
Abstract
Interferon-gamma (IFN-γ) is a paracrine inhibitor of melanocytes and genetic variability due to intron 1 polymorphisms in IFNG has been reported to be associated with increased risk for several autoimmune diseases. The aim of present study was to determine whether intron 1 +874A/T (rs2430561) and CA microsatellite (rs3138557) polymorphisms in IFNG are associated with generalized vitiligo (GV) susceptibility and expression of IFNG and intercellular adhesion molecule-1 (ICAM1) affects the disease onset and progression. Here we report that IFNG CA microsatellite but not +874A/T may be a genetic risk factor for GV; however, +874T allele plays a crucial role in increased expression of IFNG mRNA and protein levels which could affect the onset and progression of the disease. Active GV patients showed increased IFNG levels compared to stable GV patients. The genotype-phenotype analysis revealed that IFNG expression levels were higher in patients with +874 TT genotypes and 12 CA repeats. Patients with the early age of onset showed higher IFNG expression and female GV patients showed higher IFNG and ICAM1 expression implicating gender biasness and involvement of IFN-γ in early onset of the disease. Moreover, the increased IFN-γ levels in patients lead to increased ICAM1 expression, which could be a probable link between cytokines and T-cell involvement in pathogenesis of GV.
Introduction
G
More than 30 susceptibility, loci have been identified for GV on the basis of genetic linkage and Genome-wide association studies (GWAS) (Jin and others 2012). All but one of these genes encodes proteins involved in regulation of the immune system and/or have been genetically associated with susceptibility to other autoimmune diseases. The sole exception is TYR, encoding tyrosinase, the key enzyme of melanin biosynthesis and the principal vitiligo autoimmune antigen. These genes together account for a relatively small fraction of the genetic risk factors of GV, indicating that many additional vitiligo susceptibility genes undoubtedly remain to be discovered. Moreover, studies by Birlea and others (2013) and Singh and others (2012) in Indian population, support the association of GV with loci in the MHC class II region; however, GWAS in the EUR population vitiligo show primary association with HLA-A in the distal class I region (Jin and others 2010, 2011) in addition to the association of multiple signals in the MHC class II region. Additionally, GWAS studies in the Chinese population show principal MHC association in the class III region (Quan and others 2010) and in the proximal class I region, between HLA-B and HLA-C (Liu and others 2012). These differences and similarities in MHC genetic association in different populations may partly responsible for the differing prevalence of this autoimmune disease in different groups around the world.
Cytokines are small immune-regulatory molecules which can generate an inappropriate immune response due to their imbalance (Feldmann and others 1998). Moretti and others (2002a, 2002b) have shown cytokine imbalance in skin of vitiligo patients suggesting their role in autoimmunity. Interferon-gamma (IFN-γ, a type II interferon) is a pleiotropic cytokine and is a key regulator of immune system (Schroder and others 2004). The principal sources of IFN-γ are helper T-cells and natural killer cells, although many other cells can produce it, including keratinocytes. Apart from host defense, IFN-γ also contributes to autoimmune pathology. IFN-γ is suggested to cause induction of antimelanocyte antibodies or activation of cytotoxic T-cells (Miller and others 2002; SeÈkin and others 2004). It has been proposed that melanocyte death is mediated by apoptosis in the context of autoimmunity, and cytokines, such as IFN-γ and tumor necrosis factor (TNF)-α can initiate apoptosis (Huang and others 2002). Additionally, IFN-γ and TNF-α induce the expression of intercellular adhesion molecule-1 (ICAM1) on the cell-surface of melanocytes (Yohn and others 1990). Increased expression of ICAM1 on the melanocytes enhances T cell- melanocyte attachment in the skin and may be involved in the destruction of melanocytes in vitiligo (Al Badri 1993). Kocer and others (2009) suggested that IFN-γ therapy may cause vitiligo. These reports signify that IFN-γ may play a crucial role in vitiligo susceptibility. Therefore, it is possible that individuals who naturally produce higher levels of IFN-γ will exhibit different susceptibility, or severity towards GV.
IFNG gene is located on human chromosome 12q24 spanning ∼5.4 kb, consisting of 4 exons and 3 introns (Gray and Goeddel 1982; Naylor and others 1983). The IFNG coding region is invariant, with no reported polymorphisms (Hayden and others 1997). However, there are 2 well-known single nucleotide polymorphisms (SNPs) in the IFNG gene noncoding region (intron 1): [+874A/T polymorphism (rs2430561); CA microsatellite (rs3138557)]. Allele 2, with 12 CA repeats is associated with in vitro constitutive high IFN-γ production. In addition, +874A/T SNP at the 5′ end of the CA repeat region has been correlated with the presence or absence of the microsatellite allele 2 (Pravica and others 1999). Also, the presence of IFNG (+874Alo/Thi) polymorphism creates a putative nuclear factor-κB (NF-κB) binding site and shows preferential binding to the T allele and correlates with high IFN-γ producer phenotype (Pravica and others 2000).
In the present study, we have made an attempt to understand the role of IFN-γ and ICAM1 in pathogenesis of GV. Hence, the objectives of this study were, (1) to determine whether the intron 1 polymorphisms of IFNG [+874A/T (rs2430561) and 5′ end CA microsatellite (rs3138557)] are associated with GV susceptibility; (2) to measure and compare IFNG and ICAM1 transcripts and serum IFN-γ levels in GV patients and controls; (3) to correlate IFNG polymorphisms/levels with onset and progression of the disease.
Materials and Methods
Study subjects
The study group included 517 GV patients (including acrofacial vitiligo and vitiligo universalis) who referred to S.S.G. Hospital, Vadodara, India and B.J. Medical College and Civil hospital, Ahmedabad, India (Supplementary Table S1; Supplementary Data are available online at
The patients were divided into 2 groups based on whether the existing lesions were spreading and/or new lesions had appeared within the previous 6 months: an affirmative answer to one or both of those questions led to inclusion of the patient in the progressive (active) GV group, whereas patients with no increase in lesion size or number were included in the stable GV group. Three hundred eighty-two patients were classified with progressive GV, whereas 135 patients were included in the stable GV group (Supplementary Table S1).
A total of 881 age matched unaffected individuals were included as controls in the study (Supplementary Table S1). Demographic data for patients and controls are shown in Supplementary Table S1. None of the healthy individuals or their relatives had any evidence of vitiligo and autoimmune disease. The study plan was approved by the Institutional Ethics Committee for Human Research (IECHR), Faculty of Science, The Maharaja Sayajirao University of Baroda, Vadodara, Gujarat, India. The importance of the study was explained to all participants and written consent was obtained from all subjects before performing the studies.
Genomic DNA preparation
Five milliliters venous blood was collected from the patients and unaffected subjects in K3EDTA coated vacutainers (BD). Genomic DNA was extracted from whole blood using “QIAamp DNA Blood Kit” (Qiagen, Inc.) according to the manufacturer's instructions.
Genotyping of +874A/T SNP of IFNG
IFNG +874A/T genotyping was done using amplification refractory mutation system–polymerase chain reaction (ARMS-PCR) method. DNA was amplified in 2 different PCRs with a generic antisense primer and one of the 2 allele specific sense primers (Supplementary Table S2). To assess the success of PCR amplification in both the reactions, an internal control of 426 bp was amplified using a pair of primers designed from the nucleotide sequence of the human growth hormone (HGH) (Supplementary Table S2). The reaction mixture of the total volume of 15.1 μL included 5 μL (100 ng) of genomic DNA, 5.0 μL nuclease-free H2O, 1.5 μL 10×PCR buffer, 0.3 μL 8 mM dNTPs (Sigma Chemical Co.), 0.3 μL of 10 μM allele-specific, and common primers (MWG Biotech), 0.3 μL of 2 μM control primers (HGH), 1.5 μL Mg2+ and 0.6 μL (5 U/μL) Taq Polymerase (Bangalore Genei). Amplification was performed using a PTC-100 thermal cycler (MJ Research, Inc.) according to the protocol: 95°C for 1 min followed by 10 cycles of 95°C for 15 s, 62°C for 50 s, and 72°C for 40 s, then 20 cycles of 95°C for 20 s, 56°C for 50 s, and 72°C for 50 s.
The amplified products were separated by electrophoresis on 2.0% agarose gels and visualized under a UV transilluminator with a 100 base pair DNA ladder (Bioron). Two amplicons were available for each sample (one each specific for A or T allele of the IFNG gene). More than 10% of the samples were randomly selected for confirmation and the results were 100% concordant (analysis of the chosen samples was repeated by 2 researchers independently and also confirmed by DNA sequencing).
Genotyping of CA microsatellite (rs3138557) of IFNG
IFNG CA microsatellite (rs3138557) was genotyped by High Resolution Melt (HRM) curve analysis using LightCycler® 480 Real-Time PCR protocol. The primers used for the genotyping are mentioned in Supplementary Table S2. Real-time PCR was performed in 20 μL volume using LightCycler 480 HRM Master (Roche Diagnostics GmbH) following the manufacturer's instructions. The thermal cycling conditions included an initial activation step at 95°C for 10 min, followed by 45 cycles of denaturation, annealing, and amplification (95°C for 15 s, 56°C for 20 s, 72°C for 20 s). The fluorescence data collection was performed during the extension step. At the end of the amplification phase, a HRM curve analysis was carried out on the product formed as per the manufacturer's instructions. DNA sequencing was carried out for samples from each representative groups obtained by HRM analysis.
Determination of IFNG, ICAM1 and GAPDH mRNA expression
RNA extraction and cDNA synthesis
Total RNA from whole blood was isolated and purified using Ribopure™-blood Kit (Ambion, Inc.) following the manufacturer's protocol. RNA integrity was verified by 1.5% agarose gel electrophoresis, RNA yield, and purity was determined spectrophotometrically at 260/280 nm. cDNA synthesis was performed using 1 μg of total RNA by Thermo Scientific VersoTM cDNA Synthesis Kit (Thermo Fisher Scientific, Inc.) according to the manufacturer's instructions.
Real-time PCR
The expression of IFNG, ICAM1, and GAPDH transcripts were measured by real-time PCR using gene specific primers (Eurofins) as shown in Supplementary Table S2. Expression of GAPDH gene was used as a reference. Real-time PCR was performed in duplicates in 20 μL volume using LightCycler 480 SYBR Green I Master (Roche Diagnostics GmbH) following the manufacturer's instructions. The thermal cycling conditions included an initial activation step at 95°C for 10 min, followed by 45 cycles of denaturation, annealing and amplification (95°C for 10 s, 65°C for 15 s, 72°C for 20 s). The fluorescence data collection was performed during the extension step. At the end of the amplification phase, a melt curve analysis was carried out to check the specificity of the products formed (Supplementary Fig. S1). The PCR cycle at which PCR amplification begins its exponential phase and product fluorescence intensity finally rises above background and becomes visible was considered as the crossing point (CP) or cycle threshold (CT). The ΔCT or ΔCP value was determined as the difference between the cycle threshold of target gene (IFNG/ICAM1) and reference gene (GAPDH). The difference between the 2 ΔCP values (ΔCP Controls and ΔCP patients) was considered as ΔΔCP to obtain the value of fold change (2−ΔΔCp).
Estimation of serum IFN-γ levels
Serum levels of IFN-γ in patients with GV and controls were measured by enzyme-linked immunosorbent assay (ELISA) using the Immunotech Human IFN-γ ELISA kit (Immunotech SAS) as per the manufacturer's protocol.
Statistical analyses
Evaluation of the Hardy–Weinberg equilibrium (HWE) was performed in patients and controls by comparing the observed and expected frequencies of the genotypes using chi-squared analysis. The distribution of the genotypes and allele frequencies of IFNG polymorphisms for patients and control subjects were compared using chi-squared test with 3×2 and 2×2 contingency tables, respectively using Prism 4 software (Graphpad Software, Inc.). P values less than 0.01 were considered as statistically significant due to Bonferroni's correction for multiple testing. Odds ratio with respective confidence interval (95% CI) for disease susceptibility was also calculated. Age of onset analysis, relative gene expression of IFNG, and serum IFN-γ levels in patient and control groups were plotted and analyzed by nonparametric unpaired t-test using Prism 4 software (Graphpad Software, Inc.).
Results
Analysis of association between intron 1 +874A/T (rs2430561) polymorphism of IFNG and susceptibility to GV
ARMS-PCR for the IFNG (+874A/T) polymorphism revealed a 428 bp product for HGH as a control gene, and a 265 bp product for IFNG T874 (homozygous for allele T; TT), or A874 (homozygous for allele A; AA), or both alleles T and A (heterozygous; TA). The genotypes for IFNG intron1 +874A/T polymorphism were confirmed by running PCR amplicons on 2.0% agarose gel electrophoresis (Supplementary Fig. S2).
Intron 1 +874A/T polymorphism of IFNG gene was not found to be associated with GV patients (P=0.485) when genotypes were compared with chi-squared test-3×2 contingency table (Table 1). Also, there was no significant difference in allele frequencies of this polymorphism between patients and controls when compared with 2×2 contingency table (P=0.274) (Table 1). Both patients and control population were found to be in HWE for this polymorphism (P=0.942 and P=0.847, respectively) (Table 1). Interestingly, the frequency of minor ‘T’ allele was significantly higher in active cases as compared to stable cases of GV (45% versus 31%; P=0.0003) (Table 2). In addition, genotype frequencies and allele frequencies for this polymorphism were also analyzed between male and female patients with GV and controls, which revealed no significant difference as detected by chi-squared test (P=0.459 and P=0.030, respectively) (Table 3).
Values are significant at P≤0.01 due to Bonferroni's correction.
‘n’ represents number of patients/controls.
Vitiligo Patients versus controls using chi-squared test with 3×2 contingency table.
Vitiligo Patients versus controls using chi-squared test with 2×2 contingency table.
GV, generalized vitiligo; HWE, Hardy–Weinberg equilibrium; CI, confidence interval; Ps, patients; C, controls.
Values are significant at P≤0.01 due to Bonferroni's correction.
‘n’ represents number of patients.
Active patients versus stable patients using chi-squared test with 3×2 contingency table.
Active patients versus stable patients using chi-squared test with 2×2 contingency table.
AV, active vitiligo patients; SV, stable vitiligo patients.
Values are significant at P≤0.01 due to Bonferroni's correction.
‘n’ represents number of patients.
Male patients versus female patients using chi-squared test with 3×2 contingency table.
Male patients versus female patients using chi-squared test with 2×2 contingency table.
M, male patients; F, female patients.
Analysis of association between intron 1 CA microsatellite (rs3138557) polymorphism of IFNG and susceptibility to GV
HRM curve analysis for CA microsatellite (rs3138557) polymorphism was performed using LightCycler 480 Real-Time PCR. The 5 different CA repeats identified were, 12–12 CA, 13–13 CA, 12–13 CA, 12–14 CA, and 13–14 CA, based on normalized and temperature shifted difference plots followed by DNA sequencing of samples from each representative group (Supplementary Figs. S3 and S4).
The distribution of the IFNG microsatellite polymorphism (CA) was different between cases and controls (P<0.001). The genotype (CA) 12–12, that is, the homozygous state for allele 2 was significantly more frequent (P=0.005) in GV patients (57.0%) than in controls (23.0%), whereas the genotype (CA) 12–13 recurred less frequently in the patients (10.0%) versus controls (42%; P=0.005) (Table 4). The allelic frequency differed significantly between cases and controls (P=0.004). In GV patients, the allele 12 was significantly (P=0.004) more frequent (66.0%) than in controls (48.0%), whereas the allele 13 recurred less frequently (28.0%) than in controls (46.0%; P=0.005) (Table 4). The results suggest significant association of IFNG 12–12 CA repeats with GV susceptibility.
Values are significant at p≤0.01 due to Bonferroni's correction.
Comparison of the distribution of genotypes between the 2 groups.
Comparison of the single genotype/allele against all the others.
NS, not significant.
Effect of IFNG +874A/T polymorphisms on age of onset of GV
When age of onset of the disease was correlated with the IFNG +874A/T genotypes, patients with higher (IFN-γ) producer TT genotypes showed an early onset of the disease as compared to AA and AT genotypes (P<0.0001 and P=0.004, respectively) (Fig. 1A). Moreover, patients with genotype AT showed an early onset of the disease as compared to AA genotypes (P=0.011) (Fig. 1A) suggesting the effect of the susceptible allele ‘T’ on the early onset of disease.
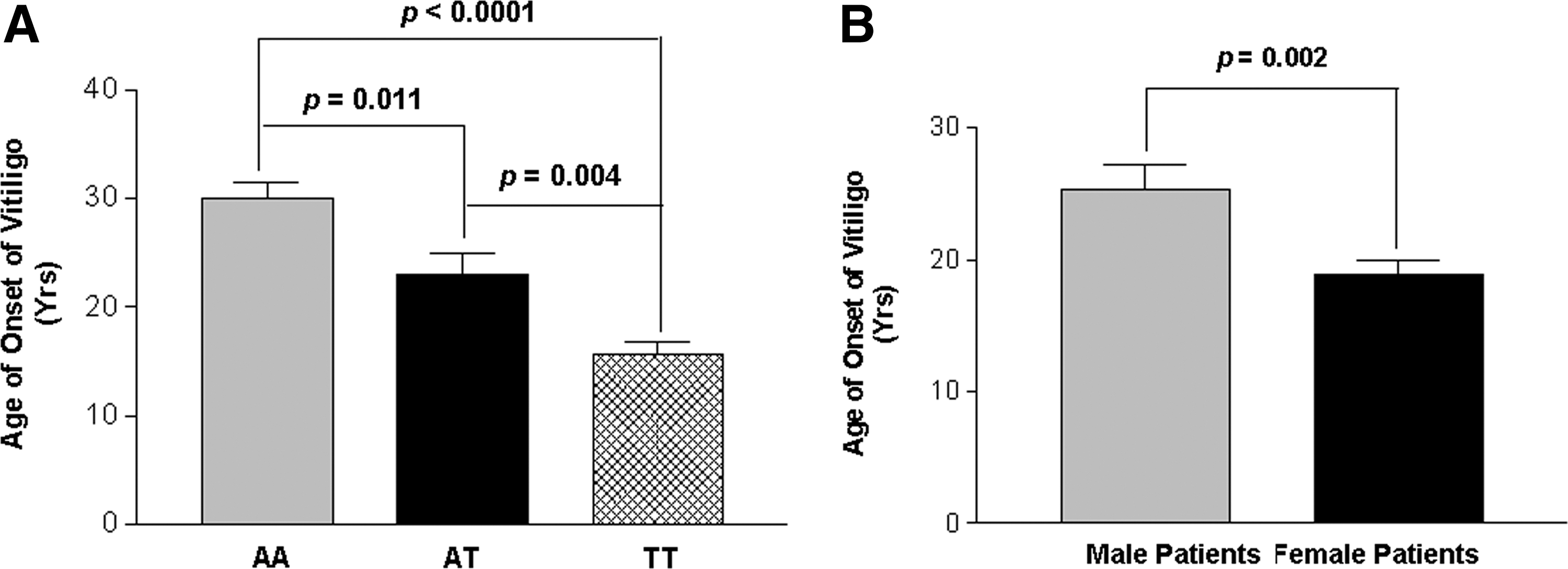
Age of onset of the disease in generalized vitiligo (GV) patients.
Interestingly, when male and female GV patients were analyzed for age of onset of the disease, female patients had significant early onset of the disease as compared to the male patients (P=0.002) (Fig. 1B).
Relative gene expression of IFNG in patients with GV and controls
Comparison of the findings showed significant increase in expression of IFNG in 122 GV patients than in 175 unaffected controls after normalization with GAPDH expression as suggested by mean ΔCp values (P<0.0001) (Fig. 2A). The 2−ΔΔCp analysis showed ∼0.047-fold change in the expression of IFNG transcript in patients as compared to controls (Fig. 2B).
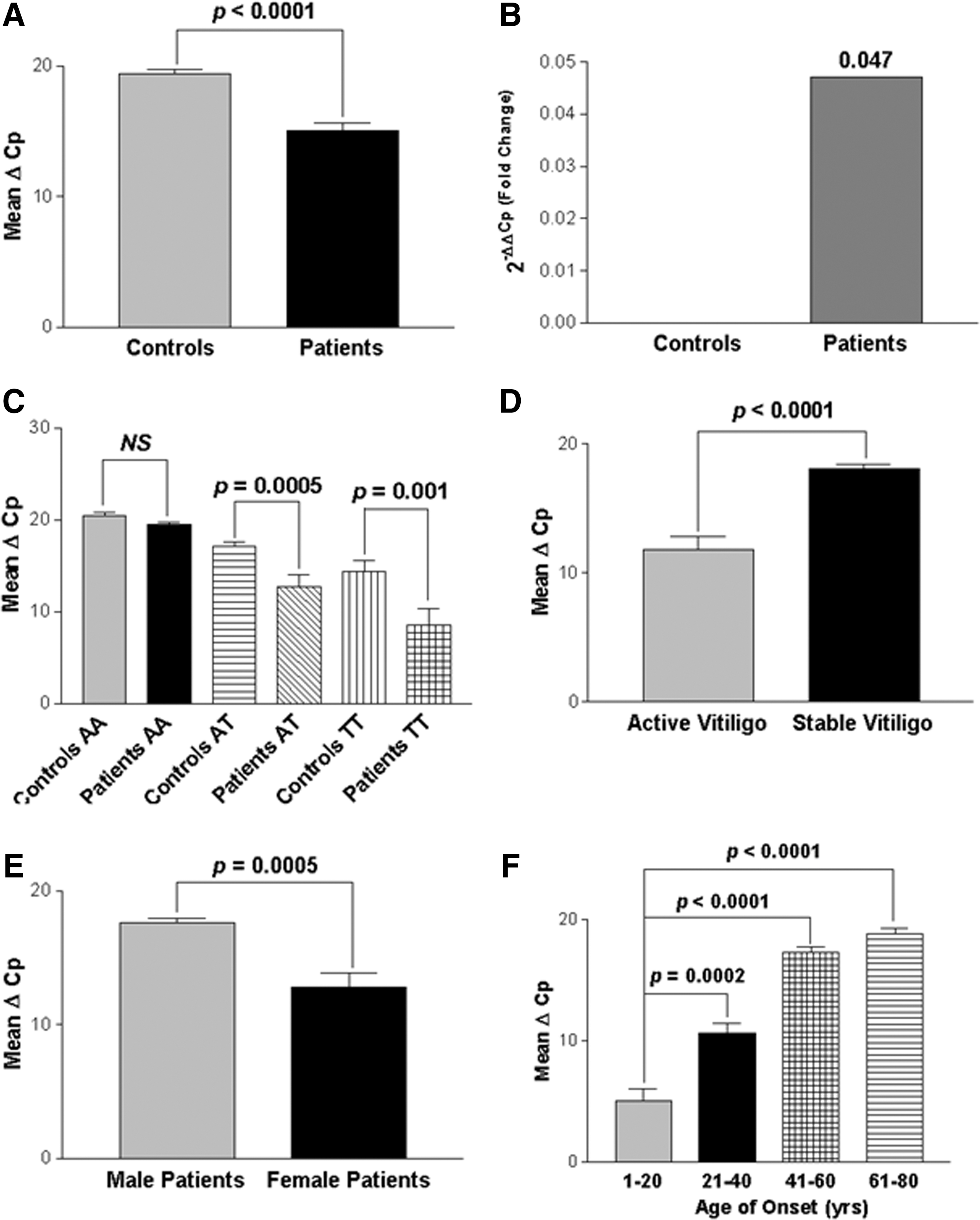
Relative gene expression of IFNG in controls and GV patients.
Correlation of IFNG transcripts with +874A/T polymorphism
Further, the expression levels of IFNG were analyzed with respect to +874A/T (rs2430561) genotypes (Fig. 2C). Interestingly, IFNG expression was significantly increased in patients with susceptible TT genotypes as compared to controls (P=0.001). Also, patients with genotypes AT showed increased IFNG transcripts as compared to controls (P=0.0005); however, no significant difference was observed in IFNG expression in patients as compared to controls with AA genotypes (P=0.057).
Effect of IFNG expression on disease progression
In addition, we also checked the effect of IFNG expression on progression of the disease, that is, active and stable cases (Fig. 2D). Interestingly, active GV patients showed significant increase in expression of IFNG transcripts as compared to the patients with stable GV (P<0.0001) suggesting the involvement of IFN-γ in disease progression. Moreover, the susceptibility of the disease was also checked based on the gender differences and we found that female patients showed significantly higher IFNG expression as compared to male patients (P=0.0005) (Fig. 2E).
When IFNG expression was monitored in different age of onset groups, patients with age of onset group: 1–20 years showed significantly increased expression of IFNG transcripts as compared to the age of onset groups: 21–40, 41–60, and 61–80 years (P=0.0002, P<0.0001, and P<0.0001, respectively), suggesting the importance of IFN-γ in early onset of the disease (Fig. 2F).
Functional correlation of IFNG +874A/T and CA microsatellite (rs3138557) polymorphisms with its levels in the serum
To find any functional correlation of the investigated IFNG promoter polymorphisms with its level in the serum, IFN-γ levels were measured in 214 GV patients and 236 unaffected controls. GV patients showed significant increased serum IFN-γ levels as compared to controls (P=0.002) (Fig. 3A). Moreover, when the patient groups were analyzed based on disease progression with respect to IFN-γ levels, patients with active GV had significantly higher IFN-γ levels as compared to stable GV (P=0.003) (Fig. 3B).
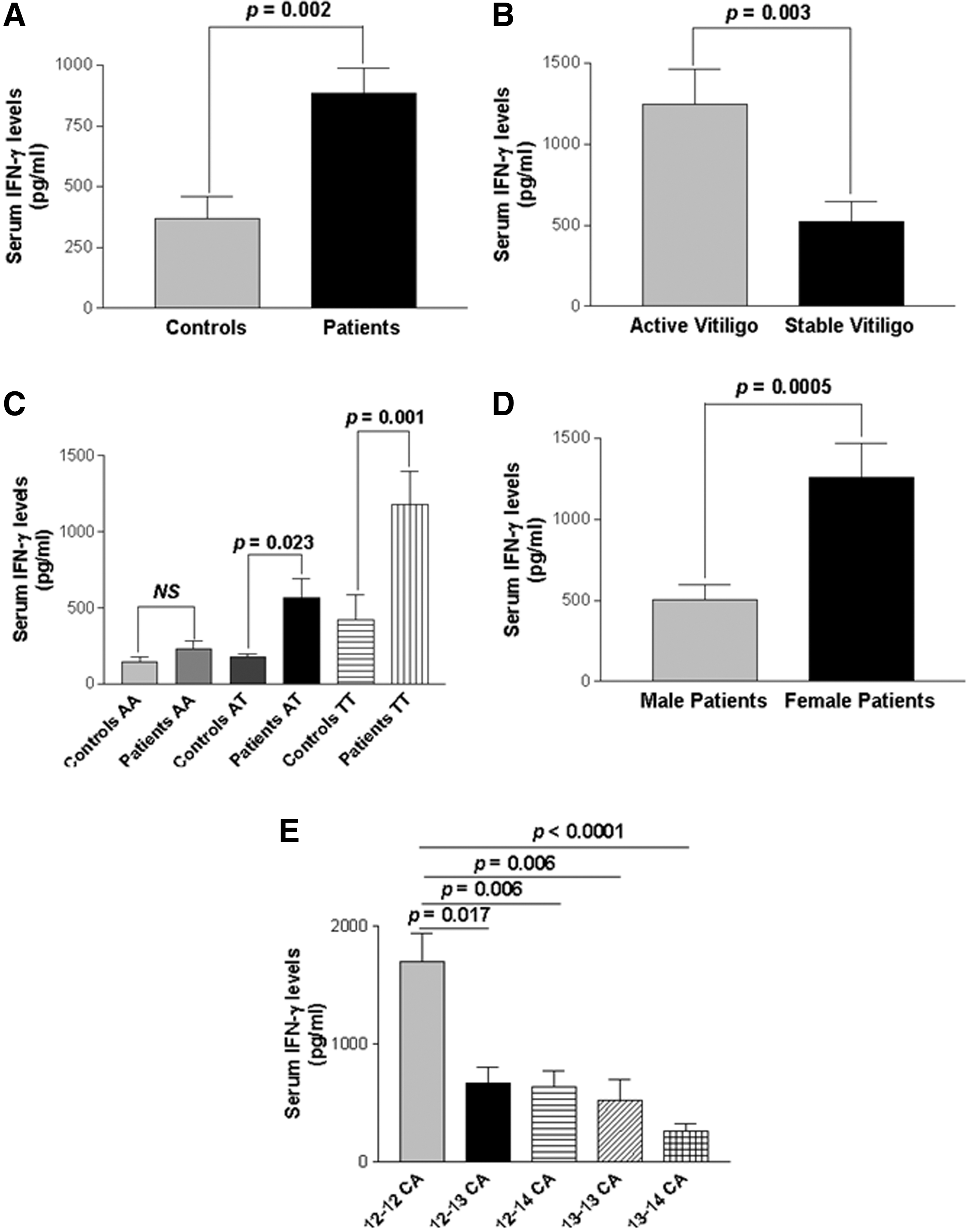
Serum interferon-γ (IFN-γ) levels in controls and GV patients.
Furthermore, when +874A/T genotypes were analyzed with IFN-γ levels, patients with TT and AT genotypes showed significantly higher levels of IFN-γ as compared to those of controls (P=0.001, P=0.023) (Fig. 3C). However, patients with AA genotype showed no significant difference in IFN-γ levels as compared to those of controls (P=0.521). Additionally, when the male and female patients were analyzed with respect to IFN-γ levels, female patients showed significantly higher levels of IFN-γ as compared to male patients (P=0.0005) (Fig. 3D).
Further, the 5 different CA repeats of IFNG were correlated with serum IFN-γ levels and interestingly, the 12–12 CA repeat showed significantly higher levels of IFN-γ as compared to longer CA repeats, that is, 12–13, 12–14, 13–13, and 13–14 (P=0.017, P=0.006, P=0.006 and P<0.0001) (Fig. 3E) suggesting the positive correlation of IFNG 12–12 CA repeats with higher levels of IFN-γ.
Relative gene expression of ICAM1 in patients with GV and controls
Comparison of the findings showed significant increase in expression of ICAM1 transcripts in 122 GV patients than in 175 unaffected controls after normalization with GAPDH expression as suggested by mean ΔCp values (P=0.008) (Fig. 4A). The 2−ΔΔCp analysis showed ∼0.369-fold change in the expression of ICAM1 transcript in patients as compared to controls (Fig. 4B).
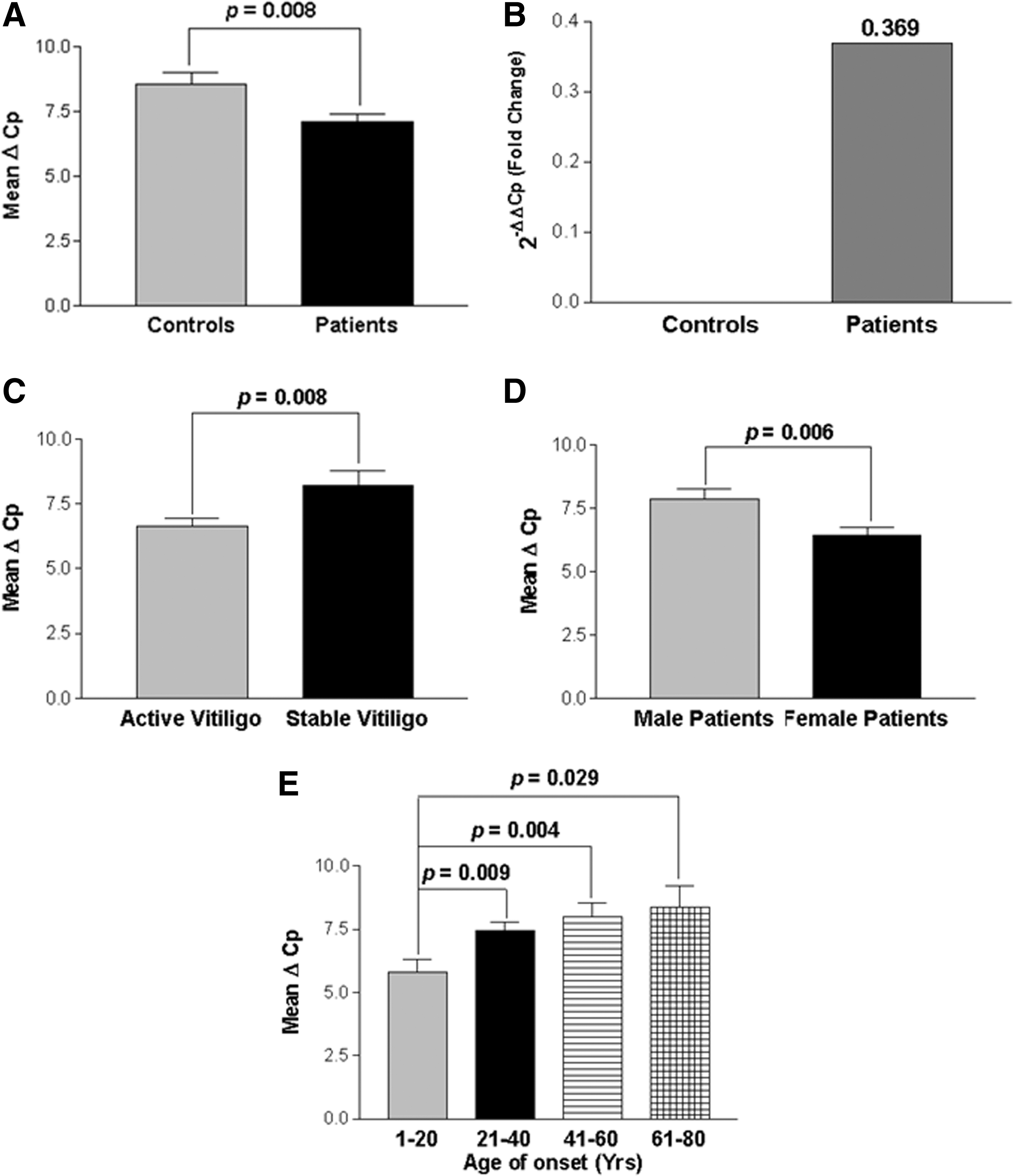
Relative gene expression of intercellular adhesion molecule-1 (ICAM1) in controls and GV patients.
In addition, the effect of ICAM1 expression on progression of the disease, that is, active and stable cases (Fig. 4C) revealed that active GV patients had significantly increased expression of ICAM1 transcripts as compared to patients with stable GV, (P=0.008) suggesting the involvement of ICAM1 in disease progression. Moreover, the susceptibility of the disease was also checked based on the gender differences and we found that female patients with GV showed significantly higher ICAM1 expression as compared to male patients (P=0.006) (Fig. 4D). When ICAM1 expression was monitored in different age of onset groups of patients, patients with age of onset group: 1–20 years showed significantly increased expression of ICAM1 transcripts as compared to the age of onset groups: 21–40, 41–60, and 61–80 years (P=0.0002, P<0.0001 and P<0.0001, respectively) suggesting the importance of ICAM1 in early onset of the disease (Fig. 4E).
Discussion
Vitiligo cannot be explained by simple Mendelian genetics; however, it is characterized by incomplete penetrance, multiple susceptibility loci, and genetic heterogeneity (Zhang and others 2005). We have earlier reported that 21.93% of Gujarat vitiligo patients exhibit positive family history and 13.68% patients have at least one first-degree relative affected (Shajil and others 2006a). The present study also shows that 12.57% of GV patients have one or more first degree relative affected, suggesting the involvement of genetic factors in pathogenesis of GV. Recently, we have shown positive association of HLA-A*33:01, HLA-B*44:03, and HLA-DRB1*07:01 with vitiligo patients from North India and Gujarat suggesting an autoimmune link of vitiligo in these cohorts (Singh and others 2012). We also showed that the 3 most significant class II region SNPs: rs3096691 (just upstream of NOTCH4), rs3129859 (just upstream of HLA-DRA), and rs482044 (between HLA-DRB1 and HLA-DQA1) are associated with GV (Birlea and others 2013). The genotype-phenotype correlation of CTLA4, IL-4, and TNFA MYG1 polymorphisms supported the autoimmune pathogenesis of vitiligo in Gujarat population (Dwivedi and others 2011; Imran and others 2012; Laddha and others 2012; Dwivedi and others 2013a), whereas our earlier studies on CAT, GPX, MBL2, ACE, PTPN22 polymorphisms did not show significant association (Shajil and others 2007; Dwivedi and others 2008, 2009; Laddha and others 2008).
Autoimmunity has been suggested to play a major role in the pathogenesis of GV (Nordlund and others 2006). The autoimmune destruction of melanocytes can be explained by the abnormalities in both humoral and cell-mediated immunity (Kemp and others 2001; Shajil and others 2006b; Laddha and others 2013). Melanocyte-specific circulating autoantibodies (Cui and Bystryn 1995; Kemp and others 1997), autoreactive CD8+ cytotoxic T-cells and macrophages (Le Gal and others 2001; Mandelcorn-Monson and others 2003) that recognize pigment cell antigens have been detected in the sera of a significant proportion of vitiligo patients. In particular, active cases of vitiligo were demonstrated to have higher levels of autoantibodies and cytotoxic T-cells (Le Poole and Luiten 2008). Our recent study has also demonstrated circulating autoantibodies in the sera of 75% of Gujarat vitiligo patients as compared to unaffected individuals (Unpublished data). The autoimmune hypothesis gains further support from immunotherapy studies of melanoma patients (Rosenberg 1997). Twenty-six percent of melanoma patients responded to IL-2 based immunotherapy, developed vitiligo suggesting that antimelanotic T-cells which might be responsible for melanoma regression may also be linked to the destruction of normal melanocytes in vitiligo (Zeff and others 1997). Moreover, in vitro direct analysis of vitiliginous skin margins showed the presence of polarized CD4+ and CD8+ T-cells, which predominantly secrete IFN-γ and TNF-α might be associated with the destruction of melanocytes during active disease (Wańkowicz-Kalińska and others 2003). Our recent study has also shown dramatic increase in CD8+ T-cell number and significant decrease in Tregs number in circulation of active GV patients (Dwivedi and others 2013b).
Transcript and protein levels of cytokines depend on both genetic and environmental factors (Gottenberg and others 2004). Analysis of cytokines gene polymorphisms would be able to detect a genetic abnormality of cytokine regulation that might play a role in the pathophysiology of the disease. Therefore, cytokines encoding genes could be considered as the candidate genes for vitiligo susceptibility. Pravica and others (1999) reported that the IFNG +874A/T SNP directly affects the level of IFN-γ production and correlates with the presence of the ‘A’ allele and low production of IFN-γ as compared to ‘T’ allele. Further, the study suggested that this polymorphism (‘T’ allele) coincided with a putative NF-κB binding site that could have functional consequences for transcription of the human IFNG gene, which could directly increase the level of IFN-γ production (Pravica and others 2000). The present study showed significant increase in IFNG mRNA and protein levels in GV patients as compared to controls. An imbalance of cytokines in the epidermal microenvironment of lesional vitiligo skin has been demonstrated, which could impair the life span and function of melanocytes (Moretti and others 2002a, 2002b; Grimes and others 2004). Recent studies have shown an increased IFNG mRNA levels in patient's involved skin (Grimes and others 2004; Schoenborn and Wilson 2007; Wang and others 2011; Reimann and others 2012) and also in PBMCs (Reimann and others 2012) compared to controls. Moreover, animal model studies for vitiligo have also confirmed the role of IFN-γ in vitiligo pathogenesis (Gregg and others 2010). Recently, increased expression of IFNG has been observed during early and active stages of Smyth line vitiligo (Shi and Erf 2012). These reports along with the present study emphasize the role of IFN-γ in the pathogenesis of GV.
Our results suggest that +874A/T intron1 polymorphism of IFNG gene is not associated with GV susceptibility in Gujarat population and our results are in accordance with those of Namian and others (2009). However, the genotype-phenotype correlation for +874A/T polymorphism suggested that IFNG mRNA and protein levels were higher in patients with homozygous TT and heterozygous AT genotypes compared with controls indicating the profound effect of ‘T’ allele with increased levels of IFN-γ in patients. Moreover, the ‘T’ allele frequency was significantly higher in patients with active GV as compared to stable GV indicating the role of +874 ‘T’ allele in disease progression. In addition, active cases of GV showed increased mRNA and protein levels of IFN-γ as compared to stable cases, further confirming the important role of IFN-γ in disease progression. Age of onset analysis with IFNG +874A/T genotypes revealed that patients with TT and AT genotypes had an early onset of the disease as compared to AA wild-type genotype suggesting the involvement of ‘T’ allele in early phase of the disease. Also, patients with an early age of onset of the disease (1–20 years) had significantly higher levels of IFNG transcript as compared to those of late onset groups suggesting the crucial role of IFN-γ in early phase of the disease. Furthermore, a gender-based study indicated that female patients had an early onset of the disease and higher levels of IFNG mRNA and protein levels as compared to males suggesting that females have increased susceptibility towards GV, implicating gender biasness in the development of autoimmunity (Panchanathan and Choubey 2012).
IFN-γ levels have also been monitored in certain autoimmune diseases. Active cases of systemic lupus erythematosus (SLE) and mixed connective tissue disease showed increased levels of IFN-γ suggesting its role in the autoimmune disease pathogenesis (Funauchi and others 1991). Rekha and others (2006) suggested a differential association of high (T) and low (A) producing alleles of IFNG with Hashimoto's thyroiditis and Graves' disease. The high IFNG producing allele T was associated with Hashimoto's thyroiditis where as in Graves' hyperthyroidism the association was stronger with the low producing allele A. In addition, a family based study (Schena and others 2006) with IgA nephropathy showed a strong association between the 13-CA repeat allele and the A variant of the +874A/T single nucleotide polymorphism. The report also confirms a correlation between the +874A allele and lower production of IFN-γ as compared to +874T allele (Schena and others 2006). Furthermore, the T allele of +874A/T polymorphism was found to be associated with elevated IFNG expression in Korean patients with SLE (Kim and others 2010).
There was a significant association between genotype and the frequency of the A allele of the +874A/T polymorphism in atopic patients in Egyptian population (Hussein and others 2009). The study also confirms decreased serum IFN-γ levels along with the presence of homozygous AA genotypes. Further, the 2 North Indian studies reported significant association of IFNG +874A/T polymorphism with increased risk of cervical cancer (Kordi and others 2008; Gangwar and others 2009). Another study on South Indian population suggested a lack of functional association of IFNG +874A/T polymorphism with tuberculosis (Vidyarani and others 2006). These Indian cohort studies for IFNG +874A/T SNP association prompted us to investigate the role of IFNG in Gujarat population with GV.
The IFNG has a (CA)n repeat element within the first intron (Ruiz-Linares 1993). The location of the CA repeat in the first intron is consistent with an effect on either gene transcription or on pre-mRNA processing (Agarwal and others 2000). It has been reported that good IFN-γ producers are considered to be individuals with 12 CA repeats and a ‘T’ at the polymorphic site (Pravica and others 1999, 2000). Individuals with the presence of ‘A’ allele with 13 repeats have low IFN-γ generation potential (Lee and others 2001; Miyake and others 2002). The current study reports for the first time significant association of 12 CA repeats with GV susceptibility. Interestingly, the serum IFN-γ were found to be increased with individuals harboring 12 CA repeats as compared to those of longer repeats suggesting the important role of 12 CA repeats in increased IFN-γ production. However, the CA microsatellite polymorphism has been associated with increased susceptibility to several autoimmune diseases, such as asthma (Nagarkatti and others 2002), type-1 diabetes (Jahromi and others 2000), polymyalgia rheumatica (PMR) (Gonzalez-Gay and others 2004), rheumatoid arthritis (Hanjani-Khani and others 2000) etc.
ICAM1, is important for activating T-cells and recruiting leukocytes (Hedley and others 1998). Beyond its classically described function as an adhesion and viral entry molecule, ICAM1 on the surface of T-cells is thought to participate in signal transduction affecting several T-cell functions, including activation, proliferation, cytotoxicity, and cytokine production and thereby, plays an important role in modulating autoimmune diseases (Stanciu and Djukanovic 1998). Furthermore, IFN-γ and TNF-α induce the expression of ICAM1 on the cell-surface of melanocytes (Yohn and others 1990). ICAM1 protein levels are upregulated in vitiligo skin and in melanocytes from perilesional vitiligo skin (Al Badri 1993). Increased expression of ICAM1 on the melanocytes enhances T cell- melanocyte attachment in the skin and may be involved in the destruction of melanocytes in vitiligo (Al Badri 1993). The present study also showed increased expression of ICAM1 in GV patients suggesting that increased IFN-γ levels might be responsible for increased ICAM1 expression in GV patients. Moreover, the ICAM1 expression was increased in active cases of GV as compared to stable GV suggesting its role in progression of the disease. The ICAM1 expression was increased at early age of onset of the disease, further implicating the important role of ICAM1 in early phase of the disease. Also, female patients showed an increased expression of ICAM1 as compared to male patients suggesting that females have more susceptibility towards GV.
In humans, IFN-γ is implicated in pathology of several autoimmune diseases, including SLE (Lee and others 2001), multiple sclerosis (Panitch and others 1987), allergic encephalomyelitis (Espejo and others 2001), autoimmune nephritis (Heremans and others 1978), and type-1 diabetes (Jahromi and others 2000). IFN-γ- a Th1 cytokine is critical for both innate and adaptive immunity and the importance of IFN-γ in the immune system stems from its immunostimulatory and immunomodulatory effects. In addition, its effect on apoptosis has been suggested as a crucial role in the context of autoimmunity. IFN-γ enhances human B-cell proliferation (Francois and others 1988), activates antigen-presenting cells (APCs) and promotes Th1 differentiation and inhibits Th2 development thereby plays an important role in modulating autoimmune diseases (Oriss and others 1997). IFN-γ promotes specific cytotoxic immunity by indirect mechanisms, such as growth inhibition of Th2 populations and upregulation of antigen processing, presentation, and APC costimulatory molecules, thereby, increasing CD4+ differentiation (Schroder and others 2004). It has been demonstrated that an imbalance between Th1 and Th2 cytokine production is highly correlated with the induction and development of several autoimmune diseases (Kidd 2003). Th1 responses, as characterized by IFN-γ, have been established in vitiligo (Sugita and others 2006). T-cells expanded from peri-lesional vitiligo skin show a predominately Type 1 cytokine profile (ie, IFN-γ and TNF-α) (Wańkowicz-Kalińska and others 2003) and the treatment of vitiligo by using IFN-γ inhibitors has also shown positive therapeutic responses (Skurkovich and Skurkovich 2006); these reports suggest the crucial role of IFN-γ in pathogenesis of GV.
The ultimate pathway of destruction of melanocytes in vitiligo is not known. Apoptosis is one of the cell death pathways suggested for melanocyte destruction. Cytokines, such as IFN-γ, TNF-α, or IL-1 released by lymphocytes and keratinocytes can initiate apoptosis (Huang and others 2002). Recently, we have also shown that increased TNF-α transcript and protein levels correlate with disease progression and higher susceptibility towards vitiligo (Laddha and others 2012). Since, IFN-γ has a role in inducing melanocyte apoptosis and destruction in GV, genetic association studies with IFNG polymorphisms in different ethnic populations need to be explored.
In conclusion, we propose that IFNG CA microsatellite but not +874A/T may be a genetic risk factor for GV in Gujarat population; however, +874T allele may play a role in increased expression of IFNG mRNA and protein levels which could affect the onset and progression of the disease. Moreover, the increased IFN-γ levels in patients can lead to increased ICAM1 expression which is probably an important link between cytokines and T-cells involved in pathogenesis of GV.
Footnotes
Acknowledgments
We thank all vitiligo patients and control subjects for their participation in this study. This work was supported by grants to RB (BMS/Adhoc/122/11-2012) ICMR, New Delhi, India and (GSBTM/MD/PROJECTS/SSA/453/2010-2011) GSBTM, Gandhinagar, Gujarat, India. NCL thanks the Council of Scientific and Industrial Research (New Delhi) for awarding SRF.
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.