and others 2004). In Mozambican children suffering from severe malaria, IL-6 levels were significantly increased compared to uncomplicated cases (Rovira-Vallbona and others 2012). In another cohort form children residing in Papua New Guinea, IL-6 production by mainly monocytes/macrophages and γδ-T cells after infection with P. falciparum was correlated with increased clinical episodes, whereas TNF-α and IFN-γ reduced the number of episodes (Robinson and others 2009). Several clinical manifestations such as periodic fever, rigors, and chills are caused by peak levels of proinflammatory cytokines produced by the cells of the innate immune system in response to high doses of parasite components released during schizont burst (Kwiatkowski 1990; Kwiatkowski and Nowak 1991). Analysis of plasma levels of patients revealed that in those with cerebral malaria or kidney failure, IL-6 and IL-6R levels were significantly higher compared to control groups. It was hypothesized that reduction of IL-6R after therapy and low CNTF expression cause cerebral malaria or renal failure (Wenisch and others 1999).
Since soluble receptors of IL-6 are present in the blood, and given the ubiquitous expression of gp130, it seems that virtually all cells can respond to IL-6. In mice, IL-6 trans-signaling through the soluble IL-6R has been demonstrated to be involved in malaria-dependent lethality (Wunderlich and others 2012). It has been shown that gp130, present on both hematopoietic and nonhematopoietic cells, varies significantly among different PBMC subpopulations depending on the activation status of the cells. Some studies showed absence of gp130 in resting NK cells (Chevalier and others 1996; Schofield and Grau 2005; Clark and others 2006). However, a role in NK cell activation measured as presence of CD54 and CD69 in response to IL-27 secreted by DCs could be assigned for gp130 (Crabé and others 2009).
It is likely that NK cells are important during early immune responses to Plasmodium ssp. infection. However, little is known about the impact on expression of receptors of the IL-6-family in this cell type in the context of malaria and also apart from that. These observations led us to investigate whether NK cells can be stimulated to gp130, IL-6R, and/or CNTFR-α expression directly by P. falciparum-infected erythrocytes or by cytokines usually provided by accessory cells during infection. To test our hypothesis, we employed NK92 cells as a repeatable model system.
Materials and Methods
NK cells
NK92 cells (DSMZ) were grown in 75% alpha-MEM supplemented with 12.5% fetal bovine serum and 12.5% horse serum, 2 mM
P. falciparum culture
The P. falciparum strains 3D7 and FCR3-CSA were maintained in continuous culture as described previously (Trager and Jensen 1976). Parasites were grown in 0+ human erythrocytes (Blood bank, University Hospital Tübingen, Germany) in RPMI 1640 supplemented with 25 mM HEPES,
NK92 activation with different stimuli
To exclude prior activation, NK92 cells were kept in a cell medium without rIL-2 overnight. NK92 cells were either cocultured with schizont-infected red blood cell (iRBC) and uninfected red blood cell (uRBC) (1:3, NK cell to RBC) or stimulated with IFN-α (500 U/106 cells), IL-6 (500 U/106 cells), or a mix of IL-2/-12/-18 (10 ng/mL IL-2, 100 ng/mL IL-12, and IL-18) for 1 or 24h. After the indicated time of incubation at 37°C and 5% CO2, lysis buffer (BD Biosciences) was added to all cultures to lyse erythrocytes. FCR3 parasite strain was selected for control experiments.
Cell surface staining for flow cytometry
Cells were stained for 30 min at 4°C in the dark with fluorochrome-conjugated antibodies to the surface markers CD56 (FITC), CD3 (PE), CD130 (PE), CD126 (PE), and PE isotype control (BD Biosciences). Cells stained in the dark with mouse anti-human CNTFR-α (1 μg IgG2a/106 cells) for 20 min on ice were washed twice with FACS buffer [phosphate-buffered saline (PBS), 1% fetal bovine serum, and 0.1% sodium azide] and then stained in the dark with goat anti-mouse IgG2a-FITC (1 μg IgG2a/106 cells; Santa Cruz) for another 20 min on ice. Dead cells were excluded by 7-AAD staining performed directly before measurements. Flow cytometry measurements were carried out using a FACSCanto flow cytometer (BD Biosciences) and analyzed with BD FACS Diva 6.0 software. Gating was done based on forward and sideward scatter. A total of 10.000 events were collected from each sample.
Real-time quantitative analysis of gp130, CNTFR-α, and IL-6R expression
Total RNA was extracted from 3×106 cells with an RNeasy Mini Kit (Qiagen) according to the manufacturer's instructions. 0.5 μg RNA was converted into cDNA using a Quantitect Reverse Transcription Kit (Qiagen), which includes a DNA elimination step before reverse transcription. For real-time polymerase chain reaction (RT-PCR), the following pre-established Quantitect Primer Assays (Qiagen) were used: QT00091294 (gp130), QT00022505 (CNTFR-α), QT00023660 (IL-6R), and QT01192646 (β-actin). The reaction mix was prepared according to the standard protocol of the Rotor-Gene SYBR Green RT-PCR Kit (Qiagen). RT-PCR was carried out with Rotor Gene 3000 Cycler. The thermal profile was set as follows: 5 min at 95°C for initial denaturation, 40 cycles of 95°C for 10 s, and 60°C for 15 s with a final amplification step at 60°C for 7 min. To exclude contamination, a nontemplate as well as a water control was run in parallel to each performed experiment. Specificity of products was verified by melting-curve analysis and agarose gel electrophoresis. Measurements were done in triplicate. For analysis of the obtained amplification curves, the ΔΔCT-method was applied. CT-values above 30 were excluded, and a threshold of 0.01 was set. RNA was isolated after 24 h and reverse-transcribed. RT-PCR was performed for the gene of interest and actin as housekeeping gene. Fold changes were calculated in comparison to untreated cells.
Statistical analysis
Analysis was performed using JMP for Windows 5.0.1 (SAS Institute, Inc., Cary, NC) running on Windows XP (Microsoft Corp., Redmond, WA). The level of significance was set at P value <0.05.
Results
NK92 upregulate transcription and surface expression of gp130 after IFN-α stimulation
To detect surface expression of gp130, NK92 cells were analyzed by flow cytometry. NK92 cells grown in a culture medium [not activated (NA)] were compared to NK92 cells stimulated either with a mix of IL-2, IL-12, and IL-18 (IL-mix), with IL-6, IFN-α, uRBC, or P. falciparum-iRBC for 24 h.
After incubation for 24 h in a growth medium without IL-2 (NA), between 0.8 and 4.4% of NK92 cells expressed gp130 (mean=2.2%±1.5%, n=4). The gp130 expression levels did not increase with IL-mix (mean=3.6%±2.7%, n=4), IL-6 (mean=4.1%±3.9%, n=4), uRBC (mean=6.1%±6.5%), or iRBC (5.4%±4.7%, n=4). However, after IFN-α stimulation, 14.6%±8.2% of NK92 cells (n=4) were stained positive for gp130 surface expression (Fig. 1). In summary, surface expression of the β-receptor gp130 was significantly upregulated upon stimulation with IFN-α compared to NA (Fig.1, paired Student's t-test, n=4, P<0.05). All other stimuli did not activate gp130 surface expression significantly.
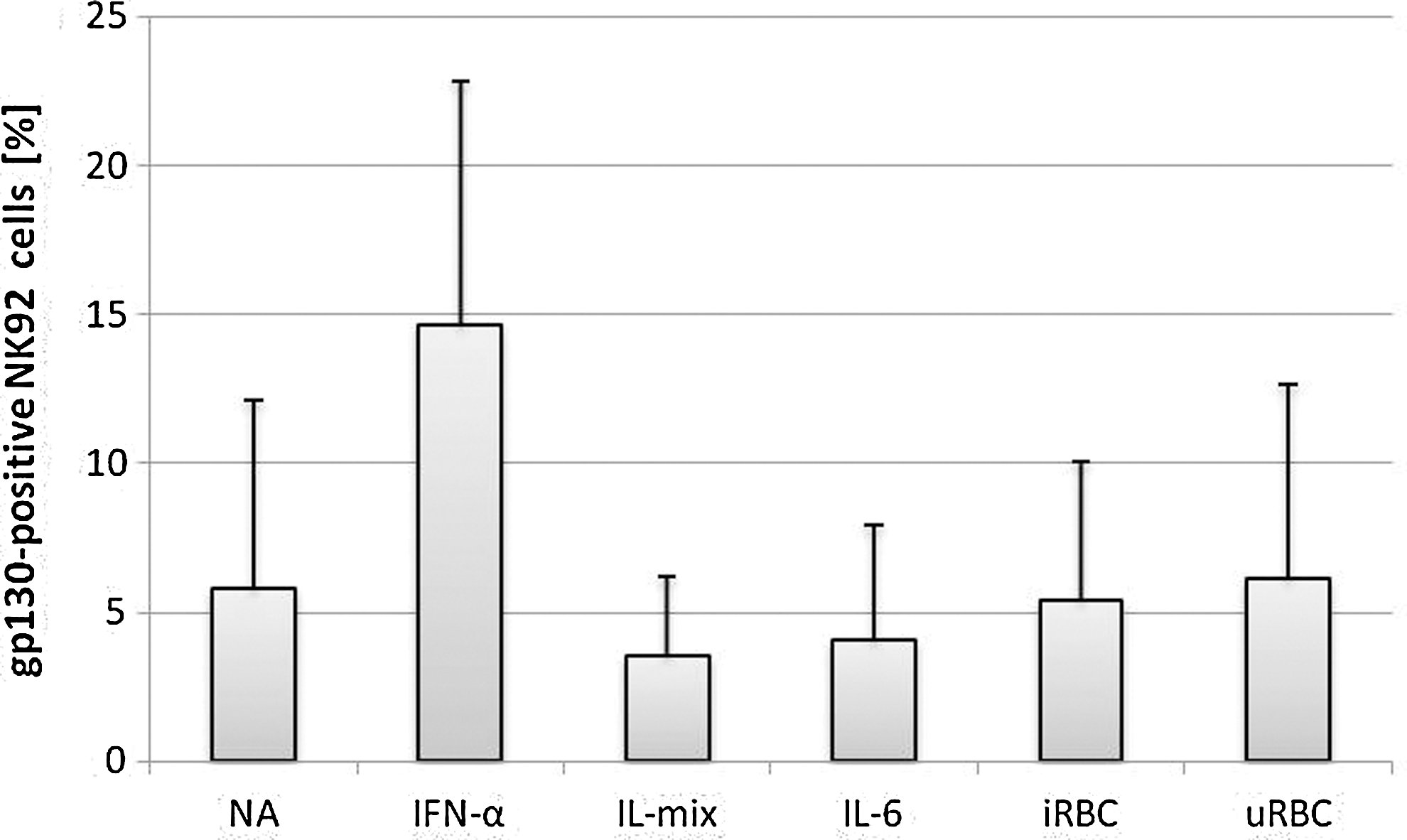
Surface expression of gp130 on NK92 cells after 24 h of stimulation. NK92 were cocultured with infected/uninfected erythrocytes in a 1:3 ratio or stimulated with interleukin (IL)-6 (500 U/mL), interferon (IFN)-α (500 U/mL) or a mixture of IL-2 (10 ng/mL), IL-12 (100 ng/mL), and IL-18 (100 ng/mL). Surface expression was analyzed by flow cytometry and compared to untreated cells [not activated (NA)]. Experiments were done in triplicate or quadruplicate.
Since a rapid effect of IL-6 stimulation is possible, levels of gp130 surface expression were determined also after 1 h of stimulation with cytokines or erythrocytes; gp130 was upregulated after 1 h of stimulation with IL-6, but not with IL-2/-12/-18 (IL-mix), uRBC, or iRBC. After IFN-α stimulation, NK cells show a tendency to express more surface gp130 compared to control cells. However, this upregulation turned out to be not significant (data not shown). The results on parasite-induced gp130 expression were confirmed using RBCs infected with the FCR3 parasite strain that was selected for their property to bind to chrondroitinsulfate A (CSA) for stimulation of NK92 cells (data not shown).
To verify whether the detected effects would also be reflected at transcriptional level, RNA was extracted from 24 h stimulated and unstimulated NK cells and reverse-transcribed. Expression of gp130 was compared to the housekeeping gene β-actin by qRT-PCR. As shown in Table 1, mRNA levels were similar after 24 h stimulation with different ILs (IL-mix=1.56±0.39-fold and IL-6=1.34±0.08-fold). After incubation with iRBC, a 2.13±0.23-fold upregulation was detected. However, uRBCs increased gp130 expression 2.44±0.18-fold as well. For IFN-α, a 2.30±0.20-fold increase was observed. These results confirm on the one hand results obtained by flow cytometry, as 24-h incubation with IFN-α, but not with IL-6, led to higher expression of gp130 in NK cells. On the other hand, a small effect of uRBCs could be observed, which was not dependent on infection with P. falciparum.
NK92 cells were incubated with the following stimuli: IL-mix: IL-2 (10 ng/mL), IL-12/-18 (100 U/mL); IL-6: 500 U/106 cells; IFN-α: 500 U/106 cells; u/iRBC: un/infected erythrocytes (1:3 NK:RBC). After 24h of stimulation, transcript levels of gp130 were analyzed by quantitative RT-PCR.
IFN, interferon; IL, interleukin; uRBC, uninfected red blood cell; iRBC, infected red blood cell; NK, natural killer.
Upregulation of CNTFR-α on the surface of NK92 after IL stimulation
In contrast to gp130, CNTFR-α was significantly upregulated on the NK92 surface after 24 h stimulation with IL-mix (50.13%±23.15%, n=3, P<0.05) compared to nonstimulated control cells (16.43%±5.58%, n=3) and iRBC-stimulated cells (14.30%±8.32%, n=3). IL-6 and IFN-α also induced surface expression of CNTFR-α however, the increase was not significant compared to nonstimulated controls (IL-6: 42.43%±20.83%, n=3; IFN-α: 30.87%±23.33%, n=3). Upon stimulation with P. falciparum-iRBC, 14.3% were stained positive, while for the uRBC control 23.50%±12.80% were positive for CNTFR-α surface expression (Fig. 2).
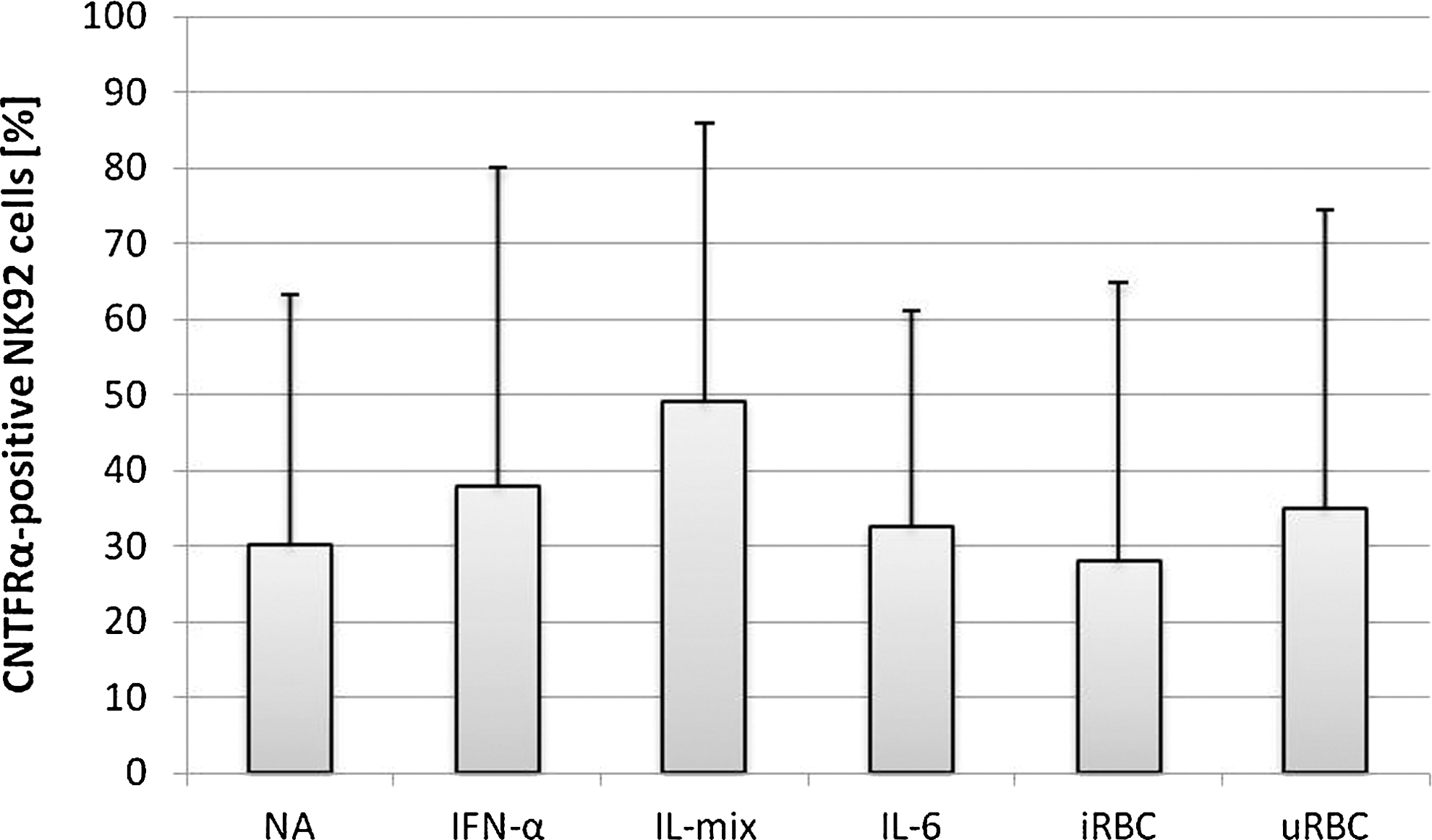
Surface expression of ciliary neurotrophic factor alpha (CNTFR-α) on NK92 cells after 24 h of stimulation. NK92 cells were cocultured with infected/uninfected erythrocytes (1:3) or stimulated with IL-6 (500 U/mL), IFN-α (500 U/mL) or a mixture of IL-2 (10 ng/mL), IL-12, and IL-18 (100 ng/mL each). Surface expression was analyzed by flow cytometry and compared to untreated cells (NA). Experiments were performed in triplicate.
IFN-α induces significant upregulation of CNTFR-α transcription levels, but does not increase surface expression
As stated above, flow cytometry analysis of 24h-stimulated NK92 cells demonstrated no major changes in surface expression of CNTFR-α. However, gene expression levels between IFN-α-stimulated NK cells were significantly higher compared to NK cells incubated with IL-2, 12, and 18 or iRBC. Surprisingly, uRBCs (2.2-fold change, n=4, P<0.01) had a significant influence on CNTFR-α transcription. NK cells stimulated with IL-mix for 24 h reduced transcription almost 4-fold (n=3, mean=0.28, P<0.01). Transcription levels decreased after challenge with iRBCs (n=3, mean=0.41, P<0.01, Fig. 3).
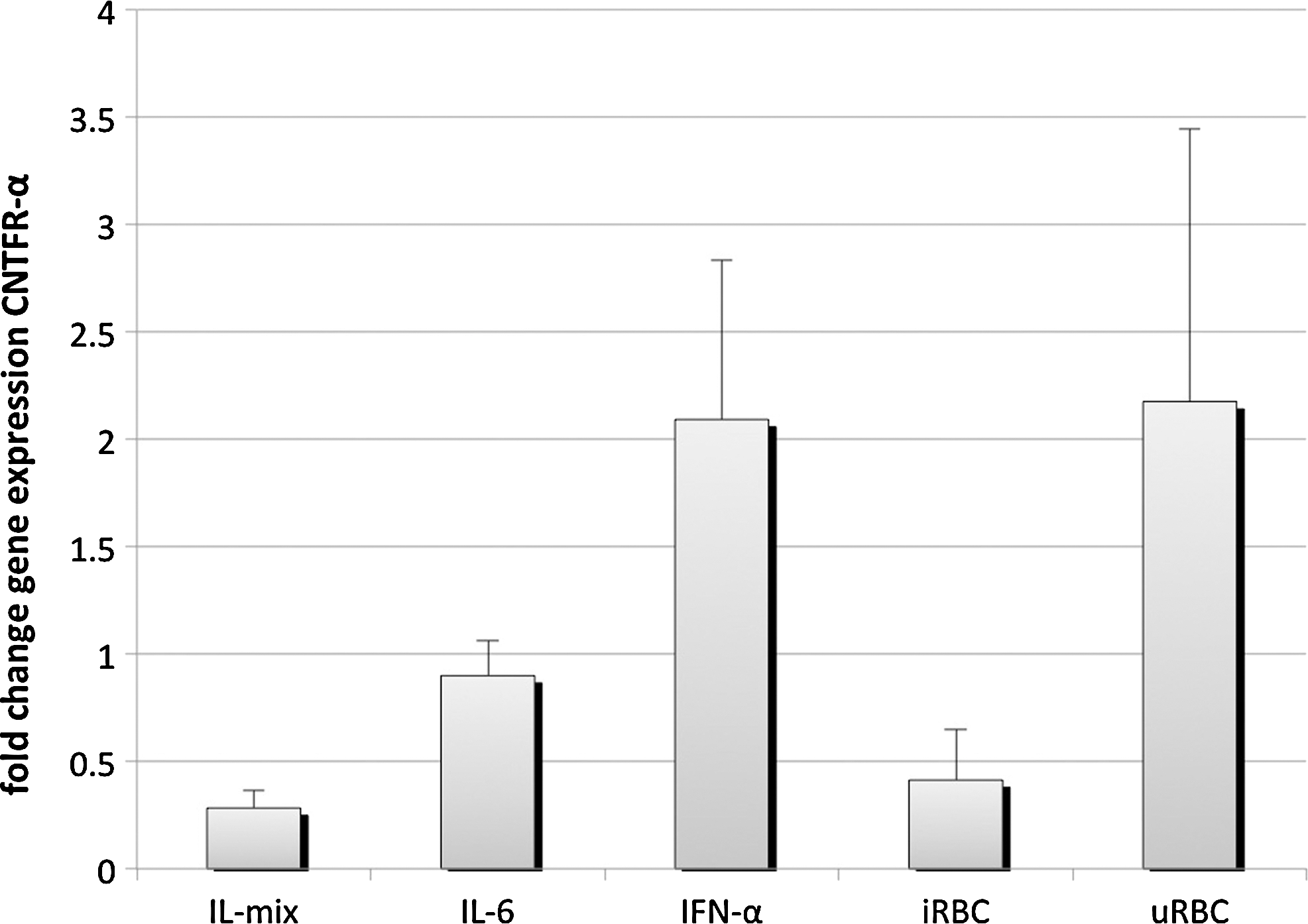
Altered gene expression of ciliary neurotrophic factor alpha (CNTFR-α) in NK92 after stimulation with P. falciparum-infected red blood cell (iRBC) or cytokine stimulation. NK92 cells were either cultured in a growth medium alone and incubated with a mixture of IL-2 (10 ng/mL), IL-12 (100 ng/mL) and IL-18 (100 ng/mL) or cocultured with u/iRBC (ratio 1:3). RNA was isolated after 24 h and reverse-transcribed. Real-time polymerase chain reaction (RT-PCR) was performed for the gene of interest CNTFR-α and β-actin as housekeeping gene. Fold changes were calculated in comparison to untreated cells applying the ΔΔCT method. Results are depicted as mean fold change±standard deviation and are representative for 3 individual experiments performed with technical duplicates each.
IL-2, 12, and 18, but not IL-6 change surface expression of IL-6R
Similar to CNTFR-α, surface expression of IL-6R was only significantly altered after 24-h stimulation with IL-mix containing IL-2, 12, and 18. Again, a slight trend to higher surface expression was detected after 24-h stimulation with IFN-α. Surprisingly, neither IL-6 after 1 and 24 h nor iRBC resulted in altered surface expression of the α-receptor on NK92 cells (Fig. 5). This could be an effect of constitutive expression of IL-6R on NK92 cells, so that no further upregulation is required for exerting IL-6 signaling.
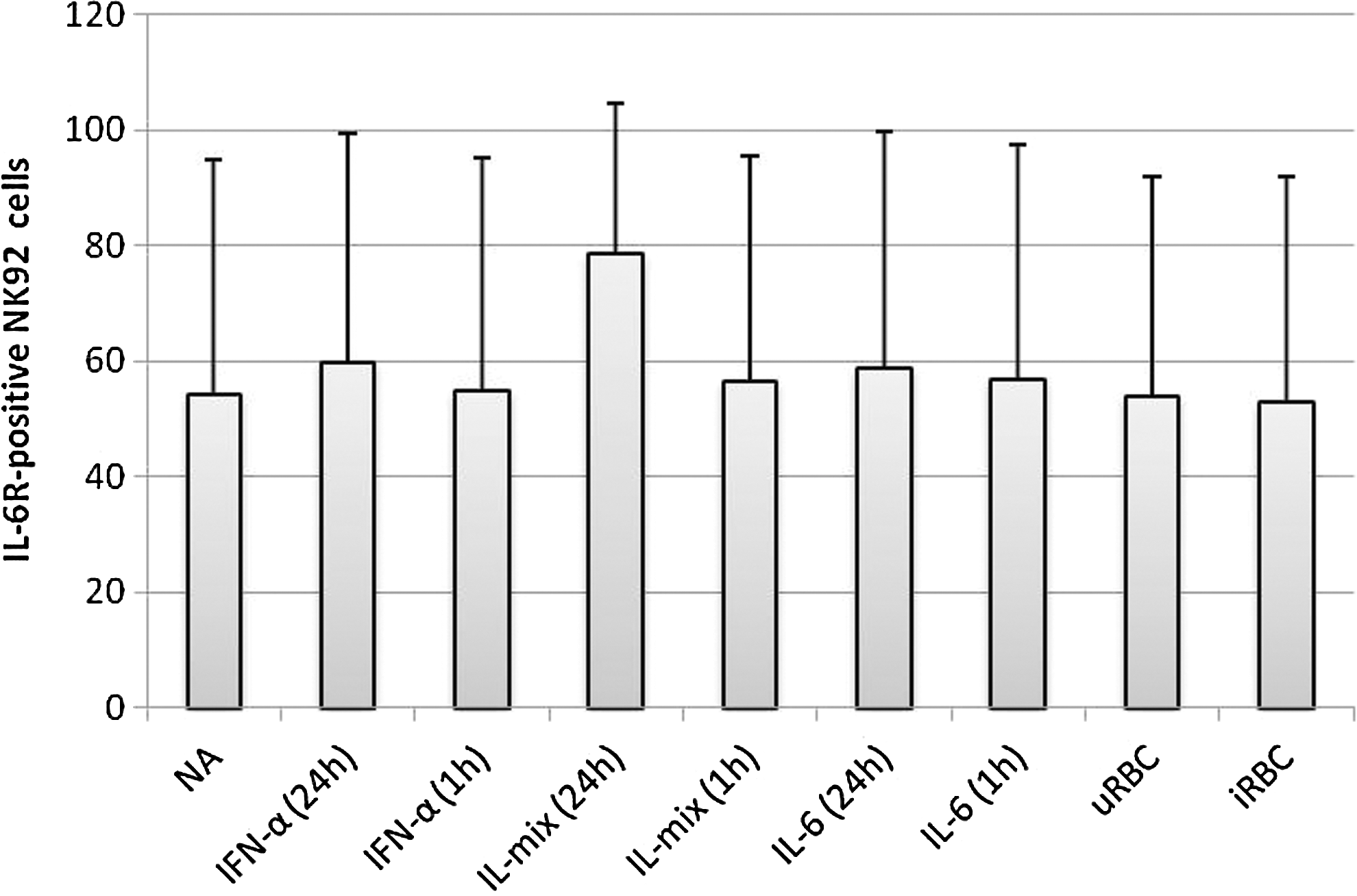
Influence of cytokine and P. falciparum stimulation on the surface expression of IL-6R in NK92 cells after 1 and 24 h of stimulation. Surface expression of IL-6R was assessed by flow cytometry compared to cells that were not stimulated (NA). NK92 cells were either cultured in a growth medium alone (NA), incubated with a mixture of IL-2 (10 ng/mL), IL-12 (100 ng/mL) and IL-18 (100 ng/mL) (IL-mix) or cocultured with u/iRBC (ratio 1:3).
Transcription levels of IL-6R are reduced after cytokine and RBC stimulation
Determination of changes in transcription revealed that with all the given stimuli, IL-6R expression was diminished, ranging from 1.85-fold after 24 h coculture with iRBC or IFN-α stimulation up to 9-fold downregulation after IL-6 treatment (Fig. 4). The strongest effect was observed for IL-6, while also stimulation with IL-mix yielded in 3.2-fold down modulation of transcription levels of IL-6R.
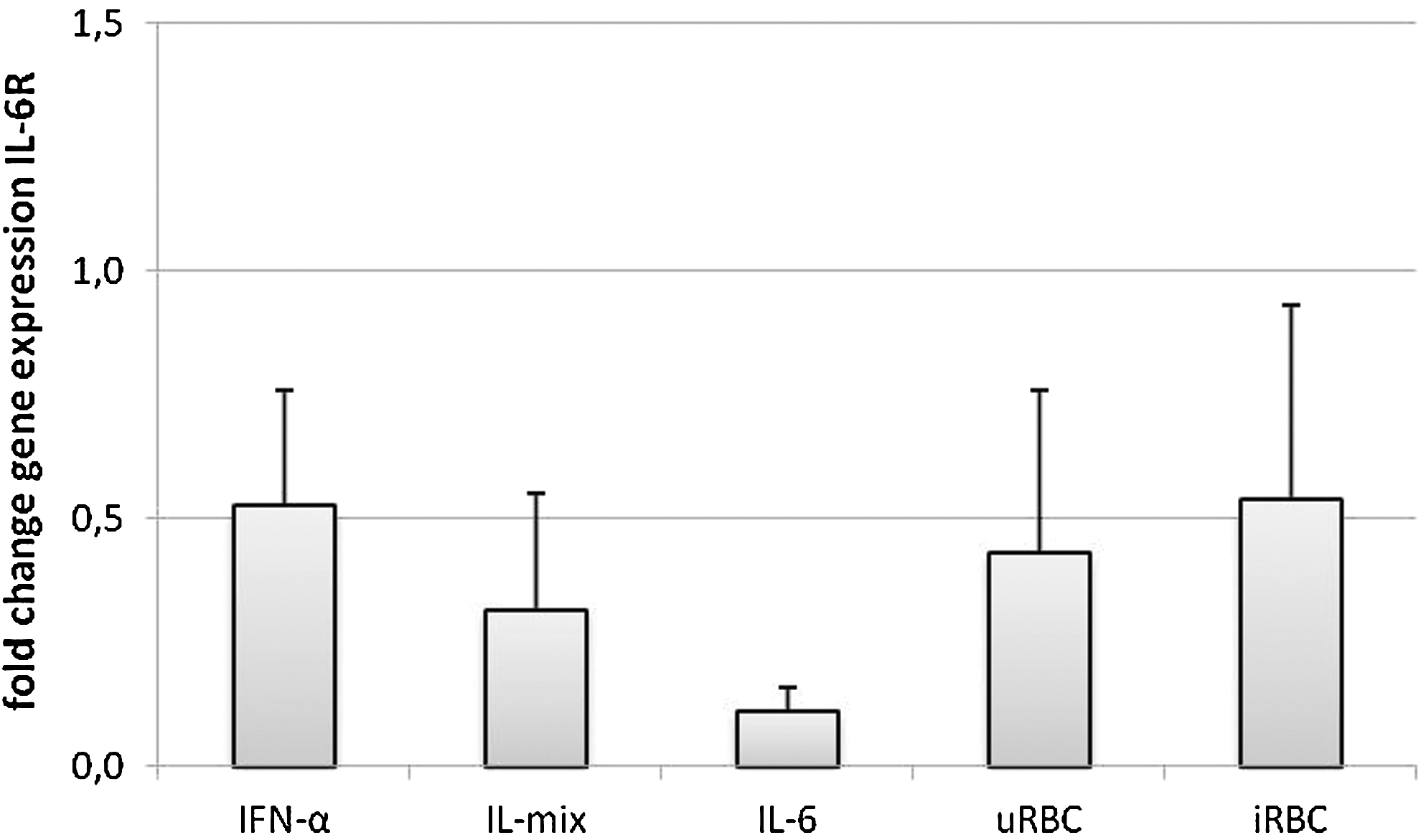
Influence of cytokine and P. falciparum stimulation on gene expression of IL-6R in NK92 cells after 24 h of stimulation. NK92 were cocultured with infected/uninfected erythrocytes in a 1:3 ratio or stimulated with IL-6 (500 U/mL), IFN-α (500 U/mL), or a mixture of IL-2 (10 ng/mL), IL-12 (100 ng/mL), and IL-18 (100 ng/mL). After 24 h, RNA was isolated and reverse-transcribed. Real-time polymerase chain reaction performed for the gene of interest IL-6R and β-actin as housekeeping gene. Fold changes were calculated in comparison to untreated cells applying the DDCT method. Results are depicted as mean fold change – standard deviation and are representative for 3 individual experiments performed with technical duplicates each.
Discussion
In the present study, we investigated the influence of different cytokines, uRBC, as well as iRBC on NK cell surface expression and transcription of IL-6 family receptors, namely gp130, CNTFR-α, and IL-6R. We conclude that uRBC and iRBC have little influence on expression of IL-6 family receptors in NK92 cells, thereby demonstrating that NK cells do not react to iRBC by upregulating receptors of the IL-6 family. Moreover, IL-6R and CNTFR-α were slightly downregulated, however, to a lesser extent than after cytokine stimulation. In our experiments, we used iRBCs containing late-stage parasites to stimulate NK92 cells, which may lead to erythrocyte breakdown and release of merozoites during the course of the experiment. It is known that schizonts can activate NK cells (Wu and others 2010), and iRBC burst is associated with peak levels of proinflammatory cytokines and fever (Kwiatkowski 1990). Interestingly, in our experiments, incubation with nonautologous uRBCs seem to influence expression patterns of some receptors as well. An uRBC control therefore appears to be crucial for correct interpretation of results. This effect was also observed in other studies investigating the influence of P. falciparum on NK cells (unpublished data).
Secondly, we show that the investigated α-receptors of the IL-6 family are similarly regulated by several cytokines. Since a proper balance of proinflammatory and anti-inflammatory cytokines seems to influence the disease outcome, we were interested to study the effects of several proinflammatory cytokines on NK cells. Our observations suggest that IL-6 acts in an autoregulatory manner, down-modulating the induced signaling cascade through negative feedback mechanisms. Except for IL-6R transcription, IL-6 showed a surprisingly minor effect, while IFN-α and the IL-mix had greater impact. The IL-mix increased surface expression, but decreased cytosolic abundance and transcription. It appears that if signaling events are transduced via IL-6R, the signal is terminated already before 24 h. This may point to a negative regulation by IL-2, IL-12, and IL-18 to curtail the provoked signal. If a single cytokine or the combination of all 3 leads to the measured effects remains to be determined. It is most likely that IL-12 might play a key role as cross talk with IL-12 family cytokines was reported for IL-6 signaling (Garbers and others 2012).
Conflicting results exist regarding IL-6R expression in NK cells. Oberg and others (2006) describe the complete lack of IL-6R and gp130 on resting NK cells of healthy donors. The same was reported by a different study that detected IL-6 receptor expression on NK cells in normal blood of rhesus monkeys (Wognum and others 1993). In contrast, we were able to detect IL-6R on the surface of NK92 cells either stimulated or not. This could be due to our approach of using a cell line, which is different from ex vivo isolated resting NK cells from healthy donors (de Carvalho and others 2011). NK92 cells already produce high amounts of IFN-γ and express the activation marker CD25 on their surface under normal culture conditions, which argue for some preactivation (de Carvalho and others 2011). Furthermore, the lack of IL-6R detection could be an effect of low template amounts for RT-PCR, since we had to adjust to proper starting quantities to obtain amplification. Apart from that, Rabinowich and others (1993) also reported response of NK cells to IL-6. Additionally, there are some reports demonstrating that IL-6 stimulates NK activity of human CD3 large granular lymphocytes in the absence of T cells (Luger and others 1989; Smyth and Ortaldo 1991) and LAK activity of IL-2-cultured human PBMC (Gallagher and others 1990). In our study, IL-6 strongly downregulated IL-6R gene expression. There is reported evidence from human hepatoma cells (HepG2) that IL-6R surface expression is downregulated by its ligand IL-6. Especially, the low-affinity-binding subunit gp80 was demonstrated to internalize the ligand within 1 h and degrade it after 4 h. The authors also reported de novo synthesis of IL-6R within a prolonged incubation time up to 24 h (Zohlnhöfer and others 1992).
In our experiments, all cytokine stimuli resulted in a higher gene expression of CNTFR-α, although not leading to a higher surface expression. We suppose that the transport to the cell surface might be differently regulated for the applied stimuli.
Because commercially available antibodies for gp130 and IL-6R to perform Western blot experiments did not detect those receptors in our experiments, we could not assess protein amounts by immunoblot. Overall, the tested cytokines showed only a slight influence on transcription levels of gp130 and CNTFR-α. Since IL-6 signals are regulated via gp130, this result was surprising for IL-6. Cytokines mainly induced either up- or downregulation of ∼2-fold, what is barely above a background level of 1.5-fold change of gene expression. In the present experiments, we tested only 2 different time points (1 and 24 h). Therefore, we could have missed the time of peak difference. It would be interesting to perform a time kinetics experiment. However, we chose 24 h, since proper NK cell activation measured as IFN-γ production and upregulation of activation markers CD25 and CD69 is usually achieved after this period.
In summary, we gained new insights into the regulation of 3 different receptors of the IL-6 family in NK cells after 1 and 24 h of stimulation induced by cytokines that may be important during malaria infection. Apparently, IL-6 itself has no important effect on surface expression of gp130, IL-6R, and CNTFR-α. Moreover, it rather triggers termination of induced signaling events by downregulating IL-6R gene expression. Furthermore, the nonsignaling α-receptors were altered by a mixture of IL-2,-12, and -18, which are activating cytokines for NK cells derived in vivo from accessory cells during early malaria infection. Additionally, the signal-transducing β-receptor gp130 was not influenced by IL-6 but by IFN-α. P. falciparum-iRBC alone could not stimulate NK cells to modulate the investigated receptor expression, suggesting that if these receptors play a role in NK cell responses during malaria, regulation is indirect through the action of accessory cells. To complete the picture, it could be interesting to assess if activation of IL-6 family receptors is diverse at different time points. However, our focus was to investigate effects elicited by P. falciparum on NK cells. Therefore, an incubation time of 24 h seemed most interesting, as maximal NK cell response is reported to take place during 12–24 h after contact to iRBCs (Artavanis-Tsakonas and Riley 2002). The proinflammatory ILs act before IFN-α by upregulating the IL-6 family receptors, followed by consecutive downmodulation of the α-receptor and later also gp130. These novel insights will help to deepen our understanding on NK cell regulation in the context of malaria and could be of interest for other infections where the investigated cytokines are of importance. In addition, experiments using ex vivo isolated NK cells of semi-immune adults and of P. falciparum-infected individuals should be performed. It would be particularly interesting to analyze expression patterns of IL-6 receptors in malaria patients, since elevated IL-6 production is associated with an increased number of clinical episodes (Robinson and others 2009). Especially in cerebral malaria, IL-6R and other members of the family may be important because of their role in neurological processes. In P. vivax and P. falciparum infection, high serum concentration of soluble IL-6R was associated with a more severe phenotype (Sarthou and others 1997; Wenisch and others 1999). This increases the layer of complexity and points toward the analysis on a systemic level, which could include downstream targets such as the JAK-STAT pathway, and the effect of IFN-α cross-signaling as influence of type I IFN on NK cell gene expression (Grangeiro de Carvalho and others 2011; Sharma and others 2011). In conclusion, this study provides insights into the regulation of IL-6 family members in a frequently used NK cell line and indicates that further research on the role of IL-6 family receptors and NK cells would benefit from system-wide approaches, including primary cells and cell–cell interactions.
Footnotes
Acknowledgments
This work was published as a part of the Ph.D. thesis of E.B. by Universitätsbibliothek Tübingen.
Author Disclosure Statement
No competing financial interests exist.