
Abstract
The mechanisms underlying the complex and multistage wound-healing process are not yet completely understood. One of the most important and intriguing questions remaining is the effect of the interactions between wounds and the microflora that are present in wounds. In this report, we describe the first study of the effect of treating murine skin wounds with topical bacterial lipopolysaccharide (LPS), the main exogenous ligand of Toll-like receptor 4. Our findings demonstrate that LPS treatment strongly affects the wound-healing process by accelerating the resolution of inflammation, increasing macrophage infiltration, enhancing collagen synthesis, and altering the secretion of a number of mediators that are involved in the skin regeneration process. Topical LPS treatment upregulated the secretion of proinflammatory cytokines [interleukin (IL)-6, IL-1β, and leukemia inhibitory factor (LIF)] and CC-chemokines (CCL2/MCP-1, CCL7/MCP-3, CCL3/MIP-1α, and CCL5/RANTES), but not CXC-chemokines (CXCL2/MIP-2 and CXCL9/MIG). The secretion of growth factors (vascular endothelial growth factor, transforming growth factor-β1 (TGF-β1), and fibroblast growth factor 2) at the wound site was also upregulated. Taken together, these results suggest that the topical application of LPS at the wound surface affects the inflammatory process and promotes the wound healing of injured skin.
Introduction
It was recently shown that the topical application of pathogen-associated molecular patterns (PAMPs) isolated from bacteria or viruses can also enhance inflammation and stimulate cutaneous wound healing. It was reported that the topical administration of MALP-2 (Deiters and others 2004), poly (I:C) (Lin and others 2012), or CpG oligonucleotides (Sato and others 2010) increases the influx of macrophages, improves the neovascularization of the wound bed, and stimulates the wound repair process. These effects were associated with the activation of Toll-like receptor 2 (TLR2), TLR3, and TLR9, respectively.
The TLR family of pattern-recognition receptors is a part of the innate immune system and may be an important modulator of inflammation during the wound-healing process, as these receptors initiate different innate and adaptive immune reactions via the activation of several transcription factors, including NF-κB, IRF3, IRF7, and AP-1. TLRs are expressed by many different cell types that are involved in the wound-healing process, such as keratinocytes, monocytes, macrophages, dendritic cells, endothelial cells of the microvasculature, fibroblasts, adipocytes, and T- and B-lymphocytes (Miller and Modlin 2007). The activation of TLRs by their ligands leads to the initiation of several signaling cascades and subsequently results in the expression of the proinflammatory cytokines, chemokines, adhesion molecules, antimicrobial peptides, and proteolytic enzymes that take part in a complex cross-talk during the wound-healing process, especially during the inflammatory stage.
Bacterial lipopolysaccharide (LPS) is a component of the outer membrane of gram-negative bacteria and is a ligand of TLR4. As of yet, the effects induced by systemic administration of LPS have been thoroughly characterized (Andreasen and others 2008). However, the effects of topical application of bacterial LPS on the inflammation process and on skin repair have not yet been studied. To evaluate how LPS affects these processes, we used Na–carboxymethylcellulose-based gels that contained various concentrations of LPS to treat murine incision wounds. The goals of our research were to determine the influence of topical LPS application on the inflammation process and the wound-healing process as a whole and to identify any changes in the secretion levels of a variety of mediators that are involved in the wound-healing process.
Materials and Methods
Animals and reagents
Inbred BALB/c male mice were used in this study. All mice were between 5 and 6 weeks of age and were purchased from the Stolbovaya nursery of the Russian Academy of Medical Sciences. The mice were fed a completely pelleted laboratory chow and had access to food and tap water ad libitum. All of the experimental procedures were in line with the local ethics committee guidelines.
LPS from Salmonella typhi was obtained from the Medgamal, a biotechnological branch of the NF Gamaleya Research Institute for Epidemiology and Microbiology. The specific activity of the LPS preparation was ∼5×106 EU/mg according to the Limulus Amebocyte Lysate test results. The effect of the LPS on the TLR4- and TLR2-mediated activation of NF-κB was tested according to a previously published method (MacGregor and others 1991). HEK-hTLR4/CD14-MD2 and HEK-hTLR2/CD14 cells that stably coexpress human TLR4 and the coreceptor molecules MD-2 and CD14 or TLR2 and CD14, respectively, were used in this study. All of the cell lines also stably expressed an NF-κB-inducible β-galactosidase reporter gene. This study determined that isolated and purified LPS, such as the commercially available phenol-purified LPS preparation from Escherichia coli 0111:B4 (L2630; Sigma-Aldrich, St. Louis, MO), had a TLR4-specific activity and had a significantly reduced TLR2-specific activity in comparison with the reference LPS.
An ELISA kit for transforming growth fector-β1 (TGF-β1) was purchased from R&D Systems (Minneapolis, MN), and ELISA kits for interleukin (IL)-1β and fibroblast growth factor 2 (FGF2) were purchased from Abcam (Cambridge, United Kingdom). Multiplex kits for the determination of the levels of cytokines, chemokines, and growth factors were purchased from Bio-Rad (Hercules, CA) and Bender MedSystems GmbH (Vienna, Austria). The Bradford protein assay kit was purchased from Silex (Moscow, Russia). The rat anti-mouse monoclonal F4/80 (BM4007), rat anti-mouse monoclonal Ly6G (NB600-1387), and biotinylated rabbit anti-rat IgG (R1371B) antibodies were purchased from Acris Novus Biologicals (Littleton, CO). Other reagents used included medium-viscosity sodium carboxymethylcellulose, glycerol (both from Sigma-Aldrich), Zoletil® (Virbac, Carros, France), Rometar (xylazine; Bioveta, Ivanovice na Hane, Czech Republic), complete protease inhibitor cocktail tablets (Roche Diagnostics, Mannheim, Germany), and tissue protein extraction reagent (T-PER
The LPS gel for topical application was formulated using the sodium salt of carboxymethylcellulose (4% w/w), glycerol (10% w/w), and endotoxin-free water obtained from a Milli-Q® Advantage A10® system (Millipore, Molsheim, France). Appropriate amounts of LPS solution were added to the gel to obtain final LPS concentrations of 0.2, 2, and 10 mg/g (mg LPS/g of gel). The placebo was formulated in the same way, but without the addition of LPS.
Wounding procedure and topical treatment protocol
The mice were anesthetized with an intramuscular injection of a mixture of xylazine (7.5 mg/kg) and Zoletil (45 mg/kg). The backs of the mice were shaved with an Oster Mark II animal clipper (Sunbeam-Oster, Fort Lauderdale, FL), and after disinfection with 70% ethanol, one 4-cm midline dorsal incision was made on the back of each mouse through the epidermis, dermis, and subcutaneous tissue layers while leaving the fascia intact. The skin incision was immediately closed with 2 silk stitches to form 3 wounds of equal size per animal (Fig. 1a). The animals were individually caged and randomly sorted into groups. The sutures were removed at the end of the 5th day after wounding. The treatment consisted of a topical application of 150 μL of the gel on the wound area once a day. The gel was first applied 24 h after wounding. At a specified time point after the first gel application, 5 mice from each group were sacrificed by CO2 inhalation. The cranial and caudal wound segments from each animal were used for biomechanical analysis, and the central wound segments were used for histological or immunohistochemical analysis or for cytokine, chemokine, and growth factor assays.
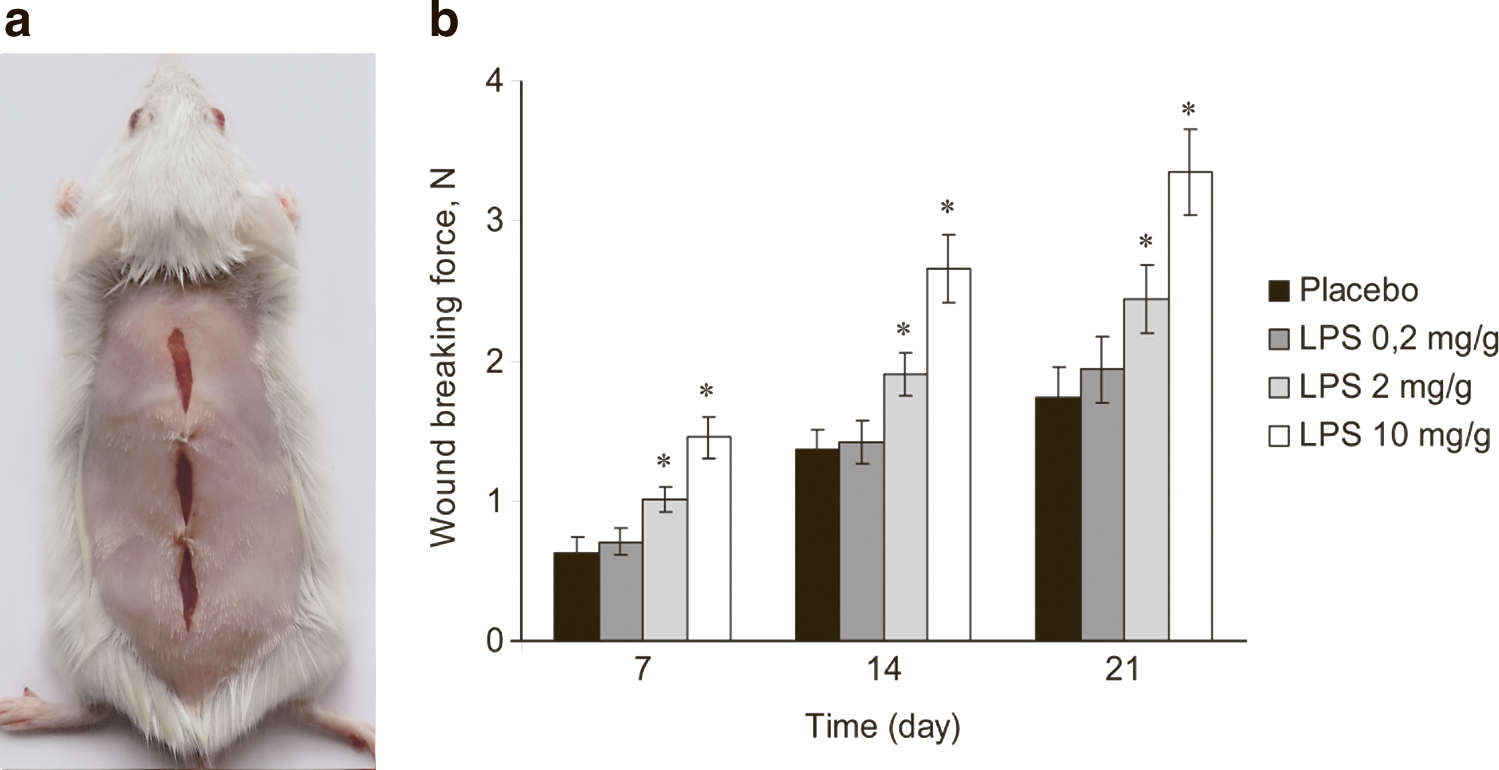
Lipopolysaccharide (LPS) treatment increases the wound-breaking force in a dose-dependent manner.
Biomechanical analysis
The biomechanical analysis was performed on days 7, 14, and 21 after the first gel application according to a previously published method (Muehlberger and others 2005). Briefly, the skin strips were harvested for the biomechanical analysis and were immediately placed in phosphate-buffered saline. Each strip was centered on a segment of the healing scar and mounted horizontally between 2 clamps of a tensiometer (Instron 5848 Microtester; Instron, Canton, MA) with a 50-N load cell. The distance between the 2 clamps was 2.0 mm. The skin was then stretched by moving 1 clamp at a constant rate of 1 cm/min until breaking. The breaking strength was defined as the point of maximal force in Newton (N) before wound separation occurred. The breaking strengths of samples from mice of the different groups were subjected to statistical analysis.
Histological evaluation
Strips of wounded skin harvested from the animals on days 1, 3, 7, 14, and 21 after the first gel application were fixed in 10% buffered formalin and were processed for histological analysis. Tissue blocks were embedded in paraffin, and 4-μm sections from the mid-portions of the wounds were stained with hematoxylin–eosin (H&E) or Masson's trichrome staining. The assessments focused on dermal alterations, including the inflammatory response indicated by the presence of inflammatory cell infiltrates, and the level of collagen deposition and remodeling. All of the slides (3 slides for each staining method per wound) were evaluated by 2 certified pathologists in a blinded manner.
Immunohistochemical analysis
The strips of wounded skin were harvested from the mice on days 1, 3, and 7 after the first gel application, fixed in 10% buffered formalin, and embedded in paraffin. The 4-μm sections from the mid-portions of the wounds were immunostained for the macrophage marker F4/80 (rat anti-mouse monoclonal IgG 2A, 1:500) and for the neutrophil marker Ly-6G (rat anti-mouse monoclonal, 1:50). The secondary antibody was biotinylated rabbit anti-rat IgG (1:2,500). The slides were developed using streptavidin–horseradish peroxidase, followed by diaminobenzidine, after which they were counterstained with Mayer's hematoxylin. The slides were then evaluated under high power (×400) by 2 independent pathologists blinded to the experimental groups. The numbers of F4/80-positive or Ly-6G-positive cells were counted in 5 consecutive high-power fields from the wound edge. No specific staining was detected in slides stained with the secondary antibody alone.
Cytokine, chemokine, and growth factor assays
Skin samples containing the central wound segments were excised with an 8-mm sterile biopsy punch (Dermo-Punch®, Milan, Italy) at the specified time points after the first gel treatment (4, 10, and 24 h, and 3, 7, and 10 days). These samples were immediately placed into an ice-cold T-PER extraction buffer containing a complete protease inhibitor. Approximately 0.75 mL of the buffer was used per 35 mg of skin sample. The homogenized samples were prepared using a FastPrep 24 device (MP Biomedicals, Solon, OH). The homogenates were centrifuged at 12,000 rpm for 12 min at 4°C. The total protein in the supernatant was measured using the Bradford method. Commercial ELISA kits were used to determine the levels of TGF-β1, IL-1β, and FGF2 according to the manufacturer's instructions. The levels of CCL2/MCP-1, CCL7/MCP-3, CCL3/MIP-1α, CCL5/RANTES, and IL-6 were determined using the FlowCytomix platform (Bender MedSystems GmbH). To assess the levels of CXCL2/MIP-2, CXCL9/MIG, leukemia inhibitory factor (LIF), and vascular endothelial growth factor (VEGF), a Bio-plex cytokine analysis kit was used in conjunction with the Bio-plex system array (Bio-Rad) according to the manufacturer's instructions.
Statistical analysis
All of the experiments were repeated at least twice. All of the values represent the mean±SD. The statistical significance of differences among the means was analyzed by one-way ANOVA using the Statistica® 7 statistical software package (StatSoft, Inc., Tulsa, OK). Differences were considered significant if P was <0.05.
Results
LPS treatment increases wound-breaking force in a dose-dependent manner
To quantify the influence of the bacterial LPS on wound healing, we first determined the wound-breaking force 7, 14, and 21 days after the first LPS gel application. The mechanical strength of healing wounds and scars is an integrative parameter that is subject to the interplay of a variety of humoral factors and cellular elements that are involved in the wound-healing process. We found that the topical application of LPS increased this parameter in a dose-dependent manner. The mean breaking forces in the groups treated with gels containing 2 or 10 mg/g LPS were significantly higher than that of the placebo-treated group at all time points. However, no statistically significant increase was observed for the group treated with gel containing 0.2 mg/g LPS (Fig. 1b). A gel containing 10 mg/g LPS was used for further experiments.
Alterations in leukocyte infiltration and inflammation after LPS wound treatment
To evaluate leukocyte accumulation at the wound site (wound histopathology is presented in Fig. 2a), wound sections were stained with monoclonal antibodies specific for macrophages (F4/80) or neutrophils (Ly6G) (Fig. 2b). Our results showed that topical LPS wound treatment induced early macrophage infiltration. One day after the first LPS gel application, each high-power field contained twice as many macrophages as the placebo-treated group. On days 3 and 7, the macrophage influx was similar for both groups, and no statistically significant differences were detected (Fig. 2c). Neutrophils appeared early and peaked on day 1 for both groups. However, the neutrophils began to disappear from the LPS-treated wounds on day 3 and were negligible by day 7. In contrast, the placebo-treated wounds showed more-intensive neutrophil infiltration on days 3 and 7, and these changes were statistically significant (Fig. 2c).

Changes in leukocyte infiltration after LPS wound treatment.
By the end of the first week, histological analysis of H&E-stained wound sections showed increased inflammatory cell infiltration in the placebo-treated wounds when compared to the LPS-treated wounds, indicating the persistence of the inflammatory process (Fig. 3). By day 14, no differences in inflammation were detected between the groups.
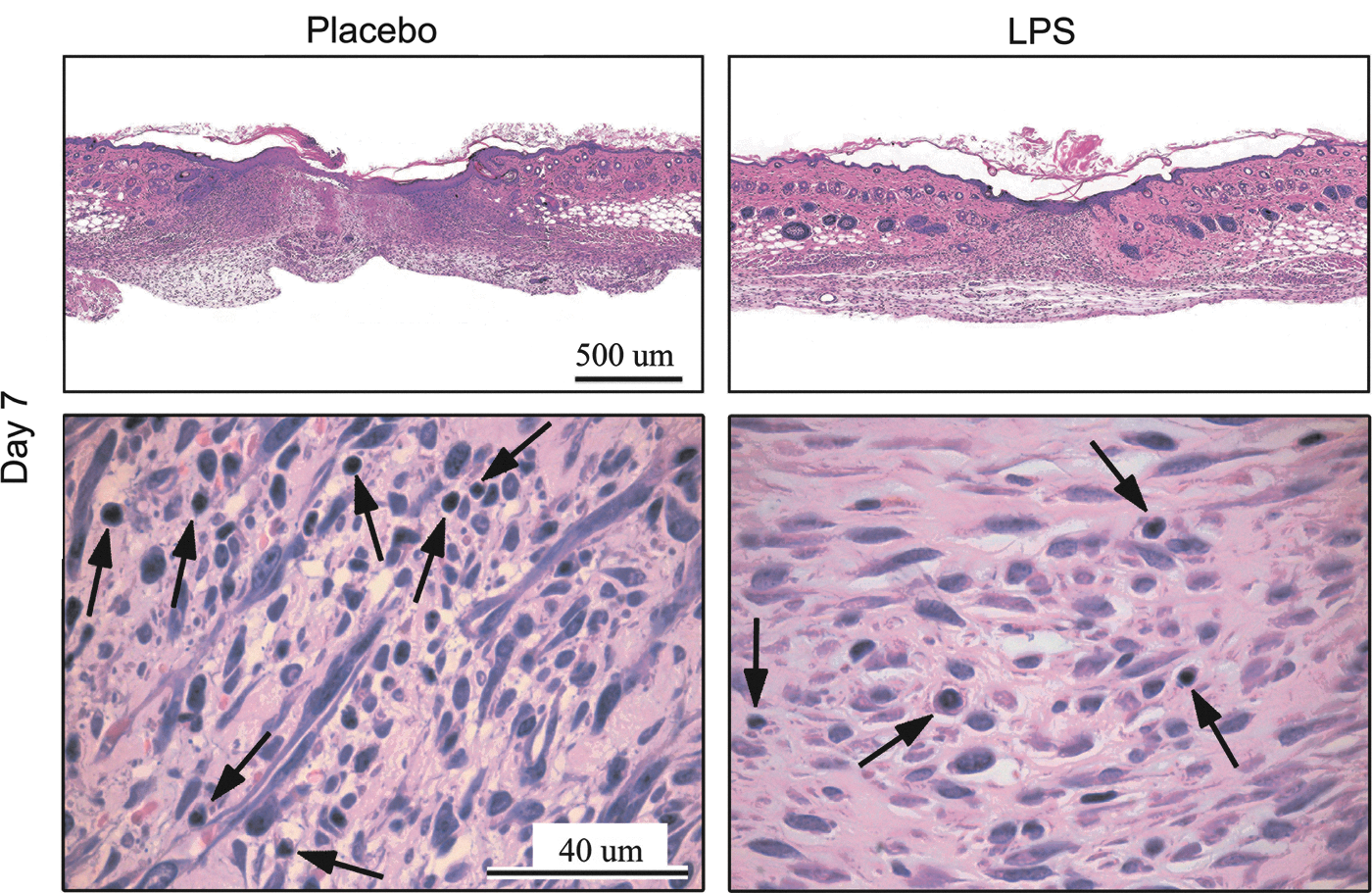
Changes in the wound inflammation process after LPS treatment. Histological hematoxylin–eosin staining of wound sections. The arrows indicate inflammatory cells. Representative results from 1 of 2 independent experiments with 5 animals in each group are shown. The presented time point (day 7) is after the first gel application.
LPS treatment improves collagen synthesis
As wound treatment with LPS increased the wound-breaking force, we hypothesized that LPS stimulates collagen synthesis. The presence and deposition of collagen were evaluated using histology sections treated with Masson's trichrome stain. No differences were detected between the groups on day 7. An increase in the intensity of collagen fiber staining was observed on day 14 in the LPS-treated group. On day 21, the fibers in the wounds from LPS-treated mice were more mature. In contrast, the wounds from placebo-treated mice contained immature collagen fibers (Fig. 4). No excessive scarring was observed after the topical LPS therapy.

Histological Masson's trichrome staining of wound sections. The arrows indicate collagen fibers. Representative results from 1 of 2 independent experiments with 5 animals in each group are shown. Time points after the first gel application are presented. Boxed areas in the upper panels are enlarged in the lower panels.
LPS treatment selectively induces the secretion of chemokines and proinflammatory cytokines and enhances the secretion of growth factors
The observed changes in leukocyte accumulation implied that there may also be changes in chemokine secretion. To test this hypothesis, we measured the protein levels of chemokines in wound homogenates. The secretion of the CC chemokines CCL2/MCP-1, CCL3/MIP-1α, CCL7/MCP-3, and CCL5/RANTES was significantly upregulated in the LPS-treated wounds, with the most dramatic change observed for CCL5/RANTES. In contrast, the protein levels of the CXC chemokines CXC2/MIP-2 and CXCL9/MIG were not altered after LPS treatment (Fig. 5a).
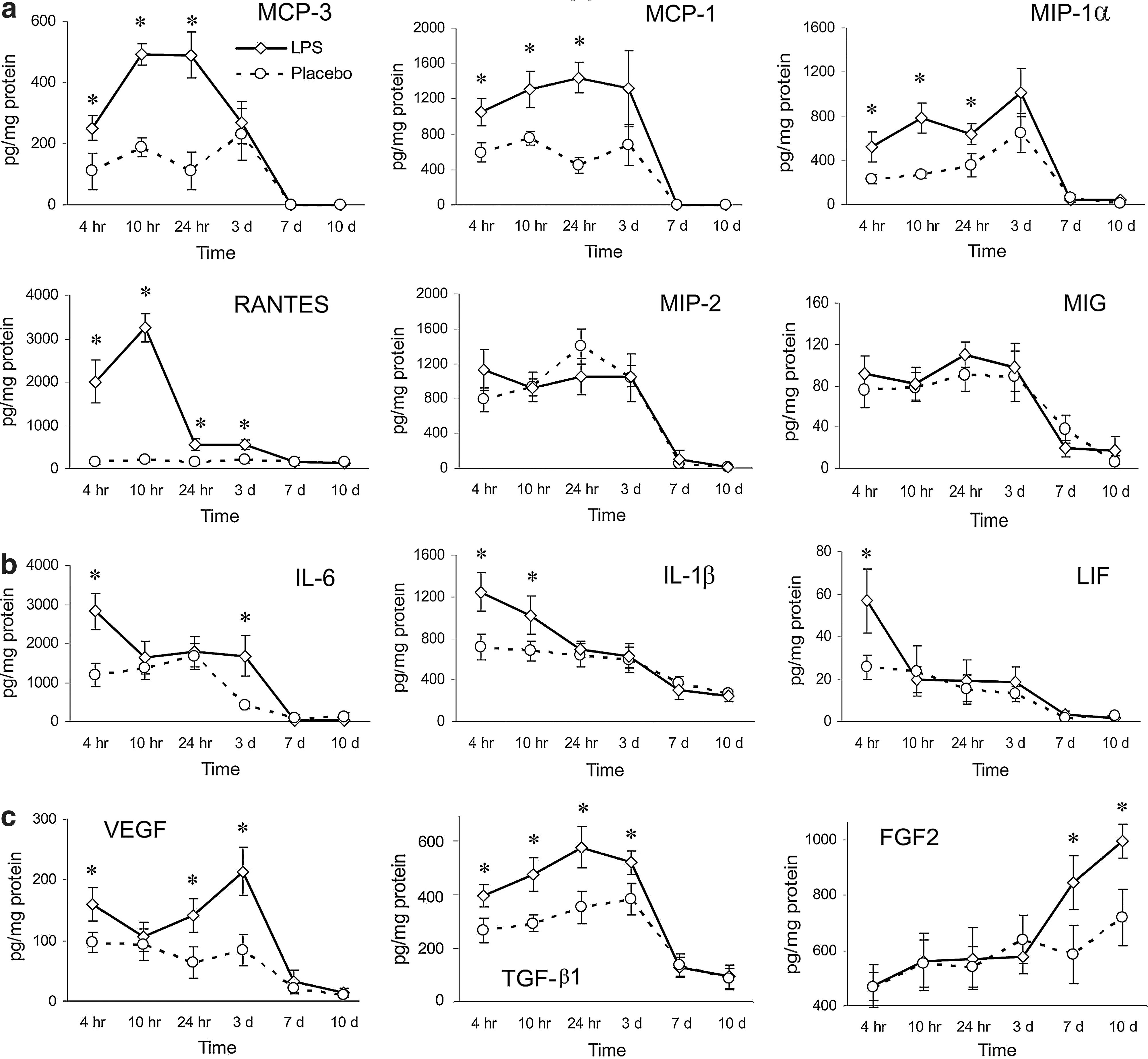
The effects of LPS wound treatment on the production of chemokines
To evaluate the changes in the wound inflammation process after LPS application, we examined the secretion of proinflammatory cytokines in the wounds. These cytokines are critical for the normal inflammatory response after skin injury (Werner and Grose 2003), and LPS is one of the most powerful inducers of proinflammatory cytokine production. LIF protein levels were upregulated 4 h after the first treatment, and IL-1β protein levels were upregulated 4 and 10 h after the first LPS treatment; however, no changes in these cytokine levels were detected at later time points. The secretion of IL-6 was upregulated after 4 h, and increased IL-6 protein levels were observed on day 3 (Fig. 5b).
To evaluate the secretion of growth factors that could be partially responsible for the improved wound healing, we measured the protein levels of TGF-β1, FGF2, and VEGF in the wound homogenates. The kinetics of the production of these growth factors in the wound homogenates after LPS or placebo treatment are shown in Fig. 5c.
Discussion
The innate immune system is involved in every aspect of the wound healing process and is especially significant during the inflammatory stage. A variety of cell types expressing innate immune receptors, including TLRs are involved in skin wound healing (Miller and Modlin 2007). Importantly, the ligands of these receptors are PAMPs, which are present in wounds because of the inevitable bacterial contamination of adult skin wounds, and damage-associated molecular patterns, which are present at the wound site as a result of tissue necrosis (Kluwe and others 2008).
Recent studies have shown that the topical activation of TLRs plays a significant role in the inflammatory stage of the repair process and results in the increased influx of macrophages, improved neovascularization of the wound bed, and accelerated wound healing (Deiters and others 2004; Sato and others 2010; Lin and others 2012). Other reports have demonstrated that the markedly slower skin wound healing observed in TLR3-deficient mice is associated with reduced chemokine expression and decreased accumulation of alternatively activated macrophages and neutrophils (Lin and others 2011). Impaired cutaneous wound healing, delayed granulation tissue formation, and reduced density of new blood vessels were also reported in MyD88-deficient mice (Macedo and others 2007). MyD88 is a signaling adaptor for nearly all of the TLRs (except TLR3). However, the influence of the topical application of bacterial LPS, a component of gram-negative bacteria and one of the most active ligands of innate immune receptors, on the processes of inflammation and wound healing has not yet been studied in detail.
In this study, we have described the ability of bacterial LPS to modulate inflammation during skin wound healing and have demonstrated its influence on the regeneration process.
Our findings demonstrate that topical LPS treatment increased the wound-breaking force in a dose-dependent manner. The mechanical strength of healing wounds is an integrative parameter and characterizes the speed at which wounds heal. Interestingly, histological analysis demonstrated that inflammation was resolved more rapidly in the LPS-treated mice than in the placebo-treated mice, suggesting accelerated wound healing after LPS treatment.
The enhanced secretion of proinflammatory cytokines (IL-6, IL-1β, and LIF) induced by LPS was detected only at early time points after LPS treatment; this upregulation was not as dramatic as that observed after the intramuscular injection of considerably smaller doses of LPS (data not shown). Therefore, the strength of innate immune signaling is determined by the route of administration of LPS. Probably because of its propensity to form micelles (Erridge and others 2002), LPS acts directly only during the early stages of wound healing (large micelles are unable to pass through the repaired skin barrier). Nevertheless, this short-term increase in proinflammatory cytokine secretion is able to influence various processes at the wound site. Proinflammatory cytokines mediate fibroblast chemotaxis, the degradation and synthesis of the extracellular matrix, keratinocyte and fibroblast proliferation, and angiogenesis (Werner and Grose 2003; Jee and others 2004).
The application of LPS resulted in increased secretion of CC-chemokines (CCL2/MCP-1, CCL7/MCP-3, CCL3/MIP-1α, and CCL5/RANTES) that play a key role in the recruitment of monocytes and macrophages to the wound site (Gillitzer and Gobeller 2001), and the increased secretion was correlated with an increased number of macrophages during the initial stage of healing. The recruitment of macrophages into wound tissues is an important step in the inflammatory response. Macrophages are considered to orchestrate the wound-healing process (Park and Barbul 2004). They prevent infection and debride the wound through phagocytosis, the generation of reactive radicals, and the production of proteolytic enzymes, and they stimulate the processes of neovascularization, fibroplasia, and re-epithelialization. Macrophages are a source of numerous cytokines and growth factors that act in a paracrine manner or recruit and activate fibroblasts, endothelial cells, epithelial cells, and other cell types (Mahdavian Delavary and others 2011). Improved macrophage recruitment may be one of the causes of improved wound healing. We did not detect increased synthesis of the CXC-chemokine CXCL2/MIP-2 (MIP-2 is a potent regulator of neutrophil chemotaxis) (Gillitzer and Gobeller 2001). This phenomenon correlated with the absence of any changes in neutrophil infiltration during the early period of wound healing.
Our findings show that the topical application of bacterial LPS upregulated the secretion of a number of angiogenesis mediators, including VEGF. Recent in vitro studies have shown that LPS acts synergistically with agonists of adenosine A2a receptors (which are naturally present at the wound site) to significantly upregulate the secretion of VEGF by macrophages (Leibovich and others 2002). Interestingly, LPS was reported to strongly induce the expression of A2a adenosine receptors on macrophages (Murphree and others 2005). In addition to the increased secretion of VEGF, topical LPS treatment upregulated the secretion of CC-chemokines that are known to be mediators of angiogenesis (CCL2/MCP-1 and CCL3/MIP-1α). Endothelial cells express CCR2, the receptor for MCP-1, and migrate toward MCP-1 (Weber and others 1999). MCP-1 was found to be a potent inducer of angiogenesis in vivo and in ovo, and this effect is independent of its leukocyte chemotactic properties (Goede and others 1999; Salcedo and others 2000). Recent reports show that MCP-1 acts as an angiogenic factor by upregulating the hypoxia-inducible factor 1α, which induces VEGF expression (Hong and others 2005). Another angiogenic chemokine, MIP-1α, stimulates the production of VEGF in an alkali-induced cornea injury (Lu and others 2008). The depletion of MIP-1α from the wound using a neutralizing anti-serum leads to a significant decrease in angiogenic activity and collagen synthesis (DiPietro and others 1998). Our study showed increased levels of the FGF2, a potent angiogenic mediator that stimulates endothelial cell proliferation as well as the physical organization of endothelial cells into tube-like structures and regulates the expression of various extracellular matrix components that contribute to the maturation of new blood vessels (Presta and others 2005). Considering the upregulation of a number of angiogenic mediators after LPS wound treatment, it may be assumed that LPS stimulates wound angiogenesis; however, this question requires further study.
Our study showed that the topical application of LPS promoted collagen synthesis and remodeling and was correlated with increased levels of TGF-β1 and FGF2, potent profibrotic factors involved in the process of skin repair (Werner and Grose 2003; Barrientos and others 2008). It was previously reported that the in vitro treatment of intestinal fibroblasts with LPS enhanced the NF-κB promoter activity and increased collagen contraction (Burke and others 2010). Moreover, LPS was shown to induce the secretion of collagen in mouse lung fibroblasts by directly activating the phosphoinositide 3-kinase-Akt (PI3K-Akt) pathway through TLR4 in vitro (He and others 2009). However, the improved collagen synthesis that we observed was probably rather a consequence of early events during repair process, such as improved macrophage migration to the wound site or upregulated proinflammatory cytokine secretion. Additional experiments are necessary to establish the precise mechanism underlying this phenomenon.
It is likely that multiple mechanisms account for the observed LPS-induced acceleration of wound repair, as wound healing is a very complex process involving a cascade of various biochemical and cellular events. Nevertheless, we hypothesize that a simplified mechanism of the observed phenomenon exists, based on our findings. (1) LPS interacts with TLR4, which is expressed on a wide range of cells in the wound-defect area, and induces a cascade of reactions that ultimately lead to the induction of proinflammatory cytokine secretion (IL-6, IL-1β, and LIF). (2) The LPS-induced secretion of monocyte/macrophage chemoattractants, particularly the CC-chemokines (CCL2/MCP-1, CCL7/MCP-3, CCL3/MIP-1α, and CCL5/RANTES), and proinflammatory cytokines enhances monocyte and macrophage migration into the wounded area and promotes monocyte differentiation into macrophages. (3) Macrophages have been shown to play a critical role in wound healing. LPS-stimulated macrophages and possibly other types of cells in the wound produce a number of molecules that regulate the repair process and the development of connective tissue, such as FGF2, VEGF, TGF-β1, and EGF. (4) The above sequence of events ultimately leads to improved wound healing.
Taken together, our findings demonstrate the robust, complex effect of LPS treatment on the wound-healing process. LPS treatment resulted in accelerated resolution of inflammation, increased macrophage infiltration, and enhanced collagen synthesis; these effects were associated with changes in the secretion of a number of mediators that are involved in the skin regeneration process.
Footnotes
Acknowledgment
We thank Dr. D.V. Prosvirnin for providing skillful technical assistance with the biomechanical tests.
Author Disclosure Statement
No competing financial interests exist.