
Abstract
In this study, we determined the effects of glutathione (GSH)-enhancing agents in restoring the levels of GSH in isolated macrophages from individuals with HIV infection thereby resulting in improved control of Mycobacterium tuberculosis. Our results indicate that treatment with N-acetyl cysteine or a liposomal formulation of glutathione (lGSH) resulted in replenishment of reduced also known as free GSH (rGSH), and correlated with a decrease in the intracellular growth of M. tuberculosis. Finally, we observed differences in the amount of the catalytic subunit of glutamine-cysteine ligase (GCLC), glutathione synthase, and glutathione reductase present in macrophages derived from healthy and HIV-infected individuals. These changes correlated with changes in free radicals as well as rGSH levels. Our results indicate that HIV infection leads to increased production of free radicals and decreased production of GCLC resulting in depletion of rGSH and this may lead, in part, to the loss of innate immune function observed in HIV patients. These findings represent a novel mechanism for control of M. tuberculosis infection, and a possible supplement to current HIV treatments.
Introduction
A
The first stage step in M. tuberculosis infection occurs when the alveolar macrophage internalizes the bacilli following inhalation of droplets. Consequently, macrophages in the alveoli are the first cells to become infected by M. tuberculosis (Russell and others 2012). Macrophages being professional phagocytic cells are the first line of defense against microbial invasion (Liu and Modlin 2000; Nathan and Shiloh 2000). The phagocytic pathway allows the macrophages to internalize microbes into compartments where the microbes are brought into contact with reactive oxygen intermediates, antimicrobial peptides, and a generally acidic and hydrolytic environment, which facilitates killing of these internalized pathogens (Russell and others 2012). We have previously reported that GSH is integral in facilitating the control of intracellular growth of M. tuberculosis in both human and murine macrophages (Venketaraman and others 2003, 2005; Dayaram and others 2006; Venketaraman and others 2008). We have also reported that GSH activates the functions of T lymphocytes and NK cells to control M. tuberculosis infection inside human monocytes (Guerra and others 2011, 2012). We have also previously reported that GSH levels are significantly decreased in PBMCs isolated from individuals with HIV infection and this decrease correlated with increased production of proinflammatory cytokines and enhanced growth of M. tuberculosis (Venketaraman and others 2006).
Composed of glutamine, cysteine, and glycine and utilized as an antioxidant, GSH is the main nonprotein thiol responsible for cellular homeostasis and maintenance of the cellular redox balance (Wu and others 2004; Foreman and others 2009). Existing in 2 forms, oxidized (GSSG) and reduced/free form (rGSH), only rGSH exhibits antioxidant activity. During oxidative stress, rGSH is utilized to neutralize reactive oxygen species leading to the formation of GSSG. GSSG is the by-product of the free radical scavenging activity of rGSH and lacks antioxidant function to (Wu and others 2004; Foreman and others 2009). De novo synthesis occurs in a 2-step reaction catalyzed by 2 separate enzymes, glutamine-cysteine ligase (GCL) and glutathione synthase (GSS). The first step in the reaction is catalyzed by GCL, a heterodimer made up of a catalytic subunit (GCLC) that possesses the enzyme's active site and performs the actual amino acid linkage, and a modulating subunit (GCLM) that regulates the activity of GCLC. This first step is rate limiting, with cysteine being the rate-limiting reactant (Wu and others 2004; Foreman and others 2009). In the final step of de novo rGSH synthesis, glycine is linked to the dimer formed in the previous step reaction by GSS. De novo synthesis of rGSH is regulated by negative feedback. Gamma-glutamyltransferase 1 (GGT1) is a membrane-bound enzyme that catalyzes the breakdown of extracellular rGSH into glutamate and cysteinyl-glycine, providing raw materials for the de novo of rGSH. Finally, rGSH can be obtained by the reduction of GSSG via the glutathione reductase (GSR) enzyme. This reaction requires NADPH and forms 2 rGSH molecules from 1 GSSG molecule (Wu and others 2004; Foreman and others 2009).
We recently reported that the levels of intracellular GSH (both total GSH and rGSH) in macrophages from HIV+ individuals are compromised (Morris and others 2012).
We hypothesize that supplementing GSH will result in a restoration of intracellular rGSH and a corresponding restoration of macrophage functions. We tested our hypothesis by performing in vitro studies using human monocyte-derived macrophages isolated from healthy individuals and individuals with HIV infection. We also investigated the mechanisms related to the loss of GSH in macrophages isolated from individuals with HIV infection and compared the findings to macrophages from healthy individuals. Our results indicate that the composition of total GSH in HIV− macrophages favors the rGSH, that is, reduced form of GSH, which has antioxidant activity, while the composition of total GSH in HIV+ macrophages favors the GSSG, that is, oxidized form of GSH, which lacks antioxidant activity. We observed that increasing rGSH in both in HIV+ and HIV− macrophages corresponded with increased control of growth of intracellular M. tuberculosis. We also observed differences in the amount of enzymes involved in the synthesis of GSH in HIV− and HIV+ macrophages. In particular, we found that the levels of GCLC, the catalytic subunit of the key GSH synthesis enzyme are reduced in HIV+ macrophages when compared to HIV− macrophages. The findings suggest that maintenance of intracellular GSH may be compromised with the use of precursor amino acids of GSH as the enzymes used to form GSH in these cells are compromised. In investigating the effect of the decreased function of the GCLC enzyme, we compared the results of GSH restoration in macrophages by treatment with N-acetyl cysteine (NAC), the rate-limiting precursor of GSH versus lGSH, a liposomal form of GSH that has been documented to provide the intact molecule of GSH to the intracellular environment. Our findings show that the lGSH is more efficient by a factor of 1,000 in replenishing the macrophage GSH to a level that was capable of controlling the intracellular replication of M. tuberculosis. These findings describe the mechanism of the loss of GSH in macrophages, discuss the possible mechanisms of this loss, and present a novel mechanism for control of M. tuberculosis infection, and a possible supplement to current HIV treatments. We conclude that restoring the levels of rGSH reverts the loss of innate immune functions observed in macrophages derived from HIV-positive individuals.
Materials and Methods
Statement of ethics
All studies were approved by both the Institutional Review Board and the Institutional bio-safety committee of the Western University of Health Sciences. All study participants were above the legal age of consent at the time of participation and written informed consent was obtained from all volunteers before participation in the study.
Subjects
A total of 26 volunteers (13 healthy subjects and 13 individuals with HIV infection) were recruited for the study. Individuals with HIV infection were recruited from the Foothills AIDS project. Healthy subjects without HIV infection or a history of TB were recruited from the faculty and staff of the Western University. All HIV-infected volunteers had been diagnosed with HIV-1, were taking some form of anti-retroviral treatment (ART), and had CD4+ T-cell counts between 271 and 1,415 cells per mm3. Thirty-five milliliters of blood was drawn once from both healthy volunteers and individuals with HIV infection.
Isolation of monocytes and in vitro culture for differentiation to macrophages
PBMCs were first isolated from the whole blood of healthy and HIV-infected individuals using density gradient centrifugation with ficoll histopaque, a high-density pH neutral polysaccharide solution (Sigma). Plasma samples from healthy and HIV-infected subjects were collected for cytokine measurement. Monocytes were isolated from PBMCs by adding 1×105 cells/well to poly-
Assay of malondialdehyde in macrophage lysates
Malondialdehyde (MDA) was measured in macrophage lysates prepared at 5 days postinfection with H37Rv by colorimetric assay (Cayman Chemical).
Assay of GSH levels in macrophages from healthy and HIV-infected subjects
GSH levels were measured in isolated macrophages from healthy subjects and individuals with HIV infection at 5 days postinfection with H37Rv by spectrophotometry using an assay kit from Arbor Assays. Briefly, macrophages (3×105) were detached from the culture plate by treatment with trypsin (50 μL/105 cells) for 10 min in culture conditions. Detached cells were washed and resuspended in an ice cold 5% 5-sulfo-salicylic acid dehydrate solution (MP Biomedicals). Supernatants collected after centrifugation were analyzed for total and oxidized GSH as per the manufacturer's instructions. rGSH was calculated by subtracting measured oxidized GSH concentrations from the measured total GSH concentrations. All GSH measurements were normalized with total protein levels.
Assay of total protein levels in macrophage supernatants and cell lysates
Proteins in the isolated macrophage supernatants and lysates were measured by the Bradford's method (Bradford 1976) using a Coomassie protein assay reagent (Thermo Scientific).
Preparation of bacterial cells for macrophage infection
All infection studies were performed using the virulent laboratory strain of M. tuberculosis, H37Rv inside the bio-safety level 3 (BSL-3) facility. M. tuberculosis was processed for infection as follows: static cultures of H37Rv at their peak logarithmic phase of growth (between 0.5 and 0.8 at A600) were used for infection of macrophages. The bacterial suspension was washed and resuspended in RPMI (Sigma) containing AB serum (Sigma). Bacterial clumps were disaggregated by vortexing 5 times with 3-mm sterile glass beads. The bacterial suspension was passed through a 5-μm syringe filter (Millipore) to remove any existing clumps. The total number of organisms in the suspension was ascertained by plating. Processed H37Rv was frozen as stocks at −80°C. At the time of infection, frozen stocks of processed H37Rv were thawed and used for macrophage infection.
Infection of macrophages with M. tuberculosis
Monocyte-derived macrophages were infected with processed H37Rv, a virulent laboratory strain of M. tuberculosis, at a multiplicity of infection of 10:1 and incubated for 2 h to allow for phagocytosis. Unphagocytosed mycobacteria were removed by washing the infected macrophages 3 times with warm sterile phosphate-buffered saline (PBS). Infected macrophages were cultured in RPMI+5% AB serum at 37°C+5% CO2. Infected cultures were terminated at 1 h and 5 days postinfection to determine the intracellular survival of H37Rv.
Treatment of macrophages with NAC, lGSH, and buthionine sulfoximine
Infected macrophages were treated with final concentrations of sterile NAC (10 mM), lGSH (5 and 10 μM; Your Energy Systems, LLC), and buthionine sulfoximine (BSO; 500 μM) for 1 h and 5 days.
Termination of infected macrophage cultures and measurement of colony-forming units
Termination of M. tuberculosis-infected cultures was performed by the addition of 200 μL of distilled sterile water to each culture well. The collected cell lysates were diluted in sterile water and plated on the 7H11 medium (Hi Media) enriched with the albumin dextrose complex, to estimate the extent of H37Rv growth in macrophages by counting of colony-forming units.
Immunocytochemical analysis of GSH synthesis enzymes
Macrophages, both infected with M. tuberculosis and uninfected (control category) were incubated for 5 days in culture conditions on sterile glass cover slips. The macrophages were fixed with 3.8% paraformaldehyde in PBS for 30 min. Fixed macrophages were washed twice for 5 min in ice cold PBS, and then permeabilized with 0.25% Triton X-100 in PBS for 10 min. Permeabilized macrophages were washed 3 times for 5 min with PBS. Macrophages were then blocked for 30 min in 1% bovine serum albumin in PBS+0.2% Tween-20 (PBST). Blocked macrophages were then incubated with mouse anti-human GSR IgG and rabbit anti-human GCLC IgG (AbCam) or mouse anti-human GSS and rabbit anti-human GGT1 (AbCam) in PBST, overnight at 4°C with mild shaking. After incubation with primary antibodies, the macrophages were washed 3 times for 5 min with PBS. The macrophages were next incubated for 2 h at room temperature with secondary antibodies. The secondary antibodies were goat anti-rabbit conjugated with the fluorescent dye Alexa Fluor 488, and donkey anti-mouse conjugated with Alexa Fluor 5446 (Invitrogen). Stained macrophages were washed 3 times for 5 min with PBS to remove excess stain. A single drop of mounting media containing 4′,6-diamidino-2-phenylindole (DAPI) was placed on glass slides before inverting the glass cover slips with attached, stained macrophages and placing them on the slide. The cover slips were then sealed to the glass slide using nail polish around the edges of the cover slip. The stained slides were then viewed using an inverted fluorescent microscope. Images were obtained using an integrated digital camera. Images were subsequently analyzed using ImageJ, a free software program available from the National Institutes of Health (
Results
Assay of GSH in macrophage lysates
Consistent with our earlier observations, we found the composition of total GSH in HIV− macrophages favors the rGSH form (Fig. 1A). Macrophages derived from HIV− individuals had 64% of rGSH and 36% GSSG (Fig. 1A). Whereas, total GSH in macrophages derived from HIV+ individuals favors the GSSG form (66% GSSG and 34% rGSH) (Fig. 1B). Our analysis of the intracellular rGSH concentration also revealed a decrease for HIV− macrophages when infected with M. tuberculosis (Fig. 1C). Treatment of both HIV+ and HIV− macrophages with 10 mM NAC resulted in a marked increase in the levels of rGSH (Fig. 1E, F). Treatment of both HIV− and HIV+ macrophages with 5 μM lGSH resulted in increased levels of rGSH (83% and 53%, respectively) (Fig. 1G, H). Treatment of HIV− and HIV+ macrophages with 10 μM lGSH also resulted in a striking increase in the levels of rGSH (80% and 93%, respectively) (Fig. 1I, J). These increases were comparable to those observed with NAC treatment. Importantly, the doses of lGSH given (5 and 10 μM) were 1,000 times lower than the dosage of NAC given (10 mM). This indicates that lGSH is a much more potent method for raising intracellular rGSH concentrations. In addition, lGSH seems to be more effective at replenishing intracellular rGSH in HIV+ macrophages than in HIV− macrophages. Treatment of both HIV+ and HIV− macrophages with BSO resulted in a decrease in the levels of total GSH (Fig. 1K, L). BSO treatment decreased the ratio of rGSH to GSSG in HIV− macrophages from 48% (Fig. 1C) to 37% (Fig. 1K) and in HIV+ macrophages from 34% (Fig. 1D) to 19% (Fig. 1L).

Determination of oxidized and reduced GSH in untreated, Mycobacterium tuberculosis-infected+NAC-treated (10 mM), M. tuberculosis-infected+lGSH-treated, and M. tuberculosis-infected+BSO-treated (500 μM) macrophages derived from healthy subjects and individuals with HIV infection at 5 days postinfection with H37Rv. Levels of oxidized and reduced GSH are represented as percentages of the total GSH for both HIV+ and HIV− macrophages.
Assay of MDA in macrophage lysates
Infection of both HIV+ and HIV− macrophages with M. tuberculosis resulted in a small increase in the levels of MDA (Fig. 2). Treatment of both HIV+ and HIV− macrophages with NAC resulted in decreases in MDA concentrations, indicating a decrease in free radicals. Treatment of macrophages from both categories with lGSH also produced decreases in MDA levels; also, lGSH (5 and 10 μM) was able to produce results similar to NAC at concentrations 1,000-fold lower than those used in NAC (10 mM) treatment (Fig. 2). Finally, treatment with BSO resulted in 10-fold elevations in the levels of MDA observed in macrophages from both categories.
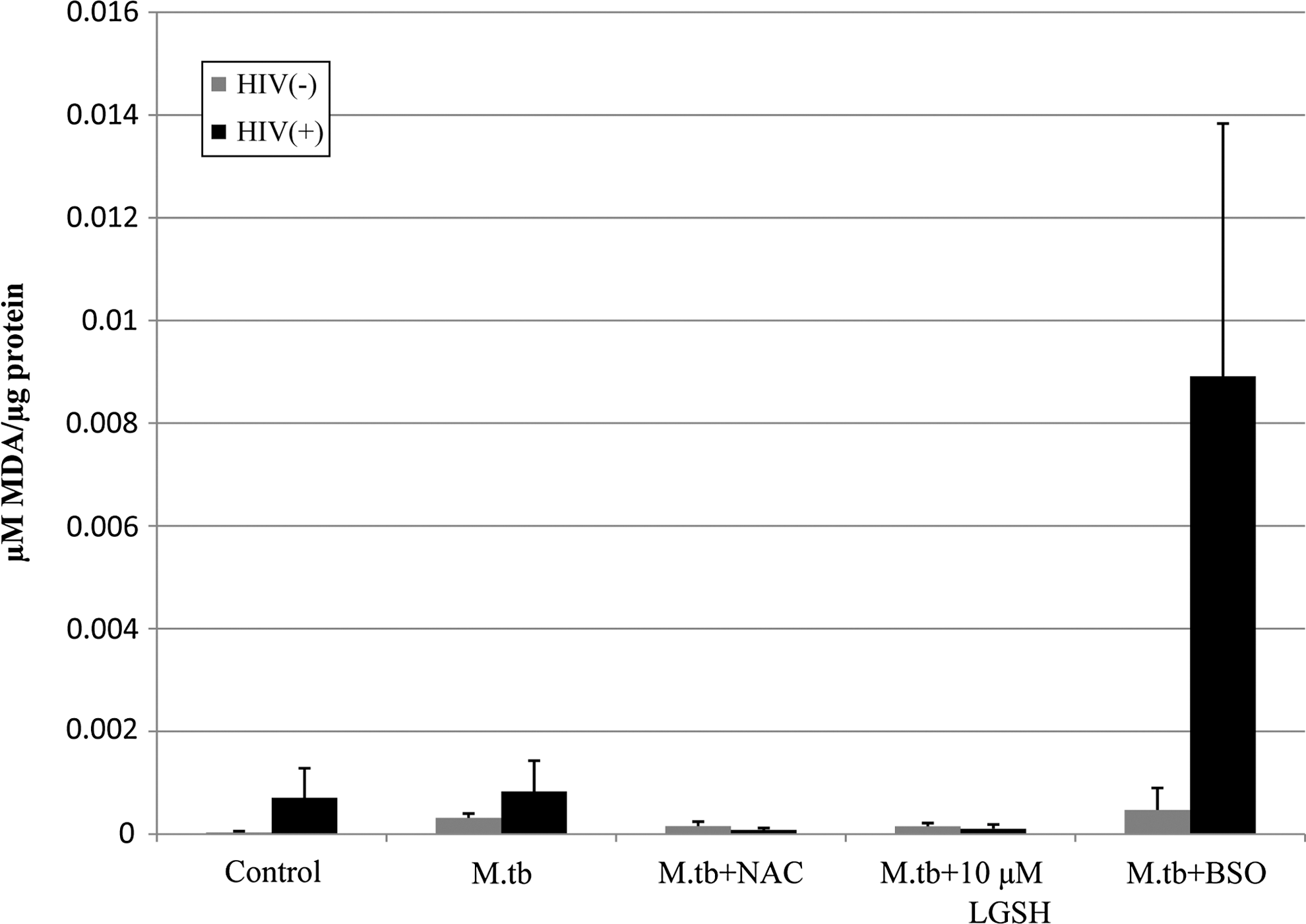
Assay of MDA in macrophage lysates from HIV-infected and healthy subjects prepared at 5 days postinfection with H37Rv. Free radical levels in macrophage lysates from healthy subjects and individuals with HIV infection were determined by measuring the levels of MDA using a colorimetric assay kit from Cayman. These elevated MDA concentrations are indicative of elevated free radical concentrations. Treatment with either NAC or lGSH reduced the amount of MDA observed in both HIV+ and HIV− macrophages. High-standard error values observed in HIV-infected subjects are likely due to the varying stages of HIV infection and antiretroviral treatment in the HIV-infected population. Results are from n=4 HIV+ and n=5 HIV− individuals. MDA, malondialdehyde.
Determination of the intracellular survival of M. tuberculosis in isolated monocyte-derived macrophages
Treatment of macrophages from both study groups with NAC resulted in the control of intracellular M. tuberculosis growth (Fig. 3A, B). The addition of lGSH to the culture media of both HIV+ and HIV− macrophages also resulted in a marked decrease in intracellular M. tuberculosis growth. lGSH was able to achieve this control at concentrations 1,000 times lower than NAC (Fig. 3A, B). Finally, treatment with BSO resulted in an increase in the intracellular growth of M. tuberculosis in both healthy and HIV-infected macrophages (Fig. 3A, B). We did not observe any loss in the number of macrophages either due to the effects of M. tuberculosis infection or due to the cytotoxic effects of NAC, lGSH, and BSO treatment. Furthermore, we measured the levels of proteins in the macrophage lysates, and we observed similar levels of proteins across various treatment categories in each study group confirming that there is no significant loss in the number of macrophages either due to infection or treatment.
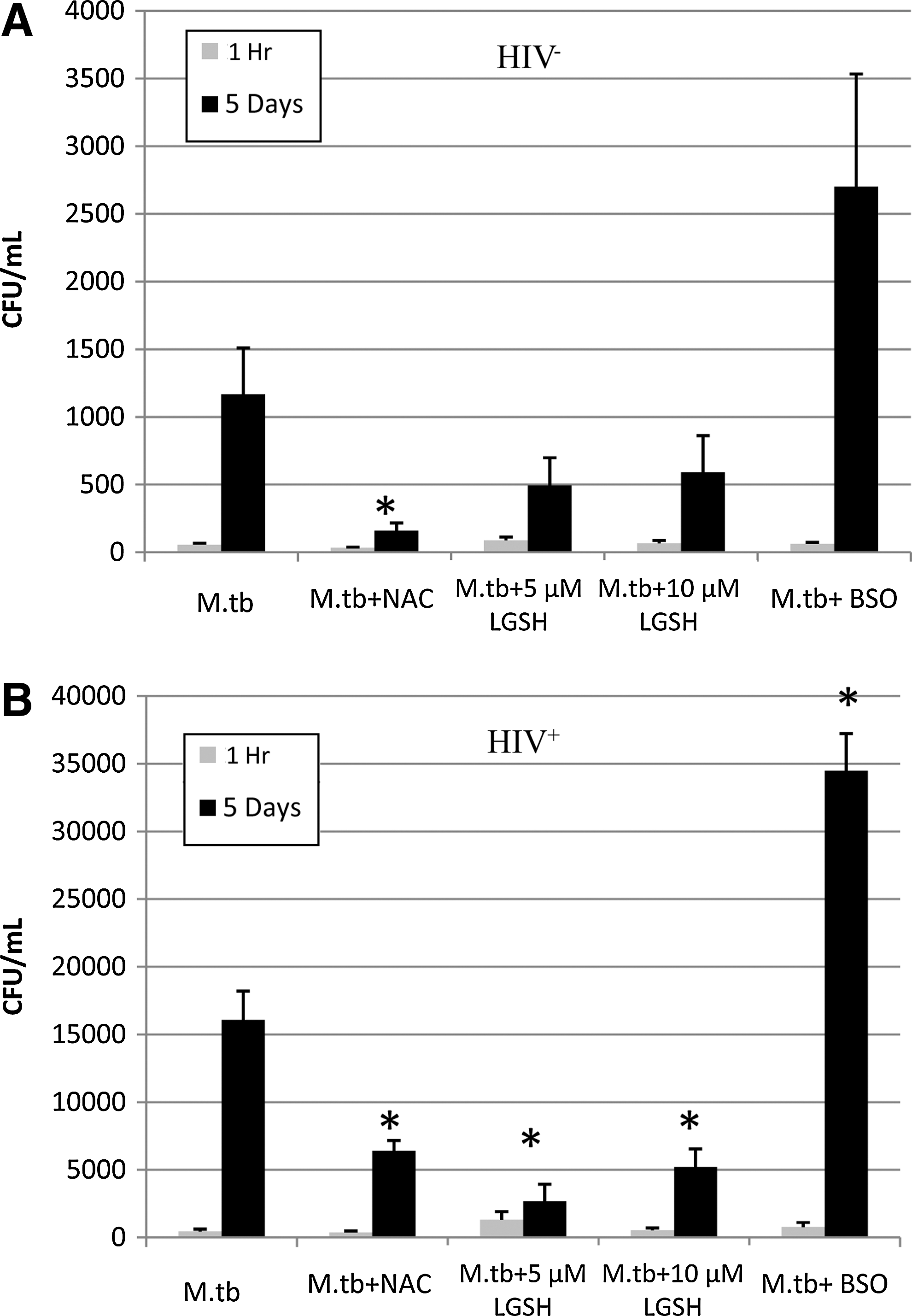
Intracellular survival assay of M. tuberculosis in macrophages from HIV+ and HIV− subjects at 1 h, and 5 days postinfection. Human monocyte-derived macrophages were infected with M. tuberculosis at a multiplicity of infection of 10:1. Infected macrophages were treated with NAC, lGSH, and BSO and terminated at 1 h and 5 days postinfection to determine the intracellular survival of M. tuberculosis inside macrophages. Macrophage lysates were plated on 7H11 medium enriched with albumin dextrose complex to estimate the growth or killing of M. tuberculosis. Each experiment was performed in triplicate. *P≤0.05 comparing each treatment category to the untreated category at the 5-day time point.
Immunocytochemical analysis of GSH synthesis enzymes
Immunocytochemical analysis to compare the expressions of GCLC in HIV+ and HIV− macrophages revealed decreased GCLC in HIV+ macrophages in comparison to HIV− macrophages for all treatment categories, with the exception of those treated with BSO (Figs. 4 and 5). There was no statistically significant change in GCLC with M. tuberculosis infection in both HIV+ and HIV− macrophages. Treatment with either NAC or BSO resulted in significant increases in GCLC for both HIV+ and HIV− macrophages (Fig. 5).

Fluorescent microscopy of HIV− and HIV+ macrophages from the control category (20× magnification). Secondary antibodies labeled with Alexa Fluor 488 (green) and Alexa Fluor 5446 (red) directed against primary antibodies for GCLC and GSR.

Immunocytochemical analysis of GCLC in macrophages isolated from healthy and HIV-infected subjects at 5 days postinfection with H37Rv. Results are for n=3 individuals for both groups. (*P≤0.05 comparing macrophages from healthy and HIV-infected subjects for each treatment category. ++ P≤0.05 comparing M. tuberculosis-infected macrophages treated with NAC or BSO to untreated M. tuberculosis-infected macrophages for both HIV+ and HIV− macrophages.) HIV+ macrophages display less GCLC than HIV− macrophages across all categories with the exception of BSO treatment. Treatment with NAC and BSO induces elevated production of GCLC in both HIV+ and HIV− macrophages.
Quantifying GSR protein levels, we found that in all categories HIV+ macrophages contain more GSR than HIV− macrophages (Figs. 4 and 6). Infection with M. tuberculosis resulted in an increase in GSR in both HIV+ and HIV− macrophages. Treatment with NAC resulted in a significant decrease in GSR for both HIV+ and HIV− macrophages that were infected with M. tuberculosis (Fig. 6). Finally, our analysis showed a large increase of GSR in both HIV+ and HIV− macrophages that were infected with M. tuberculosis and treated with BSO (Fig. 6).
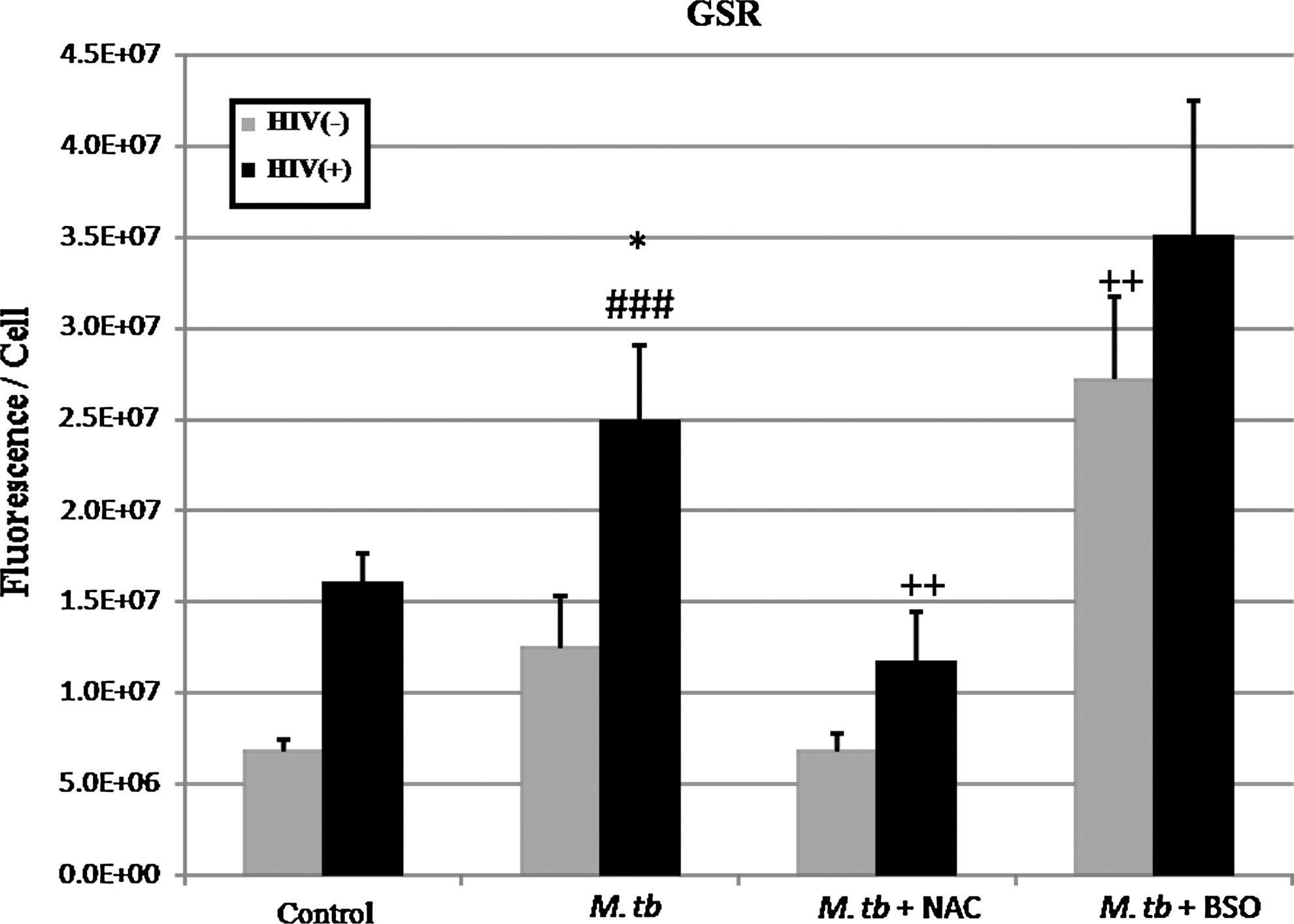
Immunocytochemical analysis of GSR in macrophages isolated from healthy and HIV-infected subjects at 5 days postinfection with H37Rv. Results are for n=3 individuals for both groups. (*P≤0.05 comparing macrophages from healthy and HIV-infected subjects for each treatment category. ++ P≤0.05 comparing M. tuberculosis-infected macrophages treated with NAC or BSO to untreated M. tuberculosis-infected macrophages for both HIV+ and HIV− macrophages. ### P≤0.05 comparing M. tuberculosis-infected macrophages without treatment to macrophages not infected with M. tuberculosis.) GSR is elevated in HIV+ macrophages across all treatment categories. Infection with M. tuberculosis correlates with increased GSR expression. Macrophages treated with NAC correlate with decreased GSR. Macrophages treated with BSO show the maximum GSR.
Upon analysis of GSS proteins in both HIV+ and HIV− macrophages, we found that HIV+ macrophages contained more GSS than HIV− macrophages across all categories. Also, BSO-treated macrophages produced the maximum GSS observed in macrophages from both HIV+ and HIV− individuals (Figs. 7 and 8).
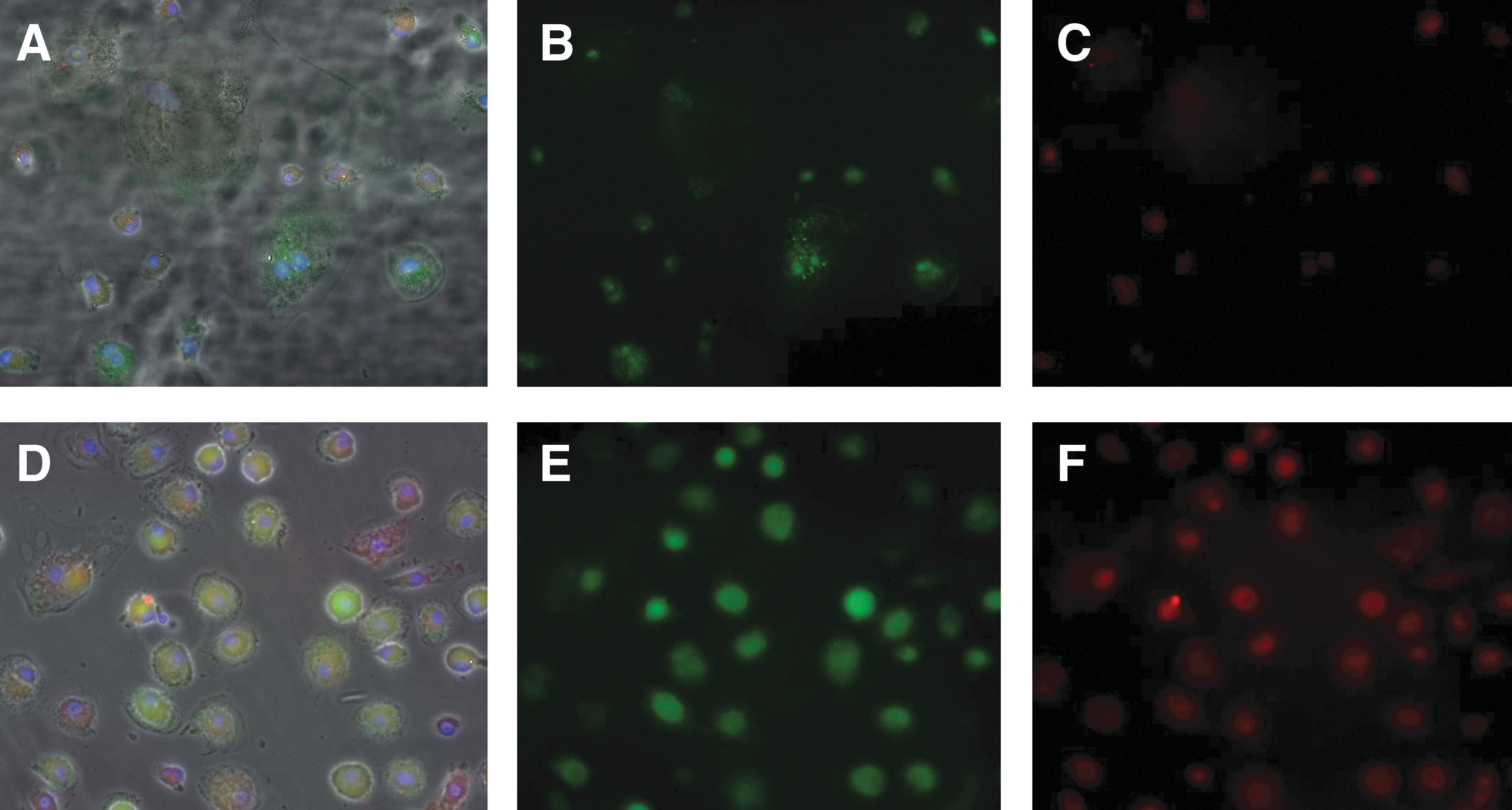
Fluorescent microscopy of HIV− and HIV+ macrophages from the control category (20× magnification). Secondary antibodies labeled with Alexa Fluor 488 (green) and Alexa Fluor 5446 (red) directed against primary antibodies for GSS and GGT.
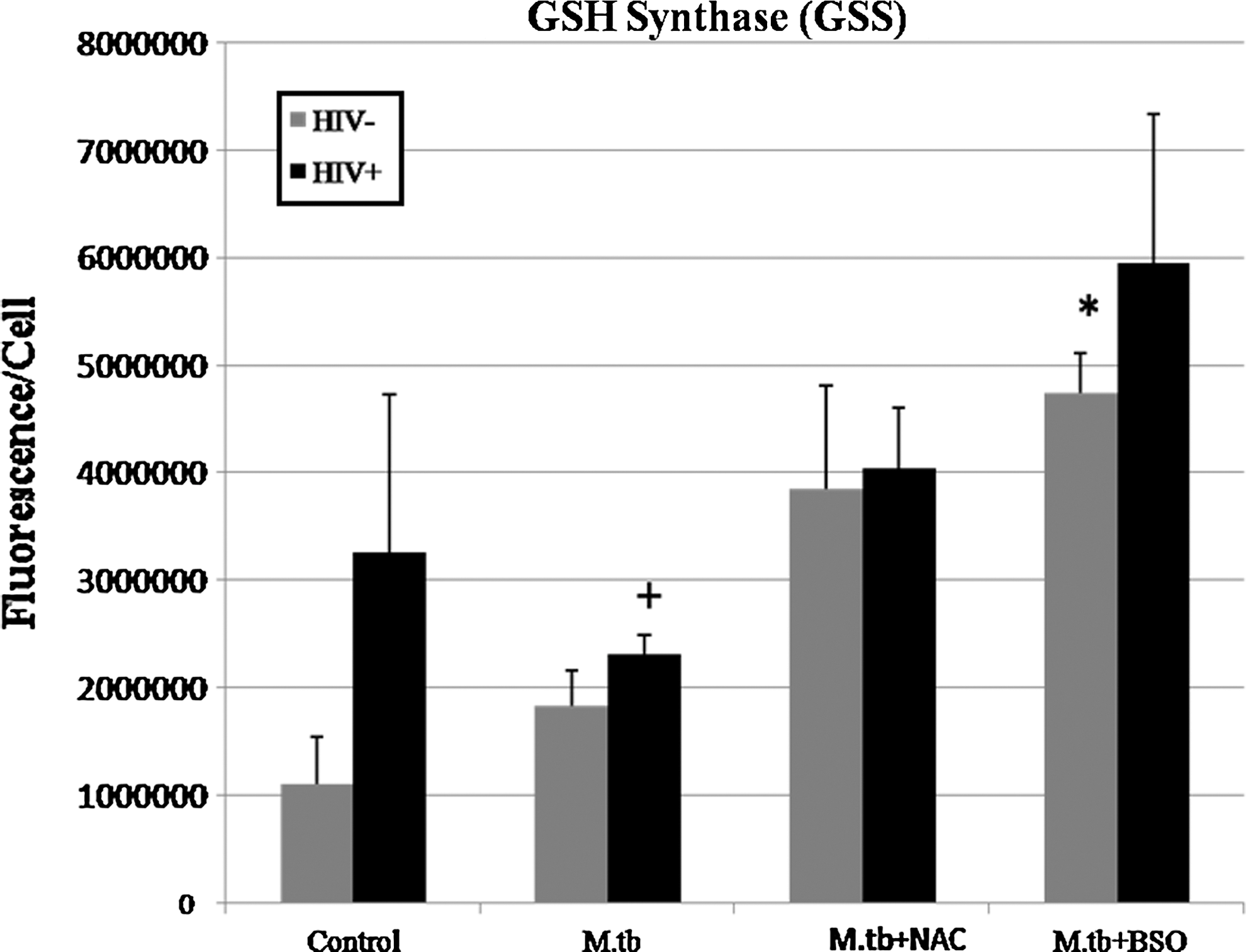
Immunocytochemical analysis of GSS in macrophages isolated from healthy and HIV-infected subjects. Results are for n=3 for HIV− and n=2 for HIV+. (*P≤0.05 comparing treated macrophages from healthy and HIV-infected subjects to their respective control categories. + P≤0.05 comparing HIV+ macrophages to HIV− macrophages of the same treatment category.) GSS is elevated in HIV+ macrophages across all treatment categories. Macrophages treated with BSO show the maximum GSS.
Results from the experiments using GGT were inconclusive due to high variation in samples from both HIV− and HIV+ macrophages; however, GGT seemed to be in greater quantities in HIV+ macrophages for all categories (Figs. 7 –9).
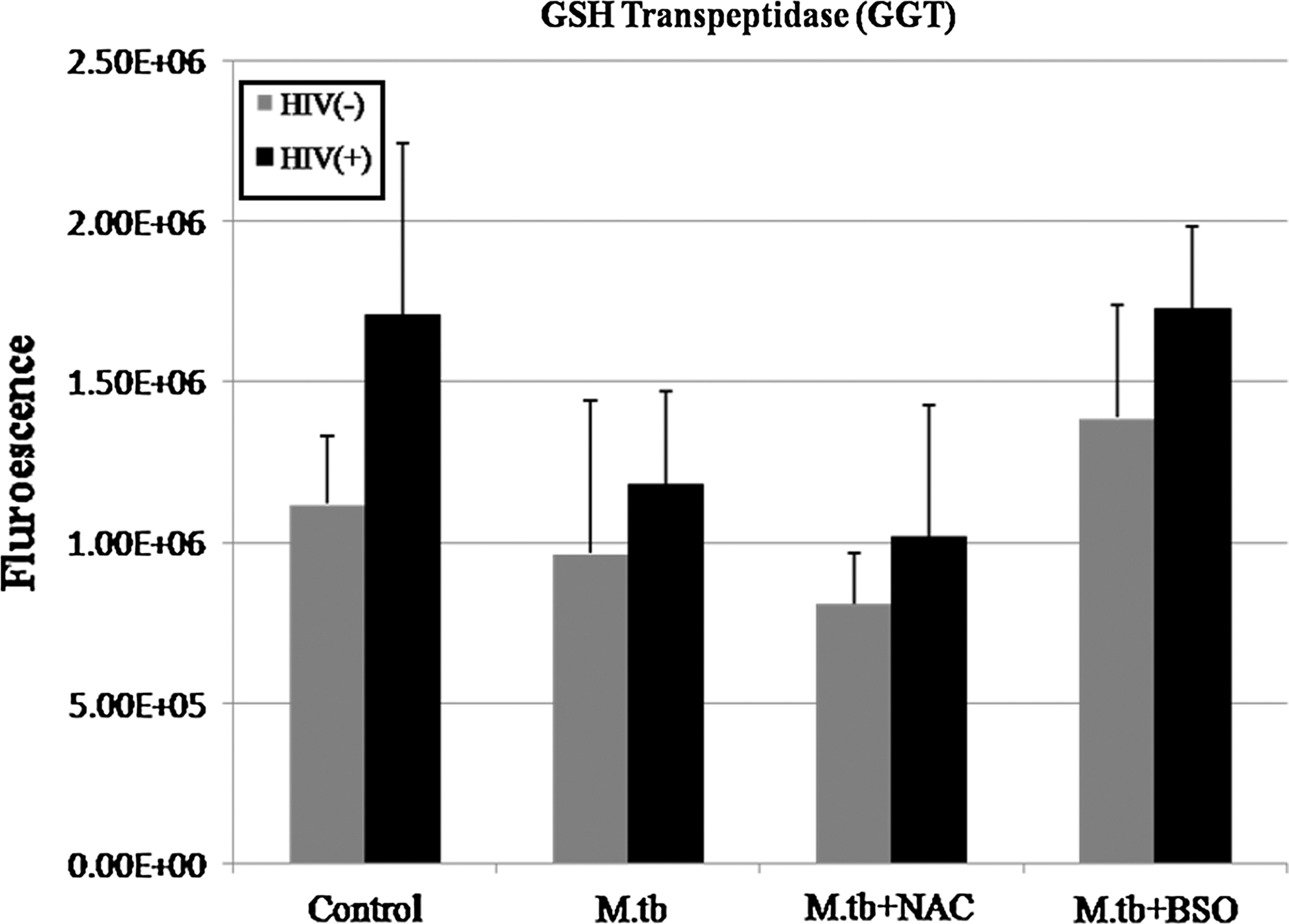
Immunocytochemical analysis of GGT in macrophages isolated from healthy and HIV-infected subjects. Results are for n=3 for HIV− and n=2 for HIV+. GGT is elevated in HIV+ macrophages across all treatment categories.
Discussion
Our assays of intracellular GSH in both HIV+ and HIV− macrophages indicate that treatment with the GSH prodrug NAC increases the amount of available rGSH. NAC appears to be more effective at increasing intracellular rGSH in HIV− macrophages than in HIV+ macrophages (Fig. 1). These results are consistent with observations made by Bakin and others (2005) and Franklin and others (2003) who report that elevated TGF-β can inhibit the production and enhance degradation of GCLC, the catalytic subunit of the enzyme responsible for the rate-limiting step in de novo synthesis of rGSH. In a previous study, we observed elevated levels of TGF-β in both the plasma and macrophage culture supernatants of HIV+ macrophages (Morris and others 2012). This elevated TGF-β will compromise the amount of GCLC present inside the cell; consequently, supplementing the raw materials for de novo synthesis in HIV+ individuals, who are overexpressing TGF-β, will not result in the same increased production of rGSH that is observed in individuals who are not overexpressing TGF-β. In addition, this phenomenon may explain why lGSH at lower concentrations than NAC is more effective at raising the concentration of reduced GSH in HIV+ macrophages than in HIV− macrophages. Supplementing with an lGSH formulation provides complete rGSH molecules to cells, circumventing the enzymatic pathway responsible for rGSH production, without the requirement for the cell to construct the tripeptide (Zeevalk and others 2010). This may also explain why treatment with lGSH seems to raise the ratio of rGSH to GSSG at much lower concentrations than NAC, as cells treated with NAC will have to produce new molecules of rGSH utilizing their own enzymatic machinery. Our assays of MDA that are indicators of lipid peroxidation and free radical formation (Yagi 1998), demonstrate a correlation between levels of intracellular rGSH and free radical formation. Raising the levels of intracellular rGSH decreased free radical formation. Decreasing intracellular rGSH resulted in increased formation of free radicals. This data also correlate with our assay of intracellular M. tuberculosis growth. It has been demonstrated that HIV+macrophages that are deficient in rGSH, have a reduced ability to control intracellular M. tuberculosis infection (Morris and others 2012). We again demonstrate this phenomenon that occurs independent of T cells (Fig. 3), the depletion of which is the major cause of immune-compromised status in HIV patients. This impaired function may be due to rGSH deficiency. In addition, increased levels of the rGSH provided by NAC or lGSH treatment resulted in control of intracellular M. tuberculosis growth. Furthermore, treatment of both HIV+ and HIV− macrophages with BSO, resulted in a large decrease in the availability of intracellular rGSH, and a corresponding decrease in the ability of the BSO-treated macrophages to control intracellular M. tuberculosis infection. Combining the data from all 3 of these experiments supports our theory of GSH, oxidative stress, immune function, and intracellular M. tuberculosis growth.
Expression of GCL and GSR is known to be controlled by nuclear factor E2-related factor 2 (Nrf2) (Hur and Gray 2011). In response to increased oxidative stress, this transcription factor facilitates expression of GCL and GSR as well as several other antioxidant-related genes by binding to specific promoter regions of the genome known as antioxidant response elements (Hur and Gray 2011). Our data for both GCLC and GSR protein levels appear to follow this model of expression closely (Figs. 5 and 6). For both HIV+ and HIV− macrophages, we have observed a decrease in rGSH (Fig. 1), corresponding with an increase in MDA (Fig. 3), in addition to an increase in GCLC and GSR proteins (Figs. 5 and 6) with M. tuberculosis infection in conjunction with BSO treatment. The HIV-1 transactivator protein (TAT) has been shown to increase expression of Nrf2 at both the mRNA and protein levels, while at the same time inducing production of free radicals (Zhang and others 2009) (Fig. 10). Additionally, we observed an increase in intracellular rGSH, corresponding with a decrease in MDA, as well as a decrease in the GSR protein in both HIV+/M. tuberculosis + and HIV−/M. tuberculosis + macrophages that were treated with NAC. Furthermore, chronic infection with HIV appears to lead to a chronic state of inflammation and elevated free radical formation, which may promote elevated expression of GSR in HIV+ macrophages, as GSR is elevated in HIV+ macrophages across all treatment categories when compared to HIV− (Figs. 4 –6). This compensatory mechanism does not, however, appear to be sufficient to replenish intracellular rGSH in HIV+ macrophages, Our studies add value to the existing information on the GCLC expression. Oxidative stress is known to induce the expression of Nrf2 genes, which in turn, will upregulate the expression of GCLC. Production of GCLC appears to be further complicated by the suppressive effect of TGF-β overproduction previously mentioned. In all treatment categories, GCLC protein levels are lower in HIV+macrophages than HIV− macrophages. Our results also indicate that the presence of cysteine (in the form of NAC) can trigger the expression of GCLC to promote the synthesis of GSH.
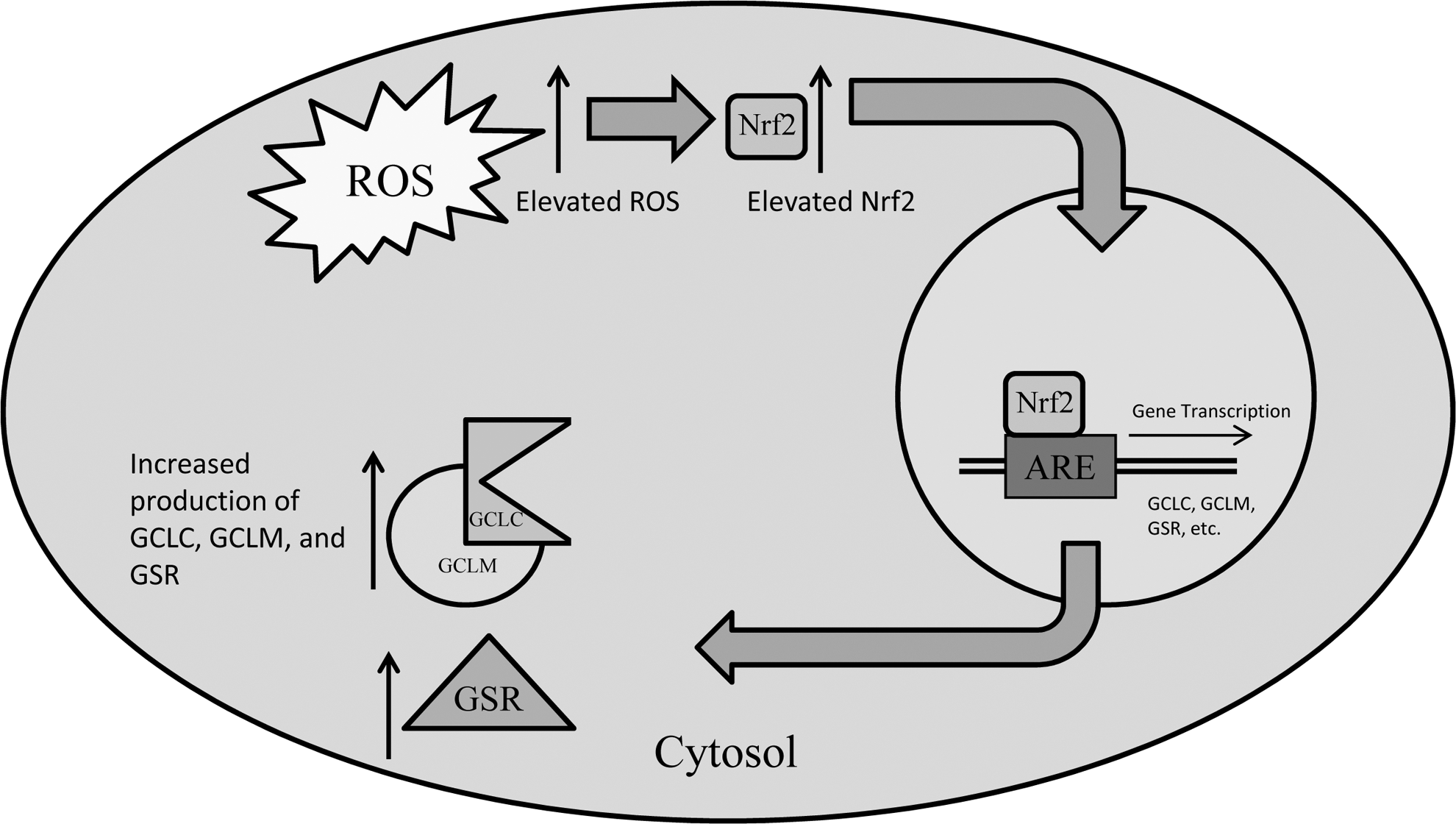
A model describing the effects of elevated ROS on Nrf2 activity and subsequent downstream transcription of genes modulated by Nrf2 activity. Elevated ROS induces enhanced production and activity of the Nrf2 transcription factor. Nrf2 travels to the nucleus and binds to the antioxidant response element, inducing gene transcription of GSH synthesis enzymes (GCLC, GCLM, and GSR), leading to enhanced production of these enzymes. HIV-1 TAT can induce elevation of Nrf2 independent of ROS. Nrf2, nuclear factor E2-related factor 2.
Interestingly, when comparing GCLC protein levels and intracellular rGSH levels with BSO treatment, we observed a large increase in the relative level of GCLC present in both HIV+ and HIV− macrophages; however, there was no corresponding increase in intracellular rGSH as one might expect knowing that GCLC is the catalytic unit of the enzyme responsible for de novo synthesis of reduced GSH. This phenomenon is explained by the mechanism of action for BSO. BSO binds irreversibly to GCLC, inhibiting its activity (Griffith 1982). Therefore, increasing GCLC in the presence of BSO will not result in an increase in rGSH production.
Taken together, the data demonstrate a pattern of chronic inflammation brought on by HIV infection that depletes rGSH and impairs the intracellular killing of M. tuberculosis in macrophages (Fig. 3). By supplementing rGSH, we were able to mitigate the production of reactive oxygen intermediates and improve the ability of macrophages to kill M. tuberculosis intracellularly. Supplementation of rGSH also corresponded with increased production of GCLC and decreased production of GSR, additional indicators that conditions of oxidative stress are decreased through the administration of GSH supplements. This data make a compelling argument for the efficacy of GSH supplements in addition to the ART regimen prescribed to HIV+ patients in improving immune function.
While our study uses an in vitro model of oxidative stress, GSH production, and immune function, these results must be thought of in the context of the human body where myriad other factors may affect these parameters. Additionally, our study utilized clinical samples from HIV patients who were currently taking antiretroviral drugs. These subjects varied in their CD4+ T-cell counts and viral load, indicating they were in different states of disease progression and control. Even with all these complicating factors, our data indicate that supplementing GSH in combination with ART may provide beneficial effects to the host immune system over ART alone. As lGSH provides complete rGSH molecules, bypassing the cellular machinery for GSH production (Zeevalk and others 2010), lGSH may prove to be more effective in supplementing rGSH in HIV patients due to the effects of TGF-β production associated with HIV infection and the observed effect at concentrations lower than NAC.
Several avenues of research have been opened by the results of this study. Of particular interest is the mechanism by which rGSH improves the ability of macrophages to control infection by intracellular pathogens like M. tuberculosis. Is recognition of pathogens improved by increased production of toll-like and/or pattern recognition receptors? Is there improvement in phagosome/lysosome fusion? Further studies may provide insights into the relationships between the macrophage's redox state and immune function.
Footnotes
Acknowledgments
This work is supported by the Potts Memorial Foundation, start-up funds from the Western University of Health Sciences, and in part by Your Energy Systems. As a study sponsor, Your Energy Systems, LLC played no role in the study design, data acquisition, analysis, or writing of the report for publication. We thank Ms. Trina Rich and all other staff at the Western University medical center for their help with drawing blood from the volunteers. We thank the volunteers for their participation in this study.
Author Disclosure Statement
No competing financial interests exist.