
Abstract
Transforming growth factor β (TGF-β), interleukin-10 (IL-10), and vascular endothelial growth factor (VEGF) are three of the commonly studied cytokines playing an important role in tumor initiation and progression. Besides their promotional effects on tumor progression, the three cytokines have immunosuppressive effects that facilitate tumor initiation and progression as well. Ganoderma lucidum polysaccharides (Gl-PS) with multiple bioactivities may have the effect on B16F10 melanoma cells to induce stronger antitumor immune response that has been demonstrated. Gl-PS may have the suppressive effects on the production of these three cytokines, which has yet to be demonstrated. In this study, we tested these effects of Gl-PS by incubating Gl-PS with malignant tumor cells such as B16F10 cells, a melanoma cell line, and LA795 cells, a lung carcinoma cell line. RT-qPCR and enzyme-linked immunosorbent assay showed that the production of TGF-β1, IL-10, and VEGF in B16F10 melanoma cells and LA795 lung carcinoma cells was suppressed by Gl-PS at both mRNA and protein levels, suggesting that the suppression on production of TGF-β, IL-10, and VEGF in B16F10 melanoma cells and LA795 lung carcinoma cells by Gl-PS may contribute to the therapy on melanoma and lung carcinoma along with the induction of stronger antitumor immune response.
Introduction
C
G. lucidum (Fr.) Krast (Ganodermataceae) is a well-known medicinal mushroom widely used for centuries in China to promote longevity and improve vigor without significant adverse effects (Chan and others 2008). The Gl-PS is one of the main effective components isolated from G. lucidum (Fr.) Krast (Ganodermataceae). As with many natural products with immunomodulatory and antitumor effects (Selvaduray and others 2010; Geldenhuys and others 2011; Kour and others 2011; Mishra and others 2011; M Ller and others 2011; Murphy and others 2011, 2012; Manca and others 2012), Gl-PS has been extensively studied in recent decades. It has been demonstrated that Gl-PS has various biological activities, including immunomodulatory and antitumor effects (Cao and Lin 2002, 2003; Lin and Zhang 2004; Zhu and Lin 2005; Zhu and others 2007; Li and others 2008; Sun and others 2011). It is widely believed that the antitumor effects of Gl-PS are primarily achieved by boosting host immune function (Lin 2005; Lin and Zhang 2004).
Besides, Gl-PS may also act on tumor cells to improve the antitumor immunity. Our previous studies demonstrated that Gl-PS promoted B16F10 cells to activate lymphocytes (Sun and others 2011b) and induced stronger cytotoxicity in CTLs with granzyme B and porforin by acting on B16F10 cells (Sun and others 2012a), while the enhanced MHC class I and costimulatory molecules on B16F10 cells (Sun and others 2012b) and the antagonism against the suppression on lymphocytes as well as macrophages caused by culture supernatants of B16F10 melanoma cells (Sun and others 2011c, 2013; Lu and others 2013) may partially explain the promoting effects of Gl-PS on B16F10 cells to activate lymphocytes and induce stronger cytotoxicity in CTLs. Considering that the TGF-β, IL-10, and VEGF have immunosuppressive activity, we hypothesize that Gl-PS may also suppress the production of TGF-β, IL-10, and VEGF in the B16F10 cells, which has yet to be demonstrated. The present study was designed to detect the suppressive effects of Gl-PS on the production of TGF-β, IL-10, and VEGF in the B16F10 cells.
Materials and Methods
Preparation of Gl-PS
The preparation of Gl-PS was described previously (Sun and others 2011c). After isolation from a boiling water extract of Gl, ethanol precipitation, dialysis, and Sevag deproteination were processed for Gl-PS preparation. The prepared Gl-PS was a polysaccharide peptide with a molecular weight of 584,900, and the ratio of polysaccharides-to-peptides was 93.51:6.49. The D-rhamnose, D-xylose, D-fructose, D-galactose, D-mannose, and D-glucose were contained in the polysaccharides, with a molar ratio of 0.793:0.964:2.944:0.167:0.389:7.94, and linked together by β-glycosidic linkages. The peptides consisted of 16 kinds of amino acids. The Gl-PS was dissolved in serum-free RPMI-1640 medium (Gibco BRL, Gaithersburg, MD), then filtered through a 0.22-μm filter, and stored at 4°C. Gl-PS was further diluted to the indicated final concentration (0.2, 0.8, 3.2, or 12.8 μg/mL), and fetal bovine serum (FBS) was supplemented to 10% of the final concentration before each assay.
Cell culture
Mouse B16F10 melanoma cells (H-2b) were grown at 37°C in a humidified atmosphere containing 5% CO2 in RPMI 1640 medium supplemented with 10% FBS, penicillin (100 IU/mL), and streptomycin (100 μg/mL). After 4 h of culture for adherence, B16F10 cells in 6-well culture plates (2×105 cells/well at the start) were treated with Gl-PS by replacement of the medium with a medium containing Gl-PS in the indicated concentration and cultured for a further 48 h. RPMI 1640 medium instead of Gl-PS was used as a control. To identify that the effects of Gl-PS are not unique to B16F10 cells but more general findings, another carcinoma cell line, LA795 mouse lung carcinoma cell line, was used to detect the production of the three cytokines with enzyme-linked immunosorbent assay (ELISA). The LA795 cells were placed into 96-well culture plates (1×105 cells/well at the start) and the Gl-PS was supplemented in indicated final concentrations. The LA795 cells were cultured for 48 h and then the culture supernatants were collected for ELISA.
Enzyme-linked immunosorbent assay
ELISA was also performed for its more quantitative potential. It was performed with ELISA kits (NeoBioscience, Shenzhen, China) according to the manufacturer's instructions. In brief, microtiter plates were, respectively, coated with specific antibodies to capture TGF-β1, IL-10, or VEGF in the cell culture supernatants of B16F10 or LA795 cells. A second-layer antibody was then added. Cytokine concentrations were determined with a standard curve derived from known amounts of the relevant cytokine using absorbance readings at 450 nm on a spectrophotometer (Bio-Rad, Hercules, CA). To improve the reliability, the experiments for ELISA, identical to RT-qPCR, were performed three separate times with three paralleled wells in each time, and the means of the three paralleled wells in each separate ELISA were calculated, respectively. The data from the three separate assays were collected together for statistical analysis.
RT-qPCR analysis for determination of gene-specific mRNA expression
Total RNA was extracted from Gl-PS-treated B16F10 cells using the TriZol reagent (Invitrogen, Carlsbad, CA). Isolated RNA (5 μg) was reverse transcribed (RT) into cDNA using M-MLV First strand kit (Invitrogen). Q-PCR (Real-Time PCR) was performed using TransStart Top Green qPCR SuperMix (TransGen Biotech, Beijing, China) on SLAN Real-Time PCR Detection System (Shanghai Hongshi Medical Technology Co., Ltd, Shanghai, China). The PCR primer sequences used were as follows: TGF-β1, forward 5′- CCTGAGTGGCTGTCTTTTGAC -3′ and reverse 5′- GTGGAGTTTGTTATCTTTGCTGTC -3′; IL-10, forward 5′- GGGAAGAGAAACCAGGGAGAT -3′ and reverse 5′- TGCTACAAAGGCAGACAAACAA -3′; VEGF, forward 5′- AGTCTGTGCTCTGGGATTTGAT -3′ and reverse 5′- GCTCTTGATACCTCTTTCGTCTG -3′; β-actin, forward 5′- AAGCCCTGGATGAAGAAACAG -3′ and reverse 5′- TGGGAACCAATCTCGTAGTGTC -3′. The expression of β-actin was used as a control for normalizing mRNA expression results. Reactions were carried out on the Real-Time PCR detection system and were subjected to initial 30-s denaturation at 94°C followed by 45 cycles at 94°C for 5 s and 51°C for 20 s as well as 72°C for 10 s for detection of TGF-β1 mRNA in B16F10 cells or 30-s denaturation at 94°C followed by 45 cycles at 94°C for 5 s and 52°C for 20 s as well as 72°C for 10 s for detection of TGF-β1 mRNA in LA795 cells, and 30-s denaturation at 94°C followed by 45 cycles at 94°C for 5 s and 50°C for 20 s as well as 72°C for 10 s for detection of IL-10 mRNA in both cells, as well as 30-s denaturation at 94°C followed by 45 cycles at 94°C for 5 s and 53°C for 20 s as well as 72°C for 10 s for detection of VEGF mRNA in both cells. When calculating for ΔCt values, geometric means of Ct values of the reference genes were used. To improve the reliability as well, the data from the RT-qPCR assays were collected and calculated identically to ELISA for statistical analysis.
Statistical analysis
The results are expressed as the mean (±SD) of the three-time experiments, and statistical comparison between the experimental groups versus the control was performed using one-way ANOVA followed by the Dunnett t-test. P<0.05 was considered statistically significant.
Results
Suppression of Gl-PS on the production of TGF-β in B16F10 cells
TGF-β plays a crucial role in the progression of cancer as well as in immunosuppression. The RT-qPCR revealed that the level of TGF-β mRNA in B16F10 cells was statistically suppressed by Gl-PS in the concentrations of 0.2, 0.8, 3.2, and 12.8 μg/mL when compared with the control (all P<0.05; Fig. 1A). The ELISA showed that the TGF-β peptide was suppressed statistically by Gl-PS in the concentrations of 0.2, 0.8, 3.2 and 12.8 μg/mL as well when compared with the control (all P<0.05; Fig. 2A).
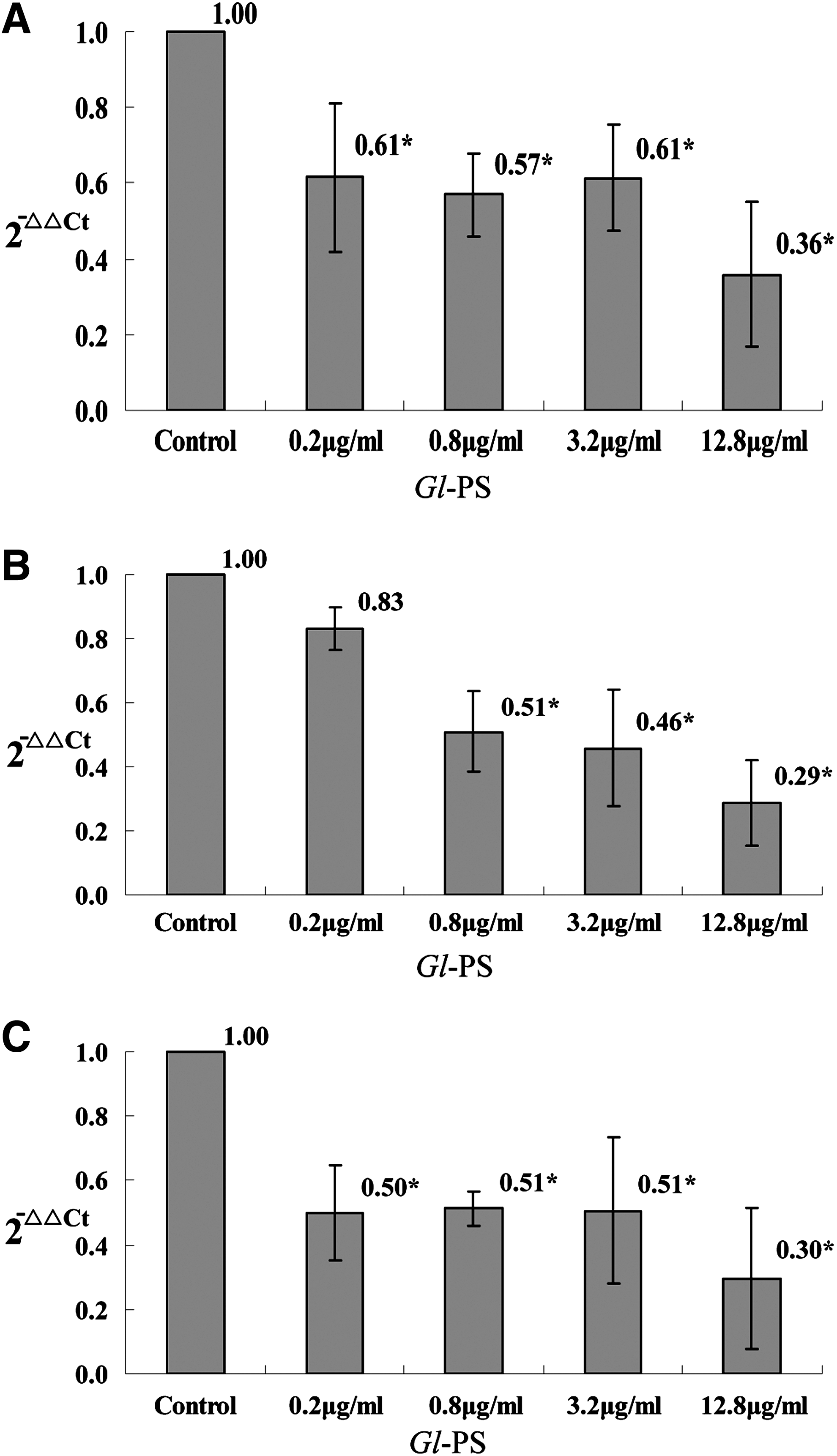
The mRNA of TGF-β1, IL-10, and VEGF in B16F10 cells treated with Gl-PS. B16F10 cells were incubated with Gl-PS for 48 h in three separate tests, and the mRNA extracted from the B16F10 cells with TRIzol reagent was tested with real-time RT-PCR (RT-qPCR) assay. Error bars indicate standard deviation of the mean. Asterisks indicate Dunnett t-test P<0.05 compared with the control after one-way ANOVA.
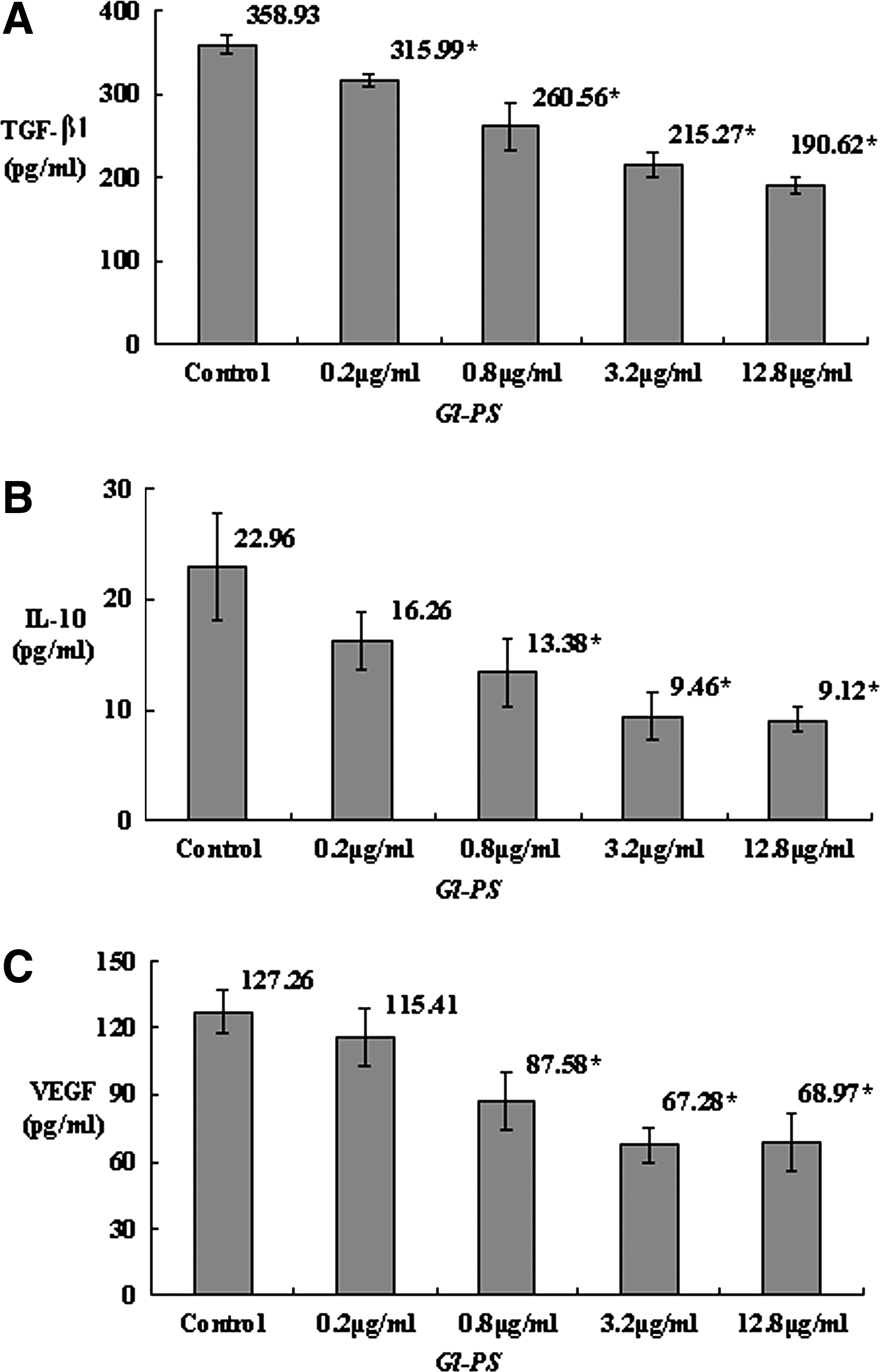
The level of TGF-β1, IL-10, and VEGF in B16F10 cell culture supernatant (B16F10-CS) treated with Gl-PS. B16F10 cells were incubated with Gl-PS for 48 h in three separate tests, and the B16F10-CS was tested with ELISA. Error bars indicate standard deviation of the mean. Asterisks indicate Dunnett t-test P<0.05 compared with the control after one-way ANOVA.
Suppression of Gl-PS on the production of IL-10 in B16F10 cells
IL-10 is a multifunctional anti-inflammatory cytokine that downregulates cell-mediated immune responses and cytotoxic inflammatory responses, and therefore may play a crucial role in the immunosuppression that facilitates cancer progression. It was shown that the level of IL-10 mRNA in B16F10 cells was suppressed by Gl-PS in the concentrations of 0.8, 3.2, and 12.8 μg/mL with statistical significance according to RT-qPCR assay (all P<0.05; Fig. 1B), while the ELISA showed that the IL-10 protein was suppressed statistically by Gl-PS in the concentrations of 0.8, 3.2, and 12.8 μg/mL when compared with the control (P<0.05; Fig. 2B) as well.
Suppression of Gl-PS on the production of VEGF in B16F10 cells
VEGF is a cytokine closely associated with cancer progression as well. The level of VEGF mRNA in B16F10 cells was suppressed by Gl-PS in the concentrations of 0.2, 0.8, 3.2, and 12.8 μg/mL with statistical significance according to RT-qPCR assay (P<0.05; Fig. 1C), while the ELISA showed that the VEGF protein was suppressed statistically by Gl-PS in the concentrations of 0.8, 3.2, and 12.8 μg/mL when compared with the control (P<0.05; Fig. 2C).
Effects of glycogen type II on the production of the cytokines in B16F10 cells
Considering that the effects of Gl-PS could be because of nonspecific actions of polysaccharides in general when applied to cells in culture at these concentrations, we also studied the effects of glycogen type II (Sigma, St. Louis, MO) (in the identical concentration with Gl-PS) on the production of the three cytokines in B16F10 cells with RT-qPCR and ELISA. There was no statistical suppression on the production of the three cytokines by glycogen type II (Figs. 3 and 4), which was different from Gl-PS, for which all of the three cytokines were suppressed, indicating that different polysaccharides had different effects other than nonspecific actions of polysaccharides in general.
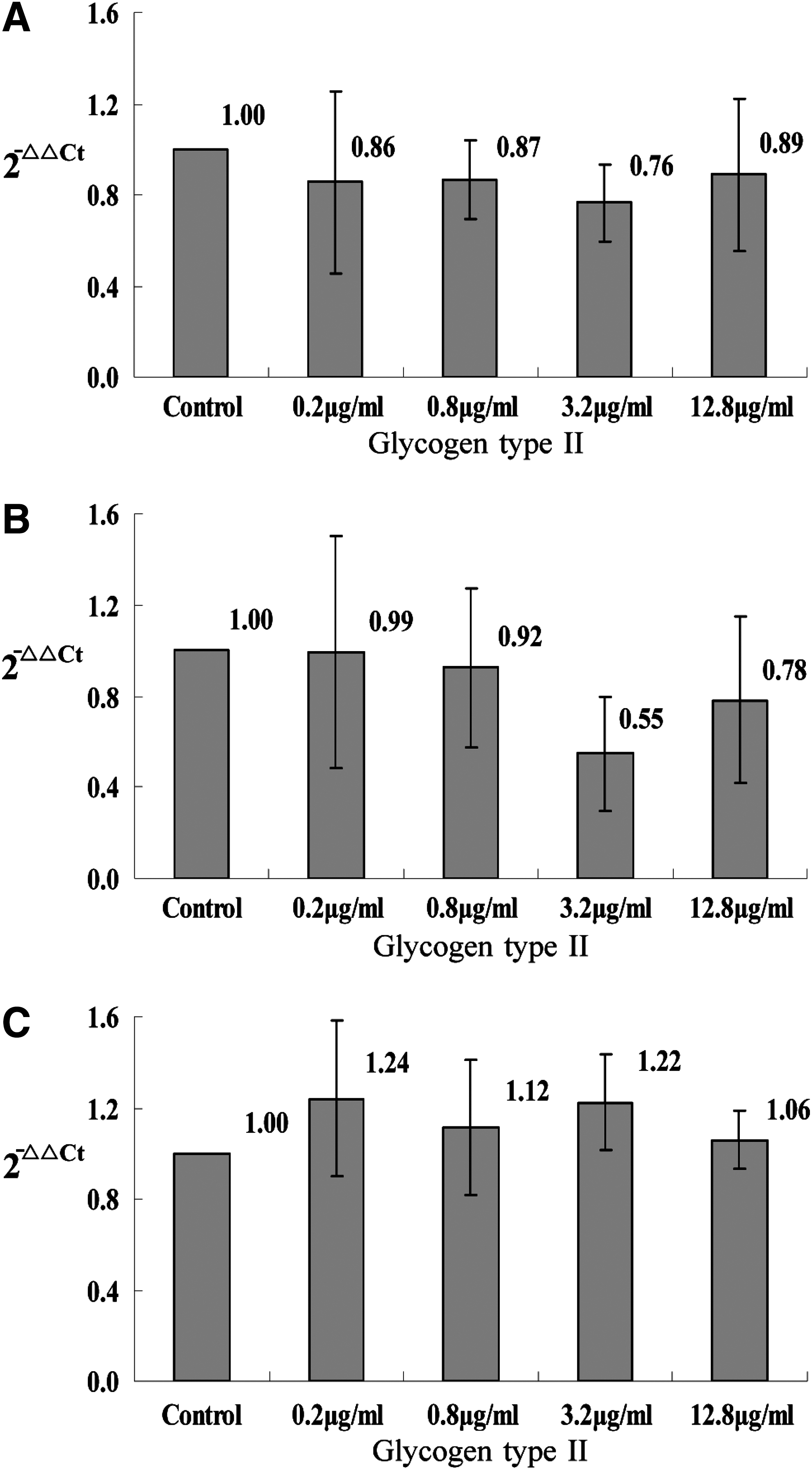
The mRNA of TGF-β1, IL-10, and VEGF in B16F10 cells treated with glycogen type II. B16F10 cells were incubated with glycogen type II for 48 h in three separate tests, and the mRNA extracted from the B16F10 cells with TRIzol reagent was tested with RT-qPCR assay. Error bars indicate standard deviation of the mean. There was no statistical significance according to Dunnett t-test (P<0.05) compared with the control after one-way ANOVA.
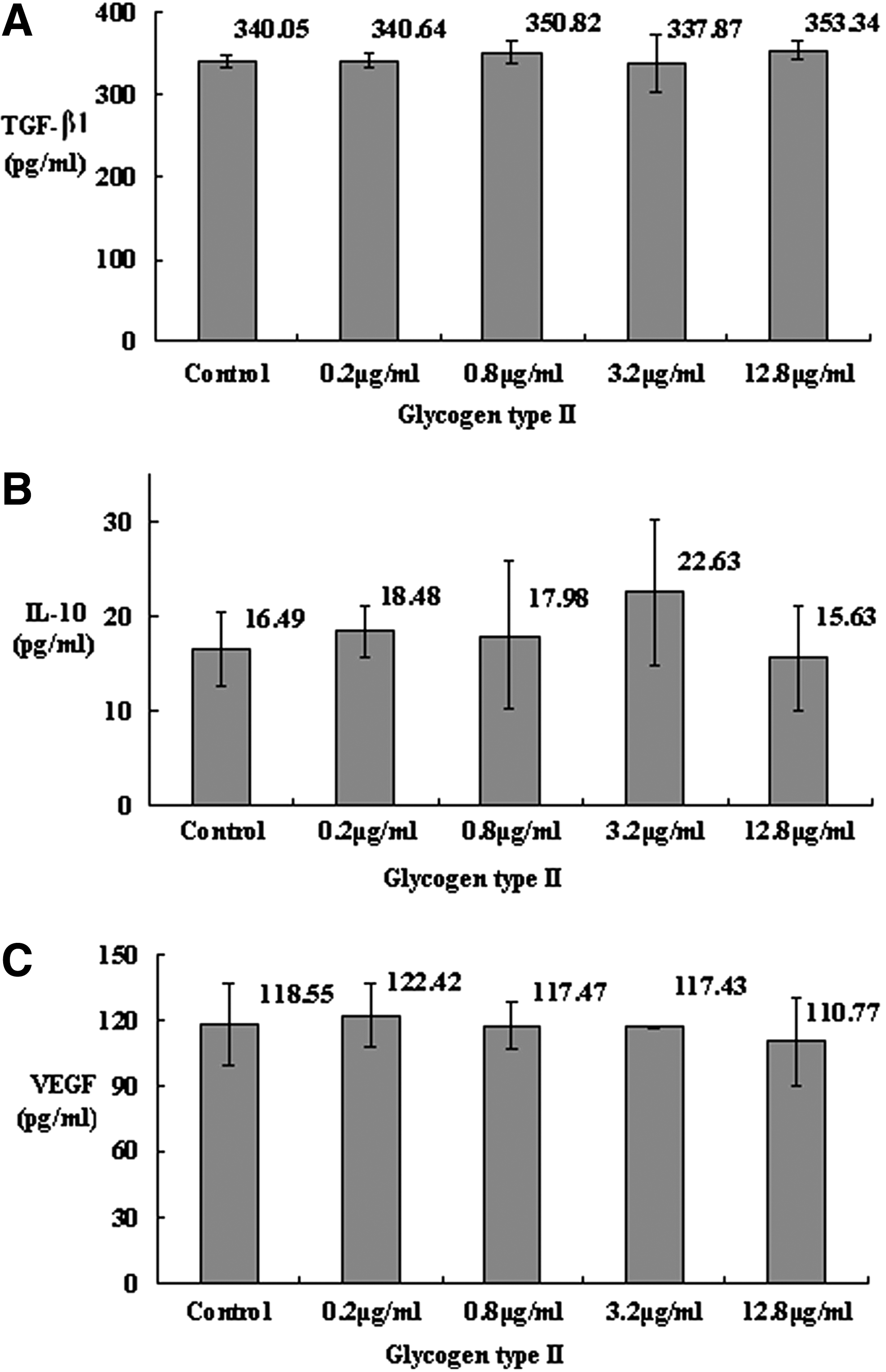
The level of TGF-β1, IL-10, and VEGF in B16F10 cell culture supernatant (B16F10-CS) treated with glycogen type II. B16F10 cells were incubated with glycogen type II for 48 h in three separate tests, and the B16F10-CS was tested with ELISA. Error bars indicate standard deviation of the mean. There was no statistical significance according to Dunnett t-test (P<0.05) compared with the control after one-way ANOVA.
Effects of Gl-PS on the production of the cytokines in LA795 cells
To identify that the effects of Gl-PS on the production of the three cytokines are more general findings instead of unique to B16F10 cells only, another cancer cell line, LA795 mouse lung carcinoma cell line, was used for RT-qPCR and ELISA. The results similar to Gl-PS were shown (Figs. 5 and 6), indicating the more general findings instead of unique to B16F10 cells.
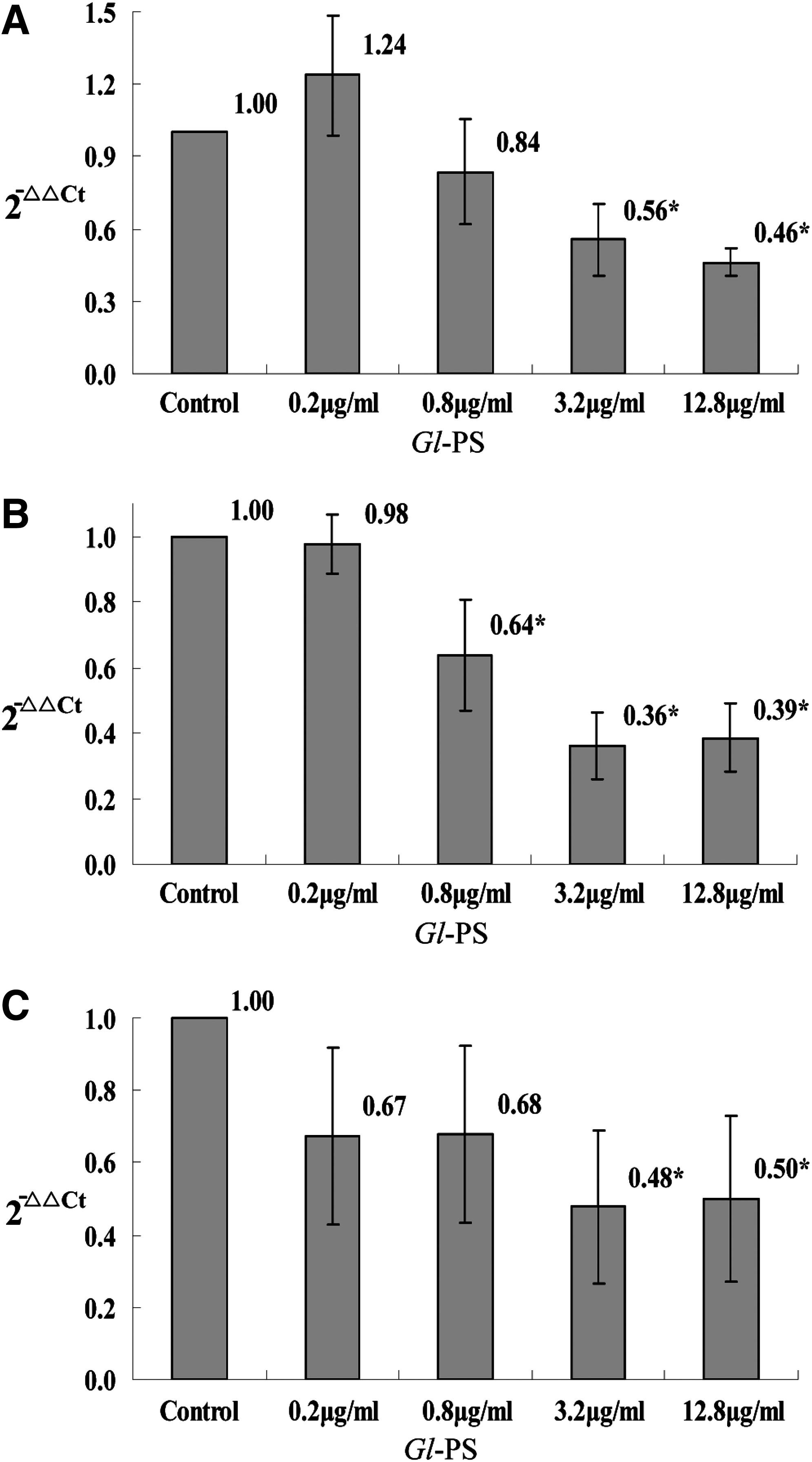
The mRNA of TGF-β1, IL-10, and VEGF in LA795 cells treated with Gl-PS. LA795 cells were incubated with Gl-PS for 48 h in three separate tests, and the mRNA extracted from the LA795 cells with TRIzol reagent was tested with RT-qPCR assay. Error bars indicate standard deviation of the mean. Asterisks indicate Dunnett t-test P<0.05 compared with the control after one-way ANOVA.
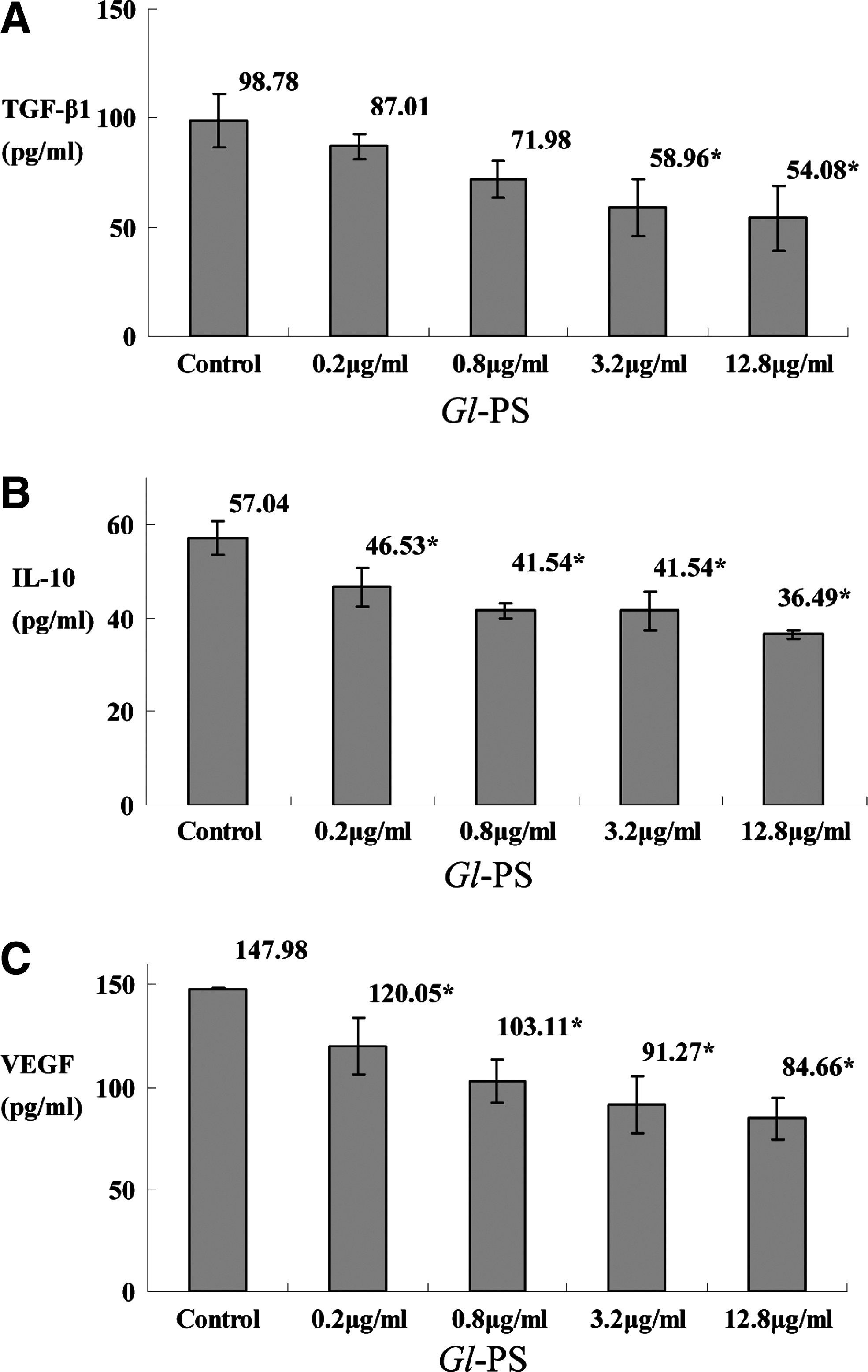
The level of TGF-β1, IL-10, and VEGF in LA795 cell culture supernatant (LA795-CS) treated with Gl-PS. LA795 cells were incubated with Gl-PS for 48 h in three separate tests, and the LA795-CS was tested with ELISA. Error bars indicate standard deviation of the mean. Asterisks indicate Dunnett t-test P<0.05 compared with the control after one-way ANOVA.
Effects of glycogen type II on the production of the cytokines in LA795 cells
To identify that the effects of glycogen type II on the production of the three cytokines in LA795 cells similar to B16F10 cells, in which no suppression on the production of the three cytokines by glycogen type II was shown, glycogen type II was used on LA795 cells for RT-qPCR and ELISA. The results similar to B16F10 cells were shown (data not shown), indicating that besides the B16F10 cells, there was no suppression by glycogen type II on the production of the three cytokines in LA795 cells as well.
Discussion
Although the immune system is capable of mounting immune responses to cancer, it is not always effective in preventing and/or controlling tumor growth. The mechanisms of tumor escaping from immune recognition/destruction are likely to be multifactorial (Ahmad and others 2004). The blockage or inhibition of the host's immune response may partially consist of the evasion mechanisms. The immune suppression occurs with tumor immune tolerance at both a molecular and cellular level gradually and locally, then progresses, and finally spreads to the whole organism (Zou 2005). TGF-β, IL-10, and VEGF are three of the commonly studied cytokines produced by cancer cells and account for part of the immunosuppressive strategies in cancer (Evans and others 2006).
The TGF-β family of cytokines encompasses a number of versatile polypeptide growth factors regulating a multitude of cellular processes in almost every aspect of cellular function, including proliferation, differentiation, apoptosis, migration and adhesion, and inappropriate signaling by these factors, therefore, is closely linked to cancer and other diseases (Zhang 2011). Although the promoter regions of genes encoding three mammalian TGF-β (TGF-β1, 2, 3) show little similarity in sequence, their protein products are functionally very similar (Kaminska and others 2005). TGF-β supports tumor progression (Drabsch ten Dijke 2011) by stimulating the transdifferentiation of epithelial cancer cells into migratory mesenchymal cells (Miyazono 2009), by promoting cell invasion, and dissemination to distant sites (Massagué 2008), enhancing angiogenesis (ten Dijke and others 2008), and mediating immune evasion of tumor cells (Flavell and others 2010). Two distinct effects of TGF-β to ultimately favor tumor progression are inhibition of T-cell proliferation and repression of the cytotoxic gene program in T cells (Thomas and Massagué 2005). TGF-β represses transcription of genes encoding multiple key proteins, such as perforin, granzymes, and cytotoxins, resulting in marked and direct suppression on the cytotoxicity of CTLs (Thomas and Massagué 2005; Trapani 2005). T-cell-specific blockade of TGF-β signaling leads to the generation of tumor-specific CTLs capable of eradicating tumors in mice challenged with EL-4 thymoma or B16-F10 melanoma tumor cells (Gorelik and Flavell 2001). Therefore, suppression on the TGF-β production in tumor cells may inhibit its promotional effects on tumor progression and suppressive effects on immune responses against tumor, facilitating inhibition of tumor progression. In this study, the level of TGF-β1 mRNA as well as the TGF-β1 peptide in B16F10 cells was suppressed by Gl-PS, implying the effects of Gl-PS to control tumor progression in this way.
IL-10 is an important tumor-growth
VEGF is another important cytokine that plays an important role in endothelial cell proliferation and the process of angiogenesis, which are essential for tumor development (Fidler 1991). Besides its important role as pivotal angiogenic factors for tumor vascularization that is important for invasion and metastasis of tumor, VEGF also induces immune suppression as well (reviewed in Evans and others 2006). VEGF inhibits DC maturation (Oyama and others 1998) and function (Laxmanan and others 2005), induces DC apoptosis, and alters DC immunophenotypic profile (Della Porta and others 2005). It interferes with the development of T cells from early hematopoietic progenitor cells (Ohm and others 2003), correlates with reduced levels of the antitumor cytokine IL-12 (Lissoni and others 2001), and is associated with an increase in the production of immature myeloid cells (Gabrilovich and others 1999). In combination with chemotherapy, Bevacizumab, a humanized monoclonal antibody that inhibits VEGF, can now be used as a first-line therapy for the treatment of patients with metastatic colorectal cancer (Ellis 2005) as it improves survival (Mulcahy and Benson 2005). Its antitumor activity is most likely because of its antiangiogenesis effect (Wang and others 2004); however, exposure to anti-VEGF blocking antibodies reverses the inhibitory effect on DC, improving both their function and numbers (Gabrilovich and others 1999; Della Porta and others 2005). In this study, the VEGF mRNA transcription and the VEGF production were suppressed by Gl-PS in certain concentrations, implying its benefit for cancer control as well.
It has not been found what level of Gl-PS in patients ingesting this mushroom may be reached. In this study, the concentrations of Gl-PS in cell culture model were chosen according to some of the previous studies in which the same concentrations were used (Cao and Lin 2002, 2003). In addition, there are studies using the Gl-PS concentrations within or over this study in vitro that showed bioactivity similar to the study in vivo, implying that the concentration used in this study may be achieved in the body by administration in vivo (Zhang and Lin 2004; Chen and others 2011).
Although a study using tissue culture model such as this study is useful and beneficial to improve the scientific knowledge and may encourage relevant research for practice, many additional relevant studies such as the in vivo studies, including clinical trials, and pharmacokinetic studies, in which whether the levels of Gl-PS in patients ingesting these mushroom reach the concentrations that were used to treat these cultured cells will be revealed, are important and valuable, which may be performed in the future.
Footnotes
Acknowledgment
The authors thank Prof. Shuqian Lin of the Fuzhou Institute of Green Valley Bio-Pharm Technology for providing the Gl-PS.
Author Disclosure Statement
The authors declare that they have no conflicts of interest to disclose.