
Abstract
Interleukin (IL)-4 promotes the regression of granulomas during the late phase of Rhodococcus aurantiacus infection. In this study, the contribution of IL-4 to the initial response against this bacterium was investigated using IL-4-deficient mice. Compared with wild-type (WT) mice, IL-4-deficient mice displayed remarkably lower tumor necrosis factor (TNF)-α and IL-6 secretion in the liver, spleen, and blood at the initial phase of infection, along with improved survival. IL-4-deficient mice also showed diminished IL-10 secretion in the spleen and blood; however, hepatic IL-10 levels were similar to those observed in WT animals, and were concomitant with augmented interferon (IFN)-γ production and decreased bacterial burden in the liver at the early infection phase. Histological examination revealed reduced hepatic granuloma formation in infected IL-4-deficient mice. On challenge with heat-killed R. aurantiacus, IL-4-deficient mouse macrophages showed reduced expression of TNF-α, IL-6, and IL-10 at both the gene and protein levels compared with WT mouse cells. These findings indicate that during the initiation of R. aurantiacus-induced inflammation, IL-4 deficiency attenuates cytokine responses in macrophages, which contributes to amelioration in mouse survival and reduction of granulomatous inflammation, and augments a hepatic IFN-γ response which transiently accelerates bacterial elimination.
Introduction
T
Rhodococcus aurantiacus is an intracellular, psychrophilic, gram-positive bacterium that is closely related to members of the genera Corynebacterium, Mycobacterium, and Nocardia (Tsukamura and Kawakami 1982). Inoculation of this bacterium to mice induces a Th1-type granulomatous inflammatory response, which resembles human sarcoidosis, and the high level of IL-4 induced during the late phase of infection contributes to the regression of granulomas (Asano and others 1996). In this experimental model, an early increase in IL-4 mRNA expression was also observed in the organs (Asano and others 1996); however, the complexity of IL-4 makes it difficult to accurately predict the role of this cytokine in the innate inflammatory response.
The aim of this study was to assess the influence of IL-4 on the initial host response on R. aurantiacus exposure by using experimental models of intravenous infection in IL-4-deficient mice. We found that IL-4 deficiency induced a significant reduction in pro- and anti-inflammatory cytokines in macrophages and an increased hepatic IFN-γ response during the initial stage of infection, which contributed to improvement in mouse survival, attenuation of granulomatous inflammation, and a transient increase in bacterial elimination. This work may provide novel insights into the function of IL-4 in innate immune-inflammatory responses and is expected to facilitate the development of immunotherapies for infectious diseases, including human sarcoidosis.
Materials and Methods
Mice
C57BL/6 wild-type (WT) mice and IL-4-deficient mice were purchased from Jackson Laboratory (Bar Harbor, ME). After 7 days of acclimation, 7- to 8-week-old female mice were used for in vivo infection experiments, and the males were used for in vitro experiments. All the mice were housed in a temperature-controlled room with a 12-h light/dark diurnal cycle and had free access to food and water throughout the experiment. Experimental procedures were approved by the Institutional Animal Care and Use Committee of Hokkaido University (approval number 13-0044) and were performed according to the Guidelines for Animal Experiments at the Graduate School of Medicine, Hokkaido University.
Experimental infections with R. aurantiacus
Female mice were inoculated via the lateral tail vein with 1×108 colony-forming units (CFU) of the viable R. aurantiacus strain 80005 suspended in 0.2 mL of phosphate-buffered saline (PBS). Mice injected with 0.2 mL of PBS alone served as controls. Mouse survival was monitored daily till day 14 postinfection, and survival curves were plotted.
Bacterial counts in organs
The livers and spleens of infected mice were harvested at the indicated times postinfection and homogenized in RPMI 1640 medium (Sigma, St. Louis, MO) at a concentration of 0.1 g/10 mL, and 100 μL of the organ homogenates and their serial 10-fold dilutions were plated on nutrient agar plates. Viable colonies were counted at 48 h after culture. Using this method, we could detect >103 bacteria/g of organ tissue.
Sample preparation for cytokine assay
The livers and spleens were aseptically removed from mice and suspended in RPMI 1640 medium containing 1% (w/v) CHAPS (Wako Pure Chemical Co., Kyoto, Japan). Homogenates (10% w/v) were prepared with a Dounce grinder. The homogenates were cleared by centrifugation at 2,000 g for 30 min at 4°C, and the supernatants were stored at −80°C until use in the cytokine assay. Blood samples were also collected by venipuncture into 1-mL sterile syringes containing heparin and were separated by centrifugation at 1,200 g for 20 min at 4°C. Separated plasma samples were aliquoted and stored at −80°C for subsequent tests.
Isolation and stimulation of peritoneal macrophages
Peritoneal macrophages were harvested from uninfected male mice and suspended in 0.83% ammonium chloride solution containing 10% (v/v) Tris buffer (pH 7.65) to lyse erythrocytes. The cells were resuspended in RPMI 1640 medium supplemented with 10% fetal bovine serum, 100 IU/mL penicillin, and 100 mg/mL streptomycin, and then plated in 24-well plates at 5×105 cells/well for TNF-α and IL-6 enzyme-linked immunosorbent assay (ELISA), and in 35-mm dishes at 2×106 cells/dish for IL-10 ELISA and total RNA extraction.
After culture for 2 h at 37°C, floating cells were removed, and the attached macrophages were further cultured for 12 h and then stimulated with heat-killed R. aurantiacus (multiplicity of infection of 2).
Quantification of cytokine production
ELISAs were used to determine cytokine concentrations from organ extracts, plasma, and cell culture supernatants. The TNF-α level was determined as previously described (Yimin and Kohanawa 2006). A purified hamster anti-TNF-α monoclonal antibody (mAb) and a purified rabbit anti-TNF-α polyclonal Ab (both from Endogen, Woburn, MA) were used as the capture and detection Abs, respectively. A standard curve was constructed for each experiment by serially diluting recombinant TNF-α (R&D Systems, Minneapolis, MN). IL-6 concentration was also measured using ELISA. Plates were coated with purified rat anti-IL-6 mAb (BD Bioscience, Inc., San Jose, CA) and incubated with culture supernatants or organ extracts. IL-6 was detected with a biotinylated rat anti-IL-6 mAb (BD Bioscience, Inc.). All ELISAs were run with recombinant IL-6 (BD Bioscience, Inc.). IL-4, IL-10, and IFN-γ concentrations were measured using the DuoSet ELISA Development kits (R&D Systems) according to the manufacturer's protocol.
Quantitative real-time polymerase chain reaction analysis
Total RNA was isolated from the spleens, livers, and peritoneal macrophages of mice using TRIzol reagent (Invitrogen, Carlsbad, CA) and was reverse transcribed to first-strand cDNA using the High-capacity cDNA Reverse Transcription kit (Applied Biosystems, Foster City, CA). The cDNA was utilized for quantitative real-time polymerase chain reaction (PCR) with the Power 2×SYBR Green PCR Master Mix and monitored on an ABI Prism 7000 sequence detection system (both from Applied Biosystems). The PCR conditions were as follows: 95°C for 10 min, followed by 40 cycles at 95°C for 15 s and 60°C for 1 min. The primer sequences of the selected genes are shown in Table 1. The relative expression levels of the target genes compared with those of the untreated control samples were normalized to the expression of the housekeeping gene GAPDH.
Histopathological examination
Animals were sacrificed, and the liver samples that were removed were fixed in 10% neutral formalin solution and embedded in paraffin blocks for Gram staining, immunostaining of F4/80 (a surface marker of mouse macrophages) and CD3, and hematoxylin and eosin (H&E) staining. Tissue sections (2 μm in thickness) were immunostained with rabbit anti-mouse F4/80 (Acris Antibodies, Inc., San Diego, CA) or rabbit anti-human CD3 (DakoCytomation, Kyoto, Japan), followed by detection using the STAT-Q rapid IHC system (Innovex Biosciences, Richmond, CA). As a negative control, slides were incubated with normal rabbit IgG (Santa Cruz Biotechnology Inc., Santa Cruz, CA). Tissue sections were also stained with H&E using standard techniques. The granuloma area was quantified using the ImageJ software, and data were expressed as the percentage of field area.
Statistical analysis
Significant differences between the values in the mouse groups were calculated using the Student's t-test. The log-rank test was used for data analyses of Kaplan–Meier survival curves. P values of <0.05 were considered statistically significant.
Results
IL-4 expression during the early phase of infection in WT mice
We first investigated endogenous IL-4 expression in the organs and blood of WT mice during the early phase of R. aurantiacus infection. IL-4 was released in the liver with a maximum level at day 1 postinfection; however, no IL-4 secretion was detected in either the spleen or blood (Fig. 1A). In agreement with hepatic IL-4 production, WT mice showed increased IL-4 mRNA expression at day 1 postinfection in the liver (Fig. 1B).

Endogenous IL-4 expression in WT mice during the initial infection. After intravenous infection with 1×108 CFU of Rhodococcus aurantiacus, the protein secretion
Survival of infected mice
After intravenous inoculation with 1×108 CFU of viable R. aurantiacus, the survival of WT and IL-4-deficient mice was monitored till 14 days. As shown in Fig. 2, WT mice displayed a 60% survival rate at day 5 postinfection; thereafter, they survived the infection until day 14 postinfection. Compared with their WT counterparts, IL-4-deficient mice showed a higher survival rate during infection (87% at day 14 postinfection; Fig. 2). PBS injection had no effect on the survival of mice (data not shown).
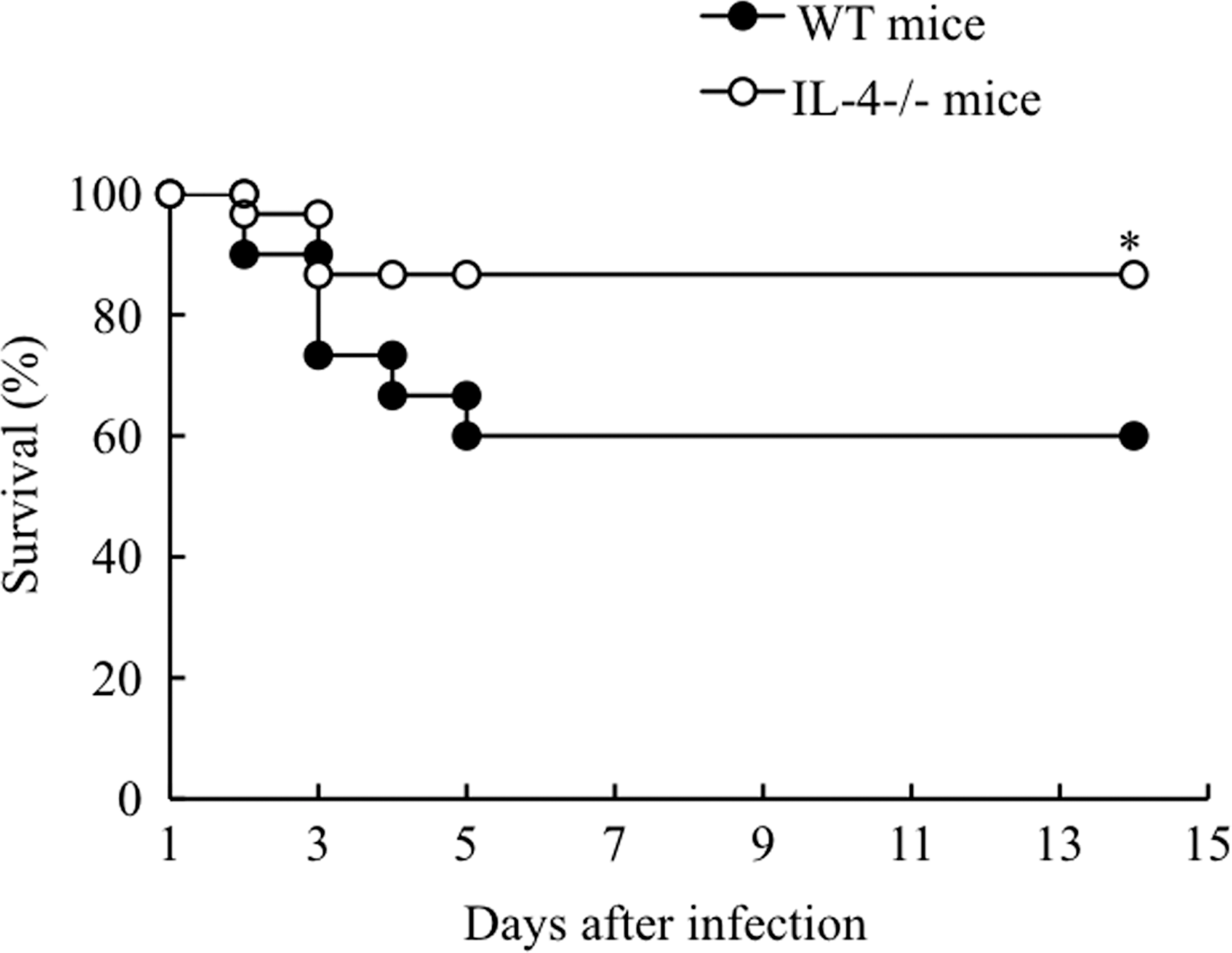
Survival rates of infected mice. After intravenous infection with 1×108 CFU of R. aurantiacus, the survival rates of WT mice and IL-4-deficient mice were determined over a 14-day experimental period. Ten mice were used in each group per experiment, and survival curves were generated from the cumulative results of 3 independent experiments. *P<0.05 versus infected WT mice.
Bacterial loads and elimination in mouse organs
In numerous infectious diseases, the host is often unable to control bacterial replication and quickly succumbs to infections; therefore, we investigated whether the improvement in survival rates observed in IL-4-deficient mice might be associated with more effective removal of R. aurantiacus. After inoculation, bacteria failed to proliferate in the organs of the WT and IL-4-deficient mouse groups (Fig. 3A, B). IL-4-deficient mice exhibited a modest, but statistically significant, reduction in bacterial counts in liver tissues within 3 days postinfection relative to WT mice (11% decrease at day 3 postinfection); thereafter, the same degree of reduction in hepatic bacteria counts was found in both mouse groups, and bacteria were undetectable at day 11 postinfection (Fig. 3B). Observation of Gram-stained liver sections of mice at day 3 postinfection also revealed that IL-4-deficient mice had fewer clusters of the bacteria at infectious foci than did WT mice (Fig. 3C). Conversely, there was no significant difference between the 2 mouse groups in the splenic bacteria load, which decreased gradually until day 11 postinfection (Fig. 3A).

Kinetics of bacterial load in mouse organs. After infection with 1×108 CFU of R. aurantiacus, the numbers of viable bacteria present in the spleens
Endogenous cytokine secretion in mice
To determine whether IL-4 deficiency influences other cytokine responses during the initiation of infection, the concentrations of IFN-γ, TNF-α, IL-6, and IL-10, which are involved in the development of R. aurantiacus-induced inflammation (Asano and others 1996; Yimin and Kohanawa 2006; Yimin and others 2008), were measured in WT and IL-4-deficient mice. As depicted in Fig. 4A and B, and 5A–C, there was no cytokine production detected in the spleens and livers of uninfected (PBS-treated) WT and IL-4-deficient mice. Inoculation with viable bacteria into either WT mice or IL-4-deficient mice induced a rapid elevation of IFN-γ in the spleen at day 1 postinfection; thereafter, the splenic IFN-γ level continuously fell until at least day 7 postinfection (Fig. 4A). There was no statistically significant difference in splenic IFN-γ secretion levels between the 2 mouse groups within 3 days postinfection; however, from day 5 postinfection, IFN-γ production was markedly lower in the IL-4-deficient group (87% decrease at day 7 postinfection vs. WT mice; Fig. 4A). The WT animals also showed a biphasic pattern of IFN-γ release in the liver, with an early peak at day 1 postinfection and a subsequent peak at day 7 postinfection (Fig. 4B). Although IL-4-deficient mice exhibited a similar secretion pattern of hepatic IFN-γ as that seen in WT mice, the early peak level of IFN-γ was further increased at day 1 postinfection (1.83-fold increase vs. WT mice), and the latter peak was markedly lower in the IL-4-deficient mice (48% decrease vs. WT mice; Fig. 4B). These findings demonstrate the distinct effects of IL-4 absence on IFN-γ secretion at different infection phases and in different organs.
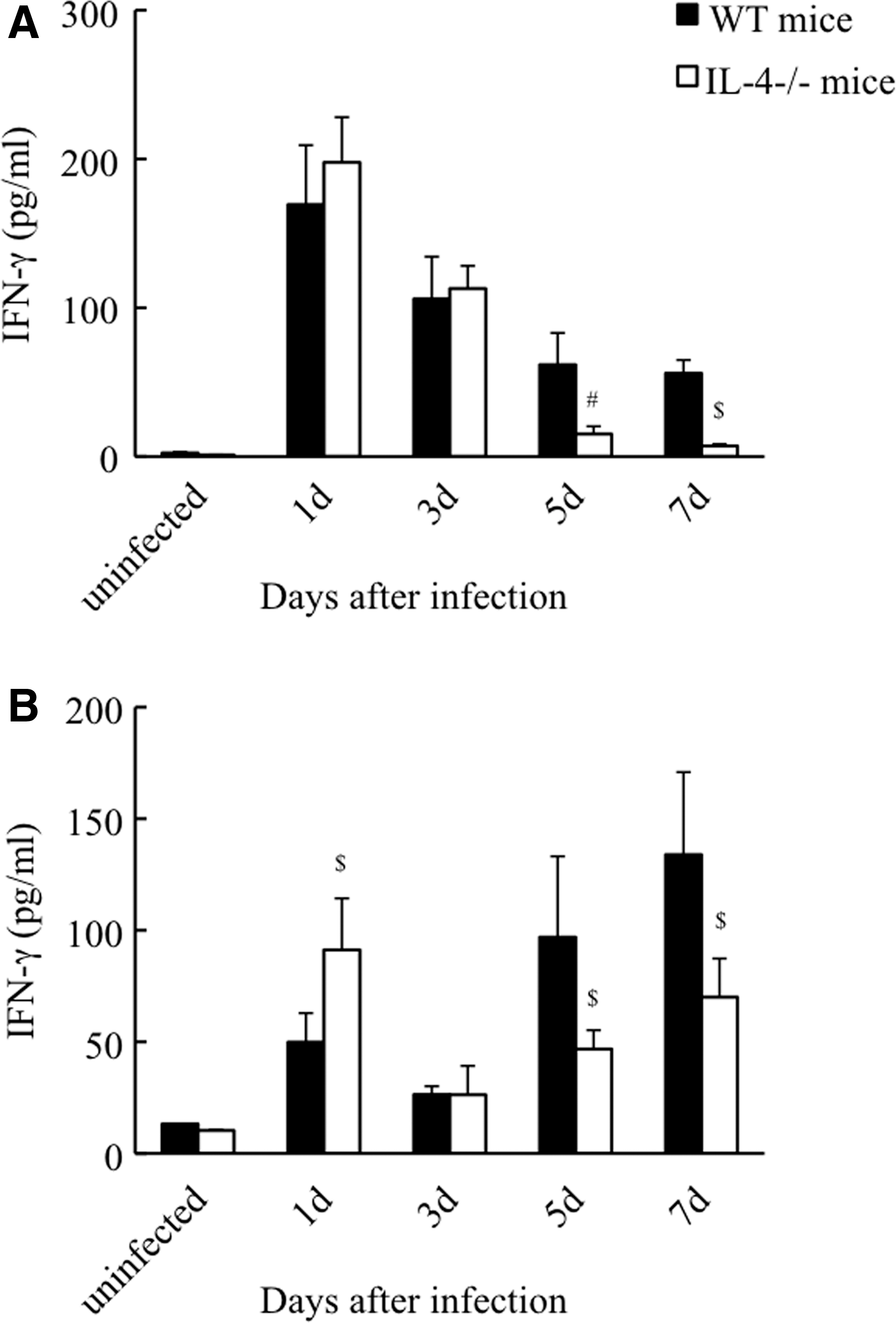
Kinetics of IFN-γ production in the spleens
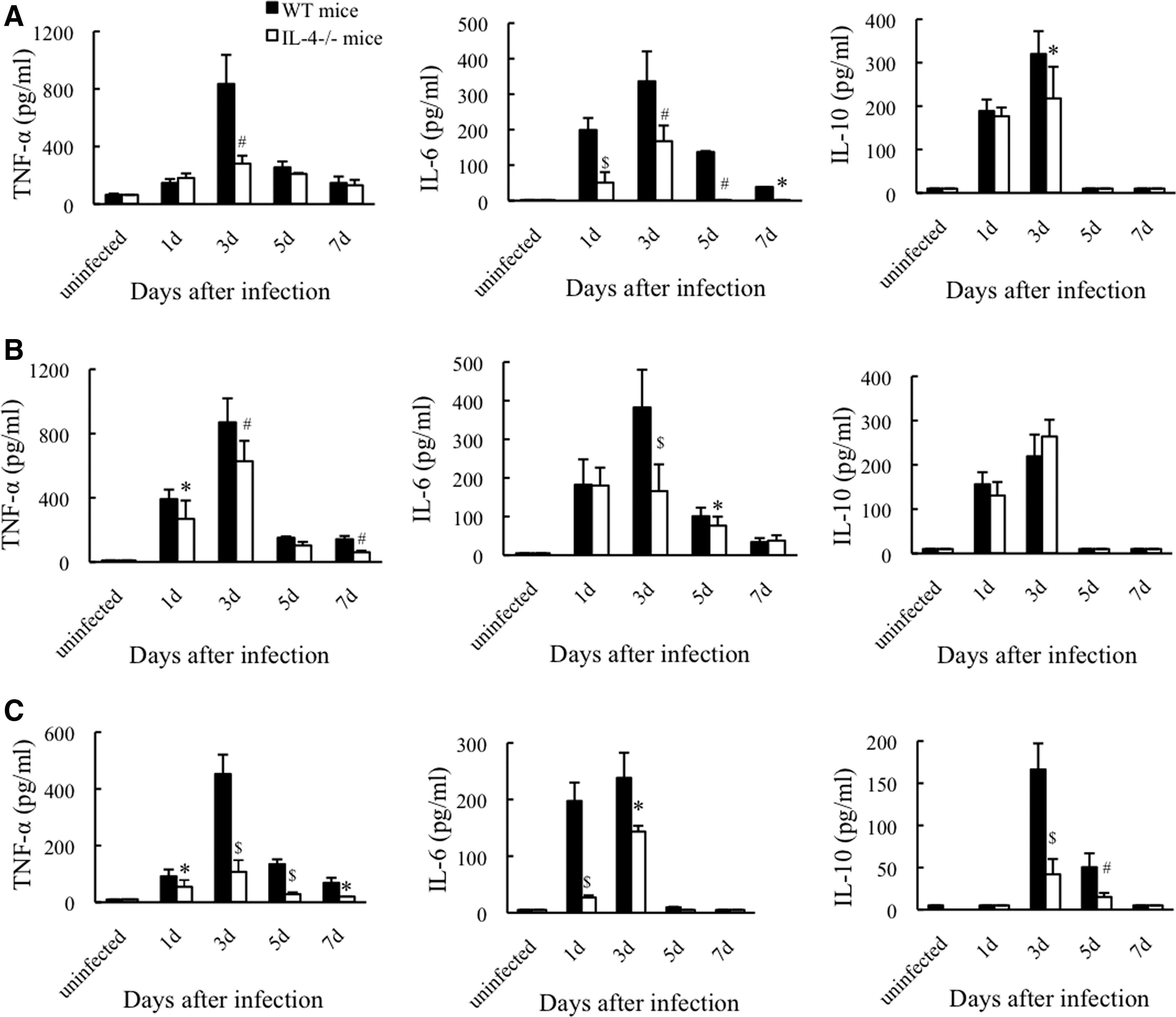
Kinetics of cytokine production in the organs and blood of mice. After inoculation with R. aurantiacus or PBS, the concentrations of TNF-α, IL-6, and IL-10 in spleen extracts
We further assessed the production of TNF-α, IL-6, and IL-10 during the early phase of infection. Although inoculation with R. aurantiacus led to similar secretion kinetics of TNF-α and IL-6 in the spleens, livers, and blood of both WT and IL-4-deficient mice, all of which peaked at day 3 postinfection, the levels of TNF-α and IL-6 were markedly impaired in the latter, which exhibited 66%, 28%, and 76% reductions in TNF-α peak levels, and 50%, 57%, and 39% decreases in IL-6 peak levels in the spleen, liver, and blood, respectively (vs. WT mice; Fig. 5A–C). After infection, IL-10 was also released in the spleen, liver, and blood of both WT and IL-4-deficient mice, which peaked at day 3 postinfection (Fig. 5A–C). However, IL-10 release in IL-4-deficient mice was significantly attenuated by 32% in the spleen and by 75% in the blood at day 3 postinfection compared with that in WT mice, while there was no difference in hepatic IL-10 secretion between the 2 mouse groups (Fig. 5A–C). These data suggest that IL-4 deficiency attenuates not only pro-inflammatory but also anti-inflammatory cytokine responses during the initiation of systemic R. aurantiacus infection.
In vitro cytokine expression in macrophages
Since the production of TNF-α, IL-6, and IL-10 is mainly induced by macrophages during the early phase of R. aurantiacus infection (Yimin and Kohanawa 2006), we further investigated the influence of IL-4 deficiency on the cytokine response in macrophages. Macrophages from WT and IL-4-deficient mice did not produce any cytokines under the no-stimulation condition (Fig. 6A). After stimulation with heat-killed R. aurantiacus, WT mouse macrophages showed enhanced TNF-α production that reached a maximum at 24 h thereafter, the TNF-α level decreased until at least 48 h poststimulation (Fig. 6A). These cells also displayed continuous increases in both IL-6 and IL-10 secretion in a time-dependent manner over a 48-h experimental period (Fig. 6A). In contrast, reduced TNF-α secretion was detected in IL-4-deficient mouse macrophages with a 77% decrease in peak production at 24 h poststimulation compared with the peak level in WT mouse macrophages (Fig. 6A). IL-4 deficiency also diminished macrophage secretion of IL-6 throughout the experimental period, which fell to 44% at 48 h poststimulation compared with levels observed in WT mouse macrophages; moreover, IL-10 expression was hardly detected in IL-4-deficient mouse cells (Fig. 6A). These results suggest that R. aurantiacus-induced cytokine production in macrophages is heavily downregulated in the absence of IL-4.
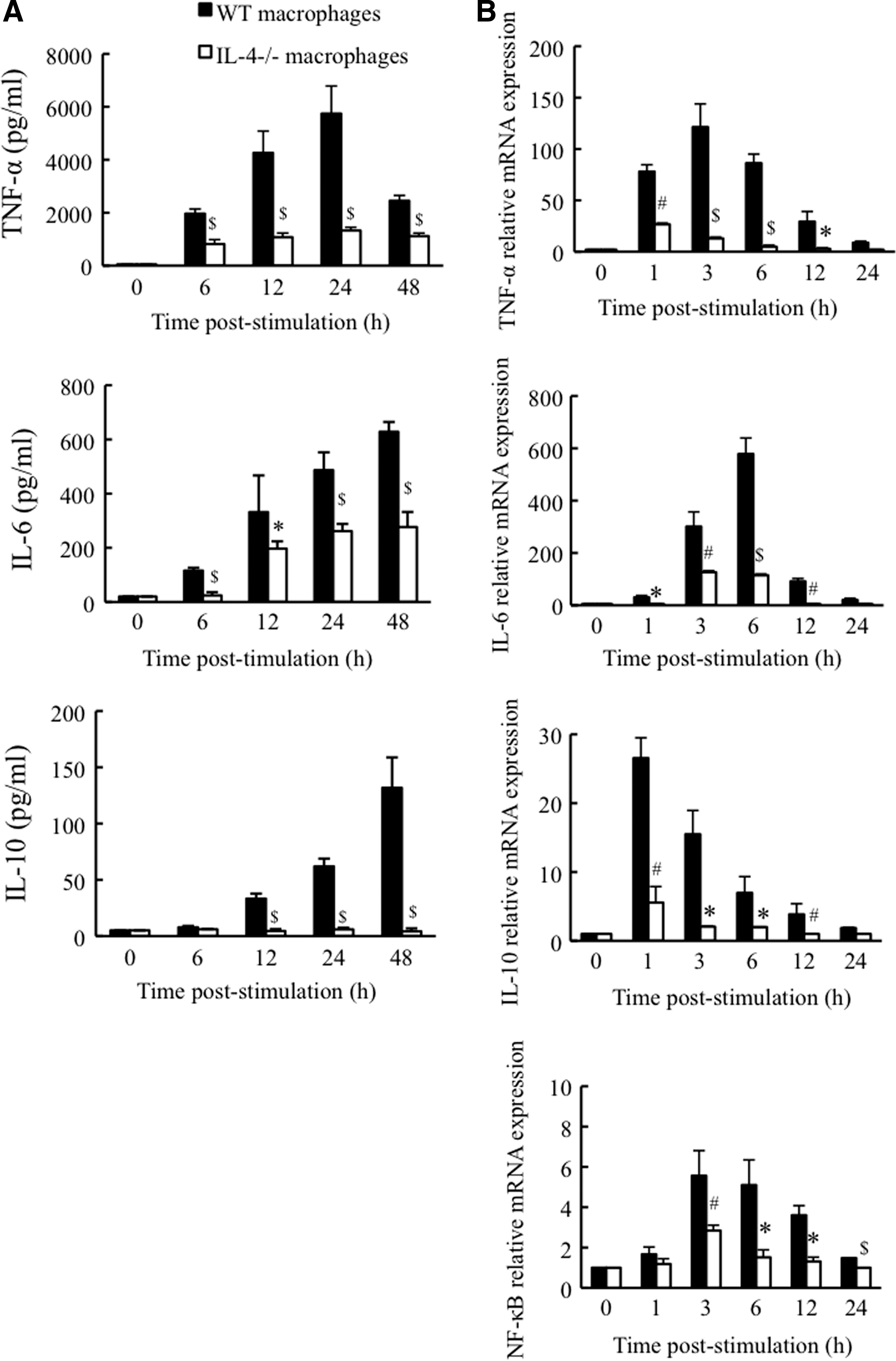
Kinetics of cytokine and NF-κB expression in mouse macrophages. Macrophages from WT and IL-4-deficient mice were unstimulated or stimulated with heat-killed R. aurantiacus (multiplicity of infection of 2), and then, the protein expression of TNF-α, IL-6, and IL-10
To determine whether the secretion of cytokines by macrophages is consistent with their gene expression, we evaluated cytokine mRNA levels using real-time PCR. As shown in Fig. 6B, heat-killed R. aurantiacus induced enhanced expression of TNF-α, IL-6, and IL-10 mRNA in WT mouse macrophages, which peaked at ∼3, 6, and 1 h poststimulation, respectively. In contrast, IL-4-deficient mouse cells showed significantly low expression of TNF-α, IL-6, and IL-10 mRNA after stimulation; the peak mRNA levels of TNF-α at 1 h poststimulation, IL-6 at 3 h poststimulation, and IL-10 at 1 h poststimulation decreased by 78%, 78%, and 79%, respectively, compared with their peak levels in WT mouse cells (Fig. 6B). Thus, these gene expression patterns were correlated with the levels of protein secretion. Moreover, we also determined the gene expression of nuclear transcription factor (NF)-κB, an important transcription factor involved in inflammatory cytokine production, in macrophages. After exposure to heat-killed R. aurantiacus, the expression of NF-κB mRNA was augmented and peaked at 3 h in WT mouse macrophages; however, the upregulation in NF-κB mRNA expression was significantly suppressed in IL-4-deficient mouse cells during the stimulation period, in which the peak decreased by 49% at 3 h compared with that in WT mouse cells (Fig. 6B). Treatment with heat-killed R. aurantiacus did not affect GAPDH mRNA levels in macrophages from either WT or IL-4-deficient mice.
Hepatic histopathology of mice
We further compared the development of hepatic granulomas between WT and IL-4-deficient mice by histological examination at day 14 postinfection, which is a sufficient time for granuloma formation (Yimin and Kohanawa 2006). No granuloma was observed in the livers of uninfected WT and IL-4-deficient mice, whereas inoculation with R. aurantiacus induced the development of a large number of non-necrotic granulomas in the livers of WT mice, which were mainly composed of macrophages and T lymphocytes (mean area of granulomas per field, 21%; Fig. 7A–D). However, IL-4-deficient mice showed a significant decrease in the area of granulomas in the liver (mean area of granulomas per field, 7%; Fig. 7A–D). These data indicate that the development of R. aurantiacus-induced granulomas is dependent on the presence of IL-4.
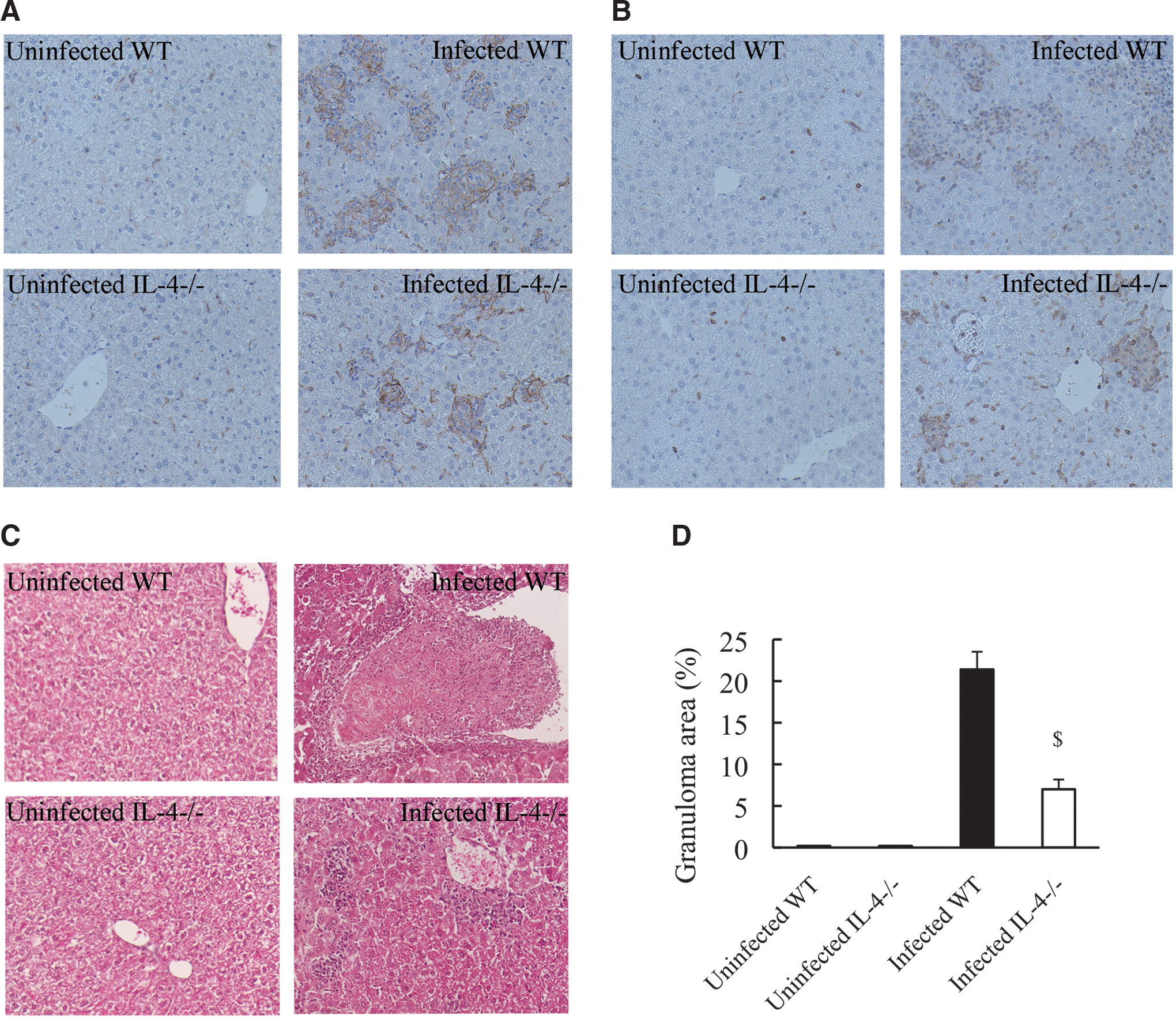
Histological analysis of mouse livers at 14 days postinfection. WT mice and IL-4-deficient mice treated with R. aurantiacus or PBS were sacrificed at day 14, and representative photomicrographs (original magnification, ×200) of liver sections stained with an F4/80 antibody
Discussion
An excessive pro-inflammatory cytokine response to invading pathogens results in exacerbated host immune responses, septic shock, and even death (Murapa and others 2011; Stegmayr and others 2012). Our previous studies also indicated that the overproduction of TNF-α and IL-6 during the initial phase of R. aurantiacus infection is responsible for mouse death; conversely, IL-10 negatively regulates TNF-α and IL-6 release (Yimin and Kohanawa 2006; Yimin and others 2008). In this study, IL-4 deficiency resulted in pronounced reduction of not only TNF-α and IL-6 secretion in the organs and blood but also splenic and circulating levels of IL-10 during the initiation of infection in mice, which was accompanied by amelioration in mouse survival within 5 days postinfection. In line with these in vivo observations, low expression of TNF-α, IL-6, and IL-10, at both the gene and protein levels, was found in IL-4-deficient mouse macrophages on stimulation with heat-killed R. aurantiacus. Moreover, these cells also exhibited a decrease in NF-κB gene expression, which is related to the reduced levels of TNF-α and IL-6 because NF-κB regulates the genes encoding these cytokines (Lawrence and others 2005; Ji and others 2012). In addition, several studies showed that IL-4 has the ability to enhance IL-10 production in human monocytes and murine macrophages on lipopolysaccharide stimulation (Cao and others 2005; Woodward and others 2012). Therefore, it is evident that IL-4 deficiency attenuated macrophage activation, including both pro- and anti-inflammatory cytokine responses, during the initial phase of R. aurantiacus infection, which correlated with improvement of mouse survival. Similarly, a recent study also showed that IL-4 gene deficiency downregulates macrophage activation and protects mice from acetaminophen-induced tissue inflammation and death (Pires and others 2014). Interestingly, in contrast to the reduced levels of IL-10 in the spleens and blood of IL-4-deficient mice during initial infection, which was mainly derived from macrophages, the hepatic IL-10 levels in these mice were comparable to those of WT mice. Moreover, rapid enhancement of hepatic IFN-γ levels was also detected at day 1 postinfection in these gene-depleted animals. In the liver, unique and organ-specific immune systems that are composed of specialized cells such as Kupffer cells and NK cells (including NKT cells) are present, which play important roles in innate defense against pathogen infection. A previous study demonstrated that early IFN-γ secretion is induced in NK cells during R. aurantiacus infection (Asano and others 1995). Several studies further revealed the capacity of hepatic NK cells to rapidly produce not only IFN-γ but also IL-10 and IL-4 in early immune responses to diverse pathogens (Gao and others 2009; Hiraki and others 2012; Renna and others 2012). Our results also confirmed the early expression of IL-4 only in the livers of infected WT mice. Furthermore, IL-4 deficiency was reported to augment the effector functions of hepatic NK cells, such as cytokine production and cytotoxic activity (Saeftel and others 2004; Crowe and others 2005). Considering our observations and those of previous reports, we conclude that the activation of hepatic NK cells was improved under the condition of IL-4 deficiency, which induced the robust production of IFN-γ and IL-10. This augmentation of IL-10 is thought to compensate for the decreased macrophage production of this cytokine in the liver. Although IL-4 was recently reported to be dispensable for the function of NK cells in lymphoid tissues (Sharma and others 2013), we believe that this disparity may be due to a difference between the studied tissues.
The formation and regression of granulomas are strongly regulated by the cytokine microenvironment. Our previous study also demonstrated that early IFN-γ is required to activate macrophages to produce pro-inflammatory cytokines, including TNF-α, and the 2 cytokines have a synergistic effect on the subsequent development of granulomatous inflammation in R. aurantiacus infection (Yimin and Kohanawa 2006). In this study, IL-4-deficient mice displayed a marked increase in IFN-γ secretion but lower levels of TNF-α in the liver during the initiation of infection when compared with WT mice. In addition, a histological examination clearly showed a remarkable decrease in the formation of hepatic granulomas during the late phase in these gene-depleted mice. Thus, it is reasonable to suggest that IL-4 deficiency-induced alterations in the cytokine microenvironment weakened Th1 polarization, further confirming the importance of the synergistic effect of early IFN-γ and TNF-α production on granuloma formation. This study also demonstrates for the first time that IL-4 plays dual roles in the initial phase of infection by potentiating the macrophage response (including TNF-α, IL-6, and IL-10 secretion) while suppressing NK cell response (including IFN-γ and IL-10 secretion). Moreover, a previous report showed that IL-4 could promote the regression of granulomatous inflammation during the late phase of R. aurantiacus infection (Asano and others 1996). These studies reveal that even in a single animal model, IL-4 possesses a wide range of immunomodulatory activities in the different immune cells and at different phases of inflammation.
With regard to bacterial clearance, the rapid bacterial clearance observed in the livers of IL-4-deficient mice within day 3 postinfection can be explained by the enhanced release of early IFN-γ, as R. aurantiacus clearance depends on the presence of this cytokine (Yimin and others 2001; Yimin and others 2003). Although numerous studies have demonstrated the strong inhibitory effect of IL-4 on both nitric oxide synthesis and the macrophage-mediated intracellular killing capacity (Ho and others 1992; Cenci and others 1993; Reljic and others 2010), we did not find a statistically significant difference between IL-4-deficient mice and their WT counterparts either in the splenic bacterial load over the entire infectious period or in the hepatic bacterial load from day 5 postinfection. These results are consistent with the findings of Cole and others (2005), who indicated that IL-4 is not critical to bacterial clearance in the keratitis model induced by Pseudomonas aeruginosa infection.
In summary, we found that IL-4 deficiency attenuates the secretion of pro- and anti-inflammatory cytokines, including TNF-α, IL-6, and IL-10, in macrophages during the early stage of R. aurantiacus infection, which contributes to increased mouse survival and decreased granulomatous inflammation. IL-4 absence also resulted in improvement of the hepatic IFN-γ and IL-10 responses during the early stage; the augmented IFN-γ production induced by NK cells was responsible for a transient increase in bacteria clearance. Therefore, IL-4 appears to play dual roles in the innate inflammatory response to systemic R. aurantiacus infection by regulating the activation of immunocompetent cells, including macrophages and NK cells.
Footnotes
Acknowledgment
The authors would like to thank Yumiko Tsuda for mouse husbandry.
Author Disclosure Statement
No competing financial interests exist.