
Abstract
The genetic association of CCR5 with human immunodeficiency virus-1 (HIV-1) pathogenesis is well known. The HIV-1 entry into target cells is initiated by the binding of the viral envelope glycoproteins (gp120-gp41) with the cell surface receptor (CD4) and the coreceptor (CCR5), followed by fusion of the viral and cell membranes. Genetic variants of the gene-encoding HIV-1 coreceptor are implicated in the susceptibility to HIV-1 infection. The prevalence of these mutations may vary according to population ethnicity. In the current study, characterization and frequency distribution of the HIV-related gene variants in 135 samples of the Saudi populations were conducted. Polymerase chain reaction (PCR) of 276 bp amplicons was used to rapidly detect Δ32 deletion in the initial sample of DNA. The direct sequence of 2 overlapping PCR amplicons flanking 1059 bp was used to detect single-nucleotide polymorphisms. A single hetrozygous Δ32 deletion allele and 6 single-nucleotide polymorphisms were detected. Only one of the identified haplotypes, Taif-1, which was found in the majority of the tested sample, is identical to CCR5 wild-type alleles. Furthermore, the results of this study raised a concern about the prospective role of the mutations detected among Saudi nationals in the HIV pathogenesis and the clinical use of CCR5 antagonists, which are currently being developed as therapeutics for HIV-1 and inflammatory diseases.
Introduction
C
The structural information of CCR5 among populations can be very useful and essential for providing insights about targeting those receptors. Computational modeling is important for understanding the underlying molecular mechanisms of diseases caused by mutations. The current study aims to screen the potential distribution of the CCR5 gene polymorphisms and to analyze the presence of possible mutations in the CCR5 gene coding region. The study was carried out on healthy Saudi subjects located in the Taif region.
Materials and Methods
Ethical approval
The project and data forms were approved by the Regional Research and Ethics Committee at the Taif University, Saudi Arabia. Written informed consent was obtained from all participants involved in our study.
Study subjects
Peripheral mononuclear cells from 135 Saudi nationals (80 male and 55 females) were collected during the period from September 2011 to March 2012 and used in the current study. Samples were randomly selected from healthy individuals of unrelated families. Name enquiries were carried out to uncover any relationship between subjects before blood collection.
Genomic DNA extraction
Mononuclear cells were collected from an EDTA-collected peripheral whole blood using the Ficoll histopaque 1077 (Sigma). DNA was extracted from the mononuclear cells using an EzWay Genomic DNA Kit (Komabiotech, Inc.) following the manufacturer's instructions.
Polymerase chain reaction
The CCR5-Δ32 genotypes of individual subjects were initially screened using short polymerase chain reaction (PCR) amplicons (Table 1), yielding a 276-bp PCR product for the wild-type CCR5 genotype, while a 244-bp was used for the CCR5-Δ32 mutation (Verma and others 2007). For analysis, 20 μL of each sample was fractionated on a 3% agarose gel with 0.05 μg/mL ethidium bromide. PCR products were directly visualized using UV fluorescence. For the wild genotype (CCR5/CCR5), a single PCR product of 276 bp appeared, while a single product of 244 bp indicated a homozygous mutant (Δ32/Δ32). The presence of both fragments indicated a heterozygous genotype (CCR5/Δ32). The CCR5 polymorphisms of the full-length gene of all individual subjects were screened by PCR using two sets of overlapping primers: CCR5-full-a/CCR5-full-b (Table 1). The two sets of primers, CCR5-full-a/CCR5-full-b, flank 751 and 584 bp amplicons respectively with a 276-bp overlapping sequence.
CCR5 genotyping
PCR amplicons from overlapping primer sets were subjected to electrophoresis in a 1% agarose gel. CCR5-specific bands were excised and purified with the QIAquick gel extraction kit (Qiagen). Each purified PCR product was sequenced directly in both forward and reverse directions (Macrogen). Sequences were trimmed to remove the amplicon primer–linker sequence and assembled. All sequence data used in this study are available in the GenBank database.
Multiple sequence analysis
Comparative analyses were performed using the CLUSTAL W multiple sequence alignment program, Mega 4.1 (Kumar and others 2001). A CCR5 wild-type representative sequence used for the alignments was obtained from the GenBank database. The CCR5 deduced amino acid sequences of the subjects were compared to the wild-type human CCR5 genotype available in the GenBank database.
Building 3D model
The SWISS MODEL service was utilized to build the 3D models for the CCR5 protein of the Saudi CCR5 genes by finding the exact templates. The PDB files were exposed by using the module of the Molsoft Internal Coordinate Mechanics (ICM-Pro) software.
Results and Discussion
The CCR5 genotype was determined by a PCR from the subjects. One hundred thirty four samples were homozygous for the wild-type CCR5 allele, while one was heterozygous for the CCR5-Δ32 allele (Fig. 1). None of the individuals tested was homozygous for the mutation (Δ32/Δ32). Two previous studies described the frequency of Δ32 mutants among Saudi populations: one showed that it possessed a high rate 2.07% (Martinson and others 1997), while the other confirmed the absence of the Δ32 mutation (Stephens and others 1998). The frequency of the CCR5-Δ32 was high in Iranians (2.38%) (Voevodin and others 1998), very low among Syrians (0.7%) and Egyptians (0.6%), and was completely absent from individuals from Lebanon, Sudan, Yemen, the United Arab Emirates, and Egyptians from the Sinai (Salem and Batzer 2007). The very low frequency of the mutant allele CCR5-Δ32 in Arab countries probably derived from the admixture with the populations of European descent rather than as a result of parallel independent mutations (Salem and Batzer 2007). Indeed, no individuals were discovered who were homozygous for any of the CCR5-Δ32.
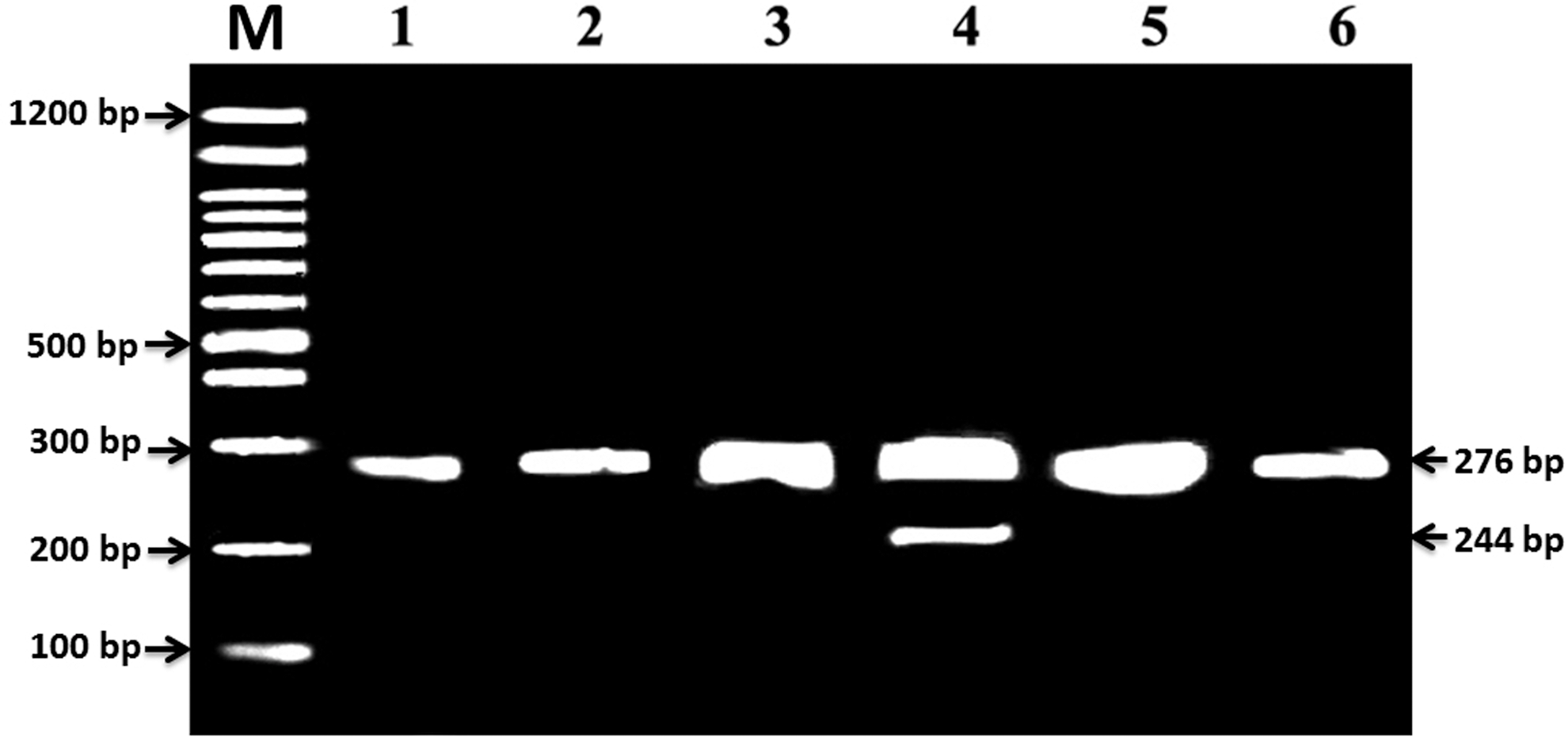
Identification of the CCR5 polymorphism. Lanes 1, 2, 3, 5, and 6 showed 276 (wt), while lane 4 showed hetrozygous Δ32 deletion allele: 276 and 244 bp (Δ32).
It is well known that homozygous CCR5-Δ32 has been shown to confer strong resistance to the HIV-1 infection as well as postponement of AIDS progression among CCR5-/Δ32 heterozygotes (Dean and others 1996; Huang and others 1996; Zimmerman and others 1997). Despite the apparent resistance afforded by the CCR5-Δ32 mutation, protection against HIV-1 infection was not absolute, as evidenced by reports of the HIV-1 infection among Δ32/Δ32 homozygotes (Biti and others 1997; Theodorou and others 1997). The acquisition of the HIV-1 infection in such individuals has been by the parenteral rather than mucosal route, suggesting utilization of the CXCR4 coreceptor by dual-tropic strains of HIV-1 among infected Δ32 homozygotes. The high frequency of CCR5-Δ32 in Europe may indicate that it possesses long-term selective advantage against infectious diseases using CCR5 as an entry coreceptor (Dean and others 1996; de Silva and Stumpf 2004; Galvani and Novembre 2005). Additionally, the high frequency of the CCR5-Δ32 allele in certain populations might affect the severity and the epidemiology of some diseases as well as the mean progression rate of HIV-positive individuals. Conversely, it might also favor the emergence of mutant strains able to bypass the CCR5 defect via using alternative coreceptors (Choe and others 1996). The bubonic plague, which claimed the lives of 25%–33% of Europeans during the Black Death from 1,346 to 1,352 and other large epidemics such as small pox have been used as examples to support this theory (Lenski 1988; Galvani and Slatkin 2003). Accordingly, certain populations have some increase in the survival rate against an HIV epidemic. Conversely, populations with lower frequencies of the CCR5-Δ32 genotype might be expected to possess a higher prevalence or a more rapid disease course. High HIV prevalence in certain areas in Asia and Africa may be attributed to social and demographic factors, as well as differences in the phenotype of circulating viruses (Soto-Ramirez and others 1996).
Mutagenesis studies afforded determinants of the CCR5 coreceptor function. The model of human CCR5 was solved by Palczewski and others (2000) and showed that it consists of a transmembrane (TM) domain of 7 α-helices connected by 3 extracellular loops (ECLs) and 3 intracellular loops (ICLs). The negatively charged amino acids and tyrosine residues in the CCR5 (Asp-11, Tyr-14, Tyr-15, and Glu-18) were found essential for CD4-induced gp120–CCR5 binding and virus entry (Doranz and others 1997; Dragic and others 1998; Farzan and others 1998; Rabut and others 1998) that was found conserved in the Saudi CCR5 genes (Fig. 2) proposing that the Saudi CCR5 genes characterized by a high affinity to the HIV infection. The following amino acid residues in the CCR5 ECLs influenced the coreceptor functions: Gln-93 in ECL1 (Kuhmann and others 1997); Gly-163 in the transmembrane helix 4 (TM4)/ECL2 junction (Siciliano and others 1999); Tyr-184, Ser-185, and Arg-197 in ECL2 (Doranz and others 1997); and Asp-276 and Gln-280 in ECL3 (Doranz and others 1997; Farzan and others 1998). Interestingly, Lys-197 was found in all Saudi CCR5 genes (Fig. 2). On the other hand, a docking study revealed that Glu283 was an important residue in CCR5 antagonism (Kothandan and others 2012). This residue was found conserved in all Saudi CCR5 alleles (Fig. 2). Kothandan and others (2012) studied the 4 CCR5 antagonists (MVC, SCH-C, TAK-779, and Vicriviroc) and close interactions were observed with Tyr89 (TM2), Gly163 (TM4), Thr167 (TM4), Asn252 (TM6), and Leu255 (TM6). Tyr37, Trp86, Tyr108, Ile198, and Glu283 were found important in CCR5 antagonism (Dragic and others 2000; Castonguay and others 2003; Govaerts and others 2003; Tsamis and others 2003; Nishikawa and others 2005; Kondru and others 2008). Interestingly, all these amino acids are conserved in all Saudi CCR5 alleles. The TXP structural motif in TM2 is central for chemokine-induced activation (Govaerts and others 2003), and found conserved in Saudi CCR5 (Fig. 2). This motif was found only in chemokine receptors and a few related peptide receptors. The conformation of the extracellular part of TM2 is governed by the TXP motif and clearly implicated in the activation process, but did not affect chemokine binding (Govaerts and others 2001).

Deduced amino acid sequence of the CCR5. The Saudi CCR5 alleles were compared to the wild-type CCR5. Residues identical to the human wild-type CCR5 molecule are indicated by dots. Transmembrane (TM) domains 1 to 7 are indicated by boxes. Sequences of different CCR5 from the studied subjects have been deposited in GenBank under the accession numbers JX141387-JX141394.
In the current study, 6 strains showed single point mutations at different sites along the CCR5 gene: one of them [Taif.8] showed silent mutations (data not shown), while the others resulted in nonsilent mutations. The Taif 3 sequence showed Pro (P) to Arg (R) 250 amino acid substitution in the TM6. Taif 4 showed Phe (F) to Tyr (Y) 158 amino acid substitution in the TM4. Taif 5 showed Glu (E) to Asp (D) 227 substitution. Taif 6 showed Met (M) to Lys (K) 64 substitution. Taif.7 showed 49 Met (M) to Iso (I) substitution in TM1 (Fig. 2). The polyphen (polymorphism phenotyping) prediction webtool (
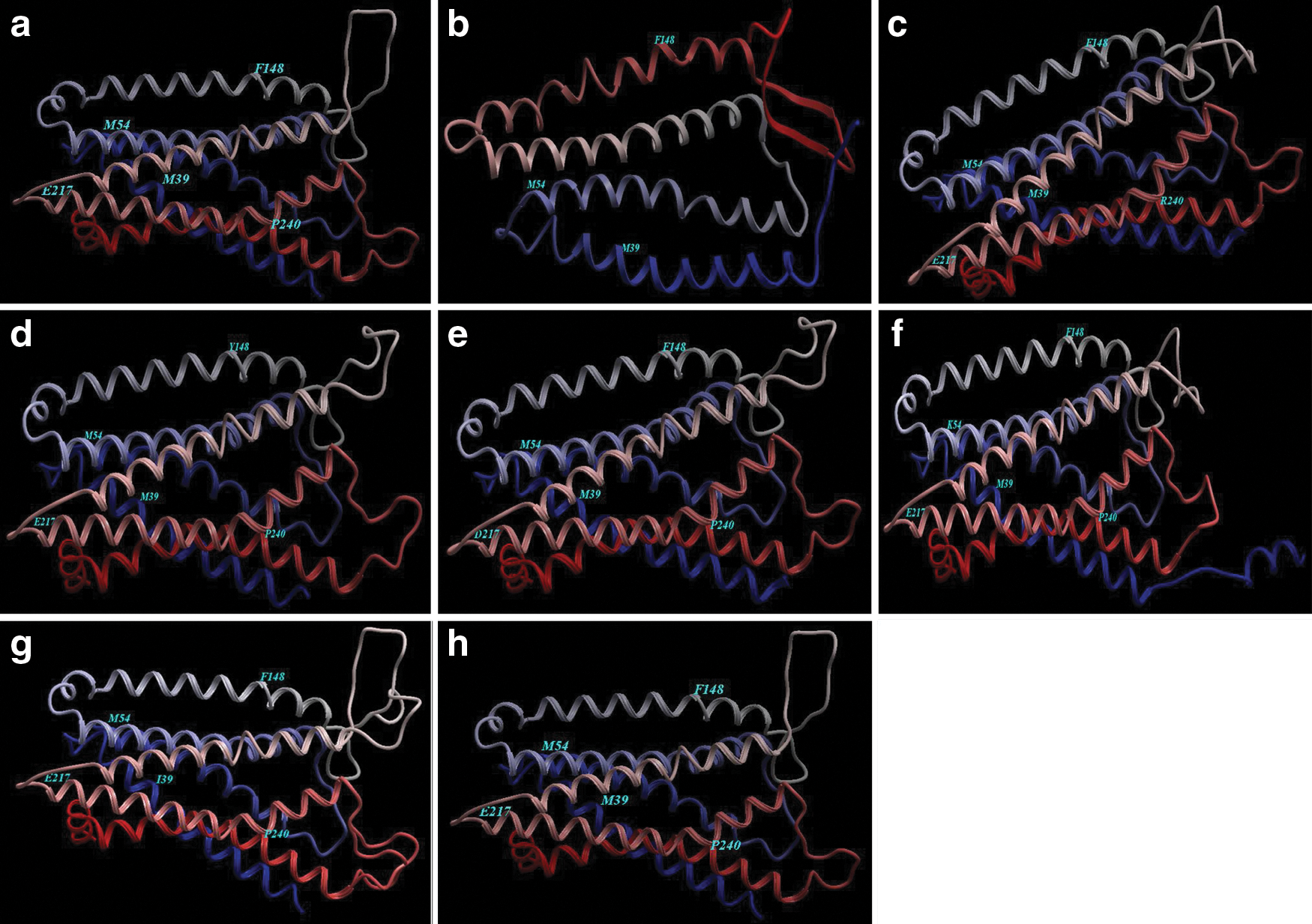
The ribbon diagram of the trimeric structural elements for Saudi CCR5. CCR5 is composed of 3 domains: the extracellular domain, the transmembrane region, and the cytoplasmic domain. It possesses a 7-helix hydrophobic sequence located at the transmembrane region; helix-4, helix-5, and helix-7 cross membrane into the intracellular domain and/or the extracellular domain.
Conservation of an amino acid within a family of proteins can indicate the importance of that amino acid to a common function in that protein family; therefore, it is useful to consider the sites of amino acid variation that are deduced from the novel CCR5 alleles. All the mutations reported here occurred at positions that were conserved throughout the members of the chemokine receptor family. Carrington and others (1997) recorded that high incidence (79%) of nonsynonymous substitutions among human variants is a highly significant elevation and denoted that the results can be interpreted in 2 scenarios: (1) most CCR5 amino acid substitutions do not affect the normal CCR5 function or (2) amino acid substitutions do affect CCR5 function negatively, but confer some other adaptive benefit. The latter explanation is consistent with the CCR5 role in HIV pathogenesis (Dean and others 1996).
In conclusion, the CCR5-Δ32 mutant was found in the Saudi population at a very low frequency. The new variants identified in this study need further studies to speculate the functional aspects of those substitutions and clarify their effects on HIV-1 pathogenicity and progression.
Footnotes
Author Disclosure Statement
The authors declare that they have no competing interests.