
Abstract
Obesity is associated with a low-grade inflammation of white adipose tissue, which can subsequently lead to insulin resistance, impaired glucose tolerance, and even diabetes. MicroRNAs (miRNAs) have been implicated as negative regulators controlling diverse biological processes at the level of post-transcriptional repression. miR-146b is an intergenic miRNA that can regulate the inflammatory process by attenuating cytokine signaling via the nuclear factor-κB pathway. In this study, we focused on investigating the expression of miR-146b in mature human adipocytes and their response to proinflammatory cytokines. Our results showed that miR-146b was highly expressed in the mature adipocytes. The mature human adipocytes responded to proinflammatory cytokines (TNF-α and IL-6) by highly increasing the expression of miR-146b. We cloned and identified a potential promoter of the transcriptional regulation of miR-146b. Interestingly, a fragment about 950-bp length upstream sequences of miR-146b had apparent transcription activity. In addition, the increase in miR-146b promoter activity by TNF-α and IL-6 was also effectively elevated. Our results indicated that a novel role for miR-146b in adipose tissue inflammation and miR-146b may be an important mediator in the process of obesity complications via its own transcription mechanism.
Introduction
O
microRNAs (miRNAs) are an evolutionarily conserved class of small endogenous non-coding RNAs (22 nt) that are involved in post-transcriptional gene repression (Ambros 2004; Bartel 2004). miRNAs have come in the spotlight of research in obesity and metabolic syndrome and found that dysregulation of miRNAs expression is closely associated with these diseases (Kolfschoten and others 2009; Alexander and others 2011; McGregor and Choi 2011). miR-146b is an intergenic miRNA, and distinct genes are encoded on chromosomes10q24. Recently, miR-146b was shown to inhibit both migration and invasion, suggesting that it plays a role in metastasis (Hurst and others 2009). Besides, miR-146b expression level was alteration in inflammatory diseases (Sonkoly and others 2008; Stanczyk and others 2008). However, the possible role of miR-146b in regulating the inflammatory response of mature human adipocytes is not yet known.
Since visceral adipose tissue can produce higher IL-6 amounts than subcutaneous adipose tissues (Fried and others 1998), the expression level of TNF-α was no different between visceral adipose tissue and subcutaneous adipose tissues (Dusserre and others 2000). Furthermore, in visceral adipose tissue, accumulation is associated with increased mortality and risk for diabetes and insulin resistance, whereas subcutaneous adipose tissue generally serves as a storage organ (Despres and Lemieux 2006; Iozzo 2009). We used visceral adipocytes in this study to examine the potential involvement of miR-146b in regulating the inflammatory response of mature human adipocytes. Therefore, we investigated what trend of miR-146b was expressed in adipocytes and how they respond to proinflammatory cytokines TNF-α and IL-6. Here, we show that miR-146b are, indeed, highly expressed in mature human adipocytes than preadipocytes and their expression is highly increased in mature human adipocytes when exposed to proinflammatory cytokines. In addition, the increase in miR-146b promoter activity by TNF-α and IL-6 was also effectively elevated.
Materials and Methods
Cell culture and treatment
Human visceral preadipocytes (HPA-V; ScienCell Research Laboratories, San Diego, CA) were maintained in preadipocyte medium (PAM, Cat. No. 7211; ScienCell Research Laboratories) containing 5% fetal bovine serum, 1% preadipocyte growth supplement, and 1% penicillin/streptomycin solution at 37°C in a humidified atmosphere under 5% CO2. To induce differentiation, serum-free PAM (containing 50 nM insulin, 100 nM dexamethasone, 0.5 mM 3-isobutyl-1-methylxanthine, and 100 μM rosiglitazone) was added to confluent human preadipocytes (Day 0), and the medium was replaced every 2 days for the next 4 days. Thereafter, the medium was replaced with serum-free PAM containing 50 nM insulin and replaced every 2 days until lipid droplets had accumulated in cells (day 15). Fat accumulation was assessed by staining formalin-fixed cells with Oil Red O. Meanwhile, the cells were collected at different time points (Day 0, 15).
Differentiated adipocytes were used for experiments at 15 days after the induction of differentiation, at which point more than 80% of cells showed the morphological and biochemical properties of adipocytes. After overnight incubation in serum-free PAM, human adipocytes were treated with a final concentration of 10 ng/mL TNF-α (Wellen and others 2007) (Sigma, St. Louis, MO) or 30 ng/mL IL-6 (Kralisch and others 2005) (Sigma).
Human embryonic kidney 293T (HEK293T) cells were purchased from the American Type Culture Collection (Manassas, VA) and maintained in Dulbecco's-modified Eagle's medium supplemented with 10% fetal bovine serum. HEK293T cells were also incubated at 37°C under a humidified atmosphere of 5% CO2 in air.
RNA isolation and quantitative real-time reverse transcriptase–polymerase chain reaction
Total RNA from human adipocytes was purified using TRIzol reagent (Invitrogen, Carlsbad, CA) according to the manufacturer's protocol, followed by DNaseI treatment (TaKaRa, Japan). The quality and concentration of the RNA was assessed by Nanodrop2.0 (Thermo Scientific, Waltham, MA). To monitor levels of miRNA, cDNA was synthesized from 200 ng of total RNA using TaqMan miRNA Reverse Transcriptase Kit (ABI, Foster City, CA). Reverse transcriptase–polymerase chain reaction (RT-PCR) was carried out using an Applied Biosystems 7500 Sequence Detection System (ABI 7500 SDS) following the manufacturer's guidelines. Briefly, samples were incubated at 95°C for 10 min for an initial denaturation, followed by 40 PCR cycles consisting of incubation at 95°C for 15 s and then 60°C for 1 min. miRNA expression was normalized to snoNA U6 and miR-103, respectively. Primer identification numbers are 001097 for miR-146b, 000439 for miR-103, and 001973 for snoU6 (ABI).
Plasmids and promoter reporter constructs
We examined the upstream region of miR-146b using UCSC genome browser (
The italic bases with underlines are restriction sites.
Fluorescence and promoter activity
HEK293 cells grew to a density of 60%–70% confluence in 6-well plates and then were transiently transfected with 2 μg reporter plasmids per well with Lipofectamine 2000 (Invitrogen). Meanwhile, empty pTB-Cherry plasmid was used as the control. The upstream fragments could drive Cherry gene to express red fluorescence protein. At 48 h post-transfection, the expression of fluorescence protein in different wells was observed under a fluorescence microscope.
Briefly, 250 ng/well of promoter–firefly luciferase reporter construct and 25 ng/well of Renilla luciferase vector (pRL-TK) were mixed with 0.6 μL of transfection reagent in 20 μL of OptiMem® I Reduced Serum Medium (Life Technologies, Grand Island, NY). The transfection reagent: DNA complex was incubated for 15 min at 25°C, then mixed with 80 μL of complete culture medium, and used to replace culture medium in each well of the 96-well culture dish of HEK293T cells. The transfection was allowed to continue for 24 h by incubating culture dishes at 37°C. The cell culture medium was removed from each well, and the cells were briefly washed with serum-free medium before treating them with indicated combinations of 10 ng/mL TNF-α (Wellen and others 2007) (Sigma) or 30 ng/mL IL-6 (Kralisch and others 2005) (Sigma) in 100 μL serum-free medium for another 24 h at 37°C. The cells were lysed in 50 μL of 1×Passive Lysis Buffer and stored at −20°C until assayed. The luciferase activity was measured using a Dual Luciferase Reporter Assay System (Promega) according to the manufacturer's instructions, and it was expressed as relative luciferase activity by normalizing firefly luciferase activity against Renilla luciferase activity.
Statistical analysis
Representatives of replicate experiments are shown in the figures, and data are shown as means±SEM. Statistical analysis was performed using 1-way ANOVA and a paired Student t-test. A P value less than or equal to 0.05 was considered statistically significant.
Results
miR-146b is increased during maturation of human preadipocytes
In this study, we first investigated the expression level of miR-146b during maturation of human preadipocytes. As shown in Fig. 1, the miR-146b expression level was relatively low in human pre-adipocytes. At 15 days after the induction of differentiation, when more than 80% of pre-adipocytes exhibited typical adipocyte morphology, miR-146b expression was significantly up-regulated using snoU6 and miR-103 as a positive control. Moreover, to certify that the miR-146b expression level was relatively low in human pre-adipocytes, we assayed the expression level of miR-103 by using U6 as an endogenous control, we found that the expression level of miR-103 during maturation of human preadipocytes was at a higher level than miR-146b, and the miR-103 expression level was not altered during preadipocytes differentiation. This observation demonstrated that miR-146b expression was stimulated during the maturation of human preadipocytes into adipocytes.

Expression of miR-146b during the conversion of human preadipocytes (Day 0) into adipocytes (Day 15). Differentiation of human preadipocytes was induced as described in the “Materials and Methods” section. Total RNA was harvested from the humanpreadipocytes on alternate days before (Day 0) and after (Day 15) the switch from growth medium to differentiation medium. miR-146b and miR-103 levels were analyzed by quantitative real-time RT-PCR and normalized to the snoU6, and miR-146b also normalized to the miR-103 levels. The results are presented as the mean±SEM of 3 experiments. P<0.01 compared with the basal levels (Day 0). RT-PCR, reverse transcriptase–polymerase chain reaction.
miR-146b is regulated by IL-6 and TNF-α in human adipocytes
The effect of proinflammatory cytokines (TNF-α and IL-6) on the expression of miR-146b in human adipocytes was investigated further. Mature adipocytes were treated with 30 ng/mL IL-6, and miR-146b expression was examined at different time points (4, 8, and 24 h) and then normalized to snoU6 expression. Noticeable increases in the expression of miR-146b were observed even in both proinflammatory cytokines at 24 h after initiating IL-6 stimulation, with approximately 2.17-fold greater expression than controls (Fig. 2A; P<0.05). Nevertheless, the response of miR-146b was gradually equal to controls at 4 and 8 h. In addition, we observed that miR-146b expression level in human adipocytes treated with 10 ng/mL TNF-α was significantly up-regulated when compared with controls at 4 h (Fig. 2A, P<0.05), and the expression remained high for approximately 24 h (Fig. 2A, P<0.05). Taken together, exposure of the cells to the cytokines TNF-α and IL-6, respectively, resulted in an increase in the expression level of miR-146b (Fig. 2A).

The increase in the expression of miR-146b in human adipocytes by the inflammatory cytokines. The cells were treated with IL-6 (30 ng/mL) and tumor necrosis factor (TNF)-α (10 ng/mL), respectively, and the miR-146b expression was analyzed by real-time PCR and normalized to the snoU6
Meanwhile, to further verify the effect of these factors on miR-146b, miR-103 was also used for normalization in this research, and results were consistent with our previous data using snoU6 for normalization (Fig. 2B).
Identification of promoter regions of miR-146b
miR-146b is an intergenic miRNA; to explore where the promoter region is, we first used the UCSC genome browser to predict their own transcription unit. Then, we cloned the predicted promoter region from more than 2 kb length upstream sequences of miR-146b into pTB-Cherry vector to generate Pro1-pTB-Cherry plasmids. We detected the promoter activity under a fluorescence microscope in transfected HEK293T cells. The results exhibited that Pro1 (chr10:104194201—104196202, 2002 bp) has promoter activity which drives the expression of downstream mCherry gene when compared with the vector alone (Fig. 3A). Then, 3 constructs were made with different lengths of Pro1 and pGL3-Basic: Pro1-pGL3-Basic, Pro2-pGL3-Basic, and Pro3-pGL3-Basic. The experiments demonstrated that Pro2-pGL3-Basic yielded higher luciferase activity when compared with Pro3-pGL3-Basic and equal to Pro1- pGL3-basic (Fig. 3B), suggesting that Pro2 was the major promoter for miR-146b.
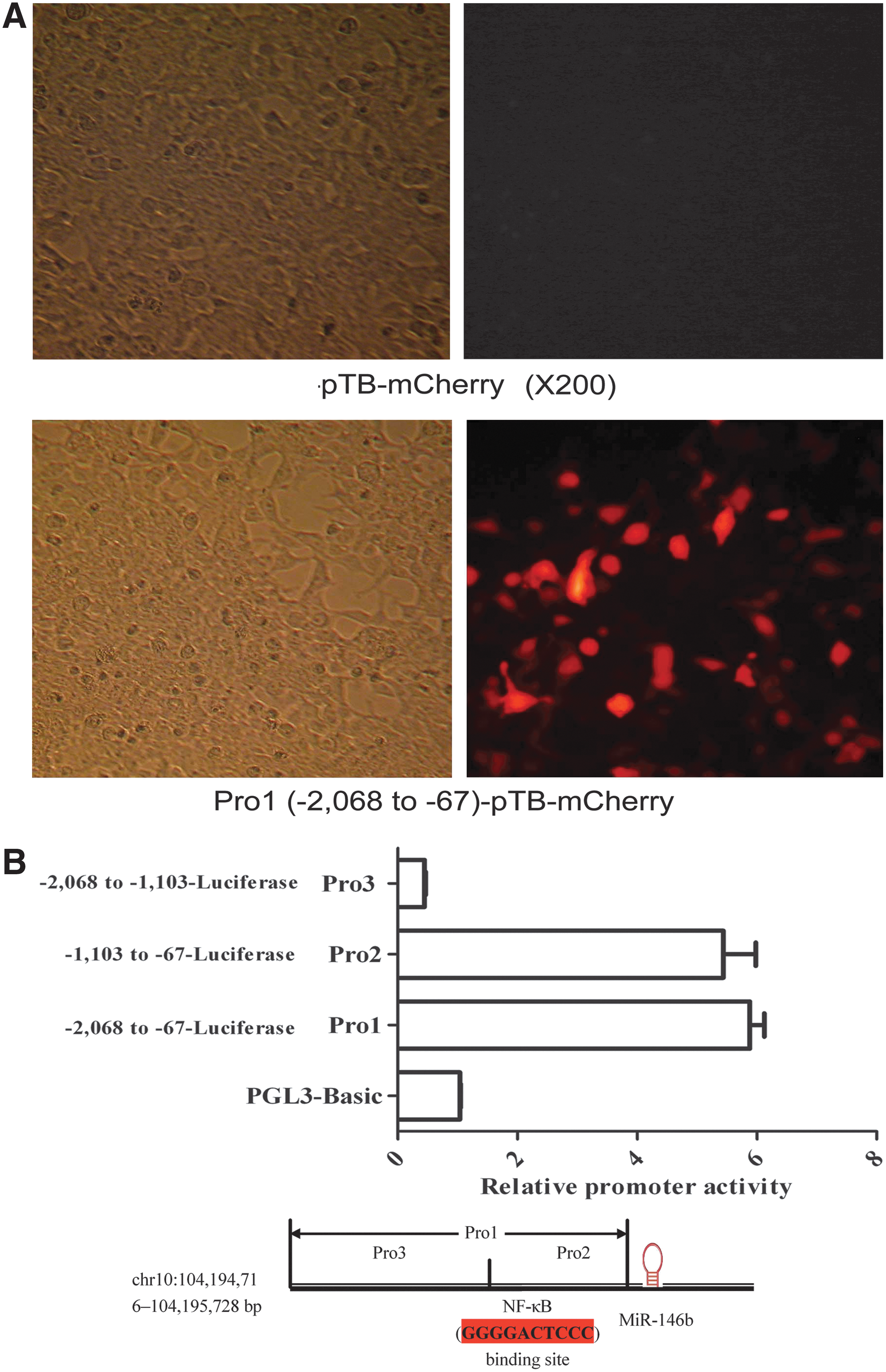
Identification of promoter regions of miR-146b. The activity of miR-146b promoter under a fluorescence microscope. HEK293 cells were transfected with Pro1-pTB-Cherry and the control pTB-Cherry, respectively. The cells were assessed under a fluorescence microscope after 48 h post-transfection. The red fluorescence was observed in the cells transfected with Pro1-pTB-Cherry
Increased promoter activity in major promoter of miR-146b resulting from the proinflammatory cytokine treatment
Furthermore, we employed the HEK-293 to analyze the effect of TNF-α and IL-6 on the major promoter activity of the genes encoding miR-146b. As a consequence, the activity of miR-146b promoter was highly elevated in comparison to the untreated cells, with approximately 4.48-fold (TNF-α) or 3.37-fold (IL-6) greater promoter activity (Fig. 4; **P<0.001), which normalized by pGL3-Basic. Taken together, these results showed that the promoter activity seemed to be up-regulated by these adipokines. Thus, human adipocytes responded to the proinflammatory cytokines treatment by increasing the expression of miR-146b, may be resulting from the effect on the promoter activity.

The activation of miR-146b promoter by proinflammatory cytokines in HEK-293 cells was increased. The cells were transfected with promoter-luciferase constructs of miR-146b (Pro2, the core promoter), and treated with IL-6 (30 ng/mL) and TNF-α (10 ng/mL), respectively. The promoter activity was estimated by measuring the luciferase activity. **P<0.01 when compared with control, n=3. The data shown are representative of 3 similar experiments.
Discussion
Obesity is associated with an insulin resistance and chronic inflammatory response that is characterized by abnormal adipokine production, and the activation of some proinflammatory signaling pathways, resulting in the induction of several biological markers of inflammation and insulin resistance (Hotamisligil and others 1993a; Bastard and others 2002). Recently, data showed that elevation of proinflammatory cytokines augments energy expenditure and decreases the risk for obesity from animal to human (Ye and McGuinness 2013), which indicated that proinflammatory cytokines have benefical effects on obesity. The link between obesity and inflammation has been derived from the reports that proinflammatory cytokines are overexpressed in obesity (Hotamisligil and others 1993a). Over weight and obesity progress to metabolic syndrome though pathophysiological mechanisms at the moment remain unclear. Recently, MiRNAs have been shown to play critical roles in adipocyte differentiation and obesity, insulin resistance, and T2DM (Alexander and others 2011; Ling and others 2012; Yang and others 2012). In addition, some miRNAs regulated by adiponectin were identified as novel targets for controlling adipose tissue inflammation (Ge and others 2012). Here, we demonstrate that a novel role for miR-146b in adipose tissue inflammation and miR-146b may be an important mediator in the process of obesity complications via its own transcriptional mechanism.
miRNAs, which are short non-coding RNAs that are involved in post-transcriptional regulation of gene expression, participate in a wide range of biological functions and play an important role in various diseases. Accumulating data suggest that miR-146b is implicated in a variety of cellular functions, including obesity, inflammatory, infection, oncogenesis, and immune response. For example, miRNA-146b was highly expressed in adipose tissue after long-term high-fat diet-induced obesity mice (Chartoumpekis and others 2012); miR-146a/b as negative regulators of inflammatory process results from their ability to repress the target gene translation, interleukin-1-receptor-associated kinase-1 (IRAK1), and TNF receptor-associated factor 6 (TRAF6), known modulators of nuclear factor-kappaB (NF-κB) pathway, which participate in innate immune response (Taganov and others 2006; Bhaumik and others 2009). Interestingly, human preadipocytes constitute one major model that is used for understanding the development of obesity and obesity-related inflammatory responses, and miR-146b was also shown to be differentially expressed during preadipocyte differentiation in this study. Thus, it can be hypothesized that miR-146b is a mediator of obesity-related inflammatory responses.
It is well known that IL-6 production by adipose tissue is enhanced in obesity (Bastard and others 2002). A previous study reported that TNF-α resulted in the inhibition of 3T3-L1 adipocyte differentiation by up-regulating miR-155 expression (Liu and others 2011). In the study by Kutty and others (2013), they found that miR-146a and miR-146b are expressed in human RPE cells in culture and their expression is highly induced by proinflammatory cytokines (IFN-γ+TNF-α+IL-1β). In our results, a significant increase in miR-146b expression was observed at the TNF-α and IL-6 concentration of 10 and 30 ng/mL, respectively, which is consistent with other reseachers' study. These inflammatory cytokines have obvious effects on miR-146b expression when we used snoU6 as an endogenous control. Moreover, miR-103 is considered the most stable miRNA transcript across all biological backgrounds in human adipose tissues of different origins (Neville and others 2011). Therefore, miR-103 was also used for normalization in this research and we got similar results. Furthermore, the expression of miR-146a and miR-146b-5p is greatly increased in human monocytes by lipopolysaccharide, TNF-α, and IL-1β (Taganov and others 2006). Thus, in combination with previous studies, our data indicate that miR-146b may be involved in obesity-related inflammatory responses. However, the possible mechanism of proinflammatory cytokines in regulating miR-146b exprssion is not yet known.
Recently, many high-throughput methods have been applied to study the transcriptional regulation and promoters of miRNA genes in mammals. The promoter of a gene is a crucial control region for its transcription initiation (Smale and Kadonaga 2003). To understand the mechanism and conditions of the activation of miRNA genes, it is required to locate their core promoter regions. According to miRNA locations in the genome, miRNA genes are classified into intragenic and intergenic ones. For intergenic miRNAs, their primary transcripts are transcribed from individual, non-protein-coding genes and have their own promoters (Weis and Reinberg 1992). miR-146b is an intergenic miRNA; therefore, we identified potential promoters of miR-146b by promoter prediction method to predict their core promoters and then verify the predictions by promoter activity assay. Our results showed that upstream (−1,103 to −67 bp) of miR-146b was its promoter and there would be some enhancers in upstream (−1,103 to −67 bp). More interestingly, the promoter activity was up-regulated by these adipokines in HEK293 cells. The region of −1,103 to −67 bp is the core promoter region that contains essential components (NF-κB) for the regulation of gene transcription, analyzed by TFSEARCH (
The results of this study show that miR-146b expression was markedly increased by inflammatory cytokines (TNF-α and IL-6). Up-regulation of miR-146b expression may be involved in TNF-α and IL-6-induced inflammatory responses, and miR-146b or its promoter may be a novel target and a mediator for regulation adipose tissue inflammation and obesity-related inflammatory response. However, the precise mechanism of obesity-related inflammatory response remains to be fully elucidated. Therefore, further investigations are required to clarify this issue.
Footnotes
Acknowledgments
This study was supported by the grants from the National Key Basic Research Program of China (2013CB530604), the National Natural Science Foundation of China (81100619, 81100618), Program for Innovative Research Teams of Jiangsu Province (LJ201108), Nanjing Technological Development Program (201104013), and the Postgraduate Program for Innovative Research of Jiangsu Province Higher Education Institutions (CXLX12-0559).
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.