
Abstract
ISG15 conjugation (ISGylation) to proteins is a multistep process involving interferon (IFN)-inducible UBE1L (E1), UbcH8 (E2), and ISG15 E3 ligases (E3s). Studies performed over the past several years have shown that ISGylation plays a pivotal role in the host antiviral response against certain viruses. Recent in vitro studies revealed that human Herc5 and mouse Herc6 are major ISG15 E3 ligases, respectively. However, the global function of Herc5/6 proteins in vivo still remains unclear. Here, we report generation and initial characterization of Herc6 knockout mice. Substantial reductions of ISGylation were observed in Herc6-deficient cells after polyinosinic-polycytidylic acid double-stranded RNA injection of mice or IFN treatment of cells. On the other hand, Herc6-deficient cells and wild-type (WT) cells had similar responses to IFN stimulation, Sendai virus (Z strain) infection, and vesicular stomatitis virus infection. These results indicate that Herc6 does not play a critical role in antiviral defense of these viral infections in mice. Interestingly, male Herc6-deficient mice showed seminal vesicle hypertrophy. No such problem was detected in WT and ISG15 activating enzyme Ube1L-deficient mice. These results suggest that in addition to promoting protein ISGylation, Herc6 has a novel and protein ISGylation-independent function in the male reproductive system.
Introduction
ISG15
ISG15-mediated antiviral activity against influenza, herpes, and Sindbis virus has been shown in vivo by means of infections in ISG15−/− and UBE1L−/− mice (Lenschow and others 2005, 2007; Lai and others 2009; Lenschow 2010). In addition, in vitro studies for either the overexpression of ISG15 or knockdown of ISG15 using siRNA have implicated ISGylation in the regulation of influenza B virus, vaccinia virus, Sindbis virus, herpes simplex-1 virus, Sendai virus, and Japanese encephalitis virus, as well as in the release of virus-like particles derived from HIV-1 and avian sarcoma leukosis virus (Okumura and others 2006, 2008; Guerra and others 2008; Malakhova and Zhang 2008; Hsiang and others 2009; Hsiao and others 2010; Lenschow 2010; Pincetic and others 2010). Although the mechanism by which ISG15 is regulating viral growth is still unknown for the majority of these viruses, it has been reported that ISG15 achieves its antiviral role by conjugating to target proteins, including both host proteins and viral proteins, and altering their functions. For example, ISG15 can be conjugated to host antiviral protein interferon regulatory factor 3 (IRF3) and, thus, stabilize IRF3 by inhibiting its interaction with peptidyl-prolyl cis-trans isomerase NIMA-interacting 1 (PIN1), a protein that promotes IRF3 ubiquitination and degradation (Shi and others 2010). In most overexpression studies, ISG15 and its conjugation-related enzymes have been coexpressed, suggesting that ISG15 conjugation to target proteins is required for these antiviral effects. Despite these observations with either in vivo or in vitro studies, there are some controversial phenotypes: No differences in viral growth of influenza A, herpes simplex virus-1, Sindbis, and wild-type (WT) vaccinia virus in ISG15−/− mouse embryonic fibroblast cells (MEFs) and vesicular stomatitis virus (VSV) in UBE1L−/− MEFs have also been reported (Osiak and others 2005; Kim and others 2006; Lenschow and others 2007; Guerra and others 2008).
A recent report indicated that human Herc5 globally targets de novo synthesized proteins for ISG15 conjugation, thereby making viral proteins major targets for ISGylation (Durfee and others 2010). However, the question remains whether Herc5 and global ISGylation are important for antiviral activity in vivo, as ISGylation-mediated antiviral effects might be due to other minor ISG15 E3 ligases with more narrow specificity, such as estrogen-responsive finger protein (EFP, also called TRIM25) (Park and others 2014). Since there is no mouse ortholog of human HERC5, the other members of HERC family proteins have been examined for ISG15 E3 activity. A recent report shows that Herc6 knockdown in mouse L929 cells abolished global ISGylation, whereas its overexpression enhanced ISGylation as well as IFN-β production, and conferred antiviral activity against vesicular stomatitis virus and Newcastle disease virus, which indicated that Herc6 is likely the functional antiviral factor in mouse cells (Oudshoorn and others 2012).
Here, we established the Herc6 null mice to study their role in protein ISGylation in vivo. Herc6 knockout mice lacked protein ISGylation. Herc6-deficient primary MEFs and bone marrow-derived macrophages (BMDM) lost the capacity to conjugate ISG15 to a broad group of proteins. These analyses with our newly generated Herc6 knockout mice confirmed the previous finding in the in vitro system that Herc6 is the major ISG15 E3 ligase in mice. Furthermore, we examined STAT1 phospharylation on IFN-β treatment, production of IFN-β and IL-6 after SV infection, or double-stranded RNA poly I:C treatment of WT and Herc6−/− BMDM. No significant difference was detected. In addition, the virus titers of VSV in WT and Herc6−/− MEFs were similar. These results indicate that Herc6-mediated protein ISGylation has no obvious effect on IFN signaling and antiviral activity against SV and VSV under current experimental conditions. As a critical different phenotype from ISG15 E1 Ube1L knockout mice that also lack protein ISGylation, male Herc6-deficient mice showed severe seminal vesicle hypertrophy. This finding suggests that Herc6 has a role in regulating sperm sac morphology, which is independent of protein ISGylation.
Materials and Methods
Generation of Herc6 knockout mice
The Herc6 mutant mice (accession No. CDB0585K;
Animal studies
CBA/C57BL6 Mix background Herc6 KO (4 times backcrossed) mice and C57BL6 pure background WT and Herc6 KO mice were maintained at Kobe BM laboratory (Oriental Bio Service, Inc.). C57BL6 pure background WT and UBE1L KO mice were maintained at UCSD Moores cancer center. Animal studies in Kobe BM laboratory were properly conducted in accordance with regulations regarding animal experiments in Japan. Animal experiments in UCSD Moores cancer center were performed in accordance with NIH policies on the use of laboratory animals and approved by the Animal Research Committee of the Office for the Protection of Research Subjects at the University of California, San Diego.
Southern blotting and PCR genotyping
Southern blots were performed using a radioactive label or DIG label (Roche) methods according to the manufacturer's protocols. The probe for Southern blotting was amplified using primers as follows: Fw (5′-TGA AGACAG ACA AGG TGG AAT AAC TTG ATA-3′), Rev (5′-CAG CTG CAG TAC CAC AGG TGA TGT GGT ACT-3′). The genotyping of mice was routinely performed with tail by PCR using a mixture of 3 primers after overnight treatment with proteinase K in PCR buffer. The sizes of the PCR products are WT allele (258 bp) and mutant allele (787 bp). The PCR primers were as follows: P1 (5′-ACA GGA TGT GAT AGG CTG CAT GTG AAA G-3′) and P3 (5′-AAA CAC CTA GTT CCC GAG GCT GTG AAC T-3′) for the Herc6 WT allele; P2 (5′-ATC AGG ATG ATC TGG ACG AAG AGC ATC A-3′) and P3 for the Neo gene. The PCR conditions were 95°C for 2 min, 40 cycles of 95°C for 30 s, 55°C for 30 s, and 72°C for 1 min, followed by 72°C for 10 min.
Cell culture and transfection
MEFs were prepared from E12.5 embryos and grown in Dulbecco's modified Eagle medium supplemented with glutamine, penicillin/streptomycin, and 10% fetal bovine serum (FBS). BMDM were cultured in RPMI1640 medium supplemented with glutamine, penicillin/streptomycin, 10% FBS, and macrophage colony-stimulating factor. For the knockdown analysis, siRNA was transfected using Lipofectamine 2000 (Invitrogen) according to the manufacturer's instruction. At 72 h after the siRNA transfection, cell lysates were prepared for examination.
Antibodies and reagents
Anti-mouse Herc6 polyclonal antibodies were generated in rabbits by using the mouse Herc6 recombinant protein that was bacterially produced using GST (6P-1) mHerc6 652-1003aa. Antibodies against STAT1, p-STAT1 (Y701), and ISG15 (#2743) were purchased from Cell Signaling. For the detection of protein ISGylation in liver and spleen tissue samples of WT and Herc6 knockout mice with or without poly I:C injection, and BMDM samples of WT and Herc6 knockout mice with or without IFN-β treatment, we used Dong-Er Zhang Lab's anti-mouse ISG15 antibody. Anti-α-tubulin (Sigma) and actin were acquired from Oncogene Research Products. Poly I:C was purchased from Amersham. Mouse IFN-β was purchased from PBL. Sendai virus (Z strain) and VSV were kindly provided by Dr. Masato Nakanishi (Research center for stem cell engineering, AIST, Japan).
Knockdown
For knockdown of Herc6, the following siRNAs were used:
si-Herc6-1: 5′-gaaauaagcuuuaugccuauu-3′ (B-Bridge International, Inc.)
si-Herc6-2: 5′-ggaacaaaguuaaagaacauu-3′ (B-Bridge International, Inc.)
si-Herc6-3: 5′-ccuacagaaugaaggaauauu-3′ (B-Bridge International, Inc.)
Control siRNA (si-GFP); 5′-acuuguacagcucguccauuu-3′ (B-Bridge International, Inc.)
Western blotting
Western blotting was conducted as previously described (Arimoto and others 2010). All samples were denatured in 1× sample buffer [50 mM Tris-HCl (pH 6.8), 2% sodium dodecyl sulfate (SDS), 2-mercaptethanol, 10% glycerol, and 1% bromophenol blue] for 5 min at 100°C. Cells were lysed in RIPA buffer composed of 25 mM Tris-HCl (pH 8.0), 150 mM NaCl, 1 mM EDTA, 1 mM dithiothreitol, 0.1% SDS, 1% Nonidet P-40, and 0.5% sodium deoxycholate. The cell lysates were centrifuged (10,000 g) at 4°C for 5 min. All lysis buffer in this study contains proteinase and phosphatase inhibitors (Roche). For the quantification, the Fujifilm Multi-gauge V3.0 was used.
Enzyme-linked immunosorbent assay
Culture media were collected and analyzed for IFN-β and IL-6 production by using enzyme-linked immunosorbent assays (ELISAs). ELISA kits for mouse IFN-β and IL-6 were purchased from PBL Biomedical Laboratories.
TCID50 assay
50% tissue culture infectious dose (TCID50) assay was conducted as previously described (Arimoto and others 2010). Approximate viral titers were calculated by TCID50 assay. Results of this assay are well in accordance with those of the general plaque assay. After 12 h VSV infection, the culture medium was diluted 3×104 times and then added to the first line of a 96-well plate with 50 μL medium containing 293T cells, making a serial 3-fold dilution. At 1–2 days after infection, more than 50% cell alterations in each dilution step was analyzed with the following formula: TCID50=(rate of dilution at first line)×(dilution rate)Σ−0.5, where Σ=the number of wells observed with more than 50% cell alteration in each dilution step/sample sum.
Histology
Whole body was fixed by 10% formaldehyde solution, and tissues were paraffin embedded into OCT compound (Tissue-Tek). For histochemical analysis, paraffin sections were stained with H&E according to standard protocols.
Results
Disruption of the mouse Herc6 gene
In line with the previous report using L929 cells and siRNA against Herc6 (Oudshoorn and others 2012), we observed the reduction of ISGylation in Herc6 knockdown primary MEFs after IFN treatment (Supplementary Fig. S1; Supplementary Data are available online at
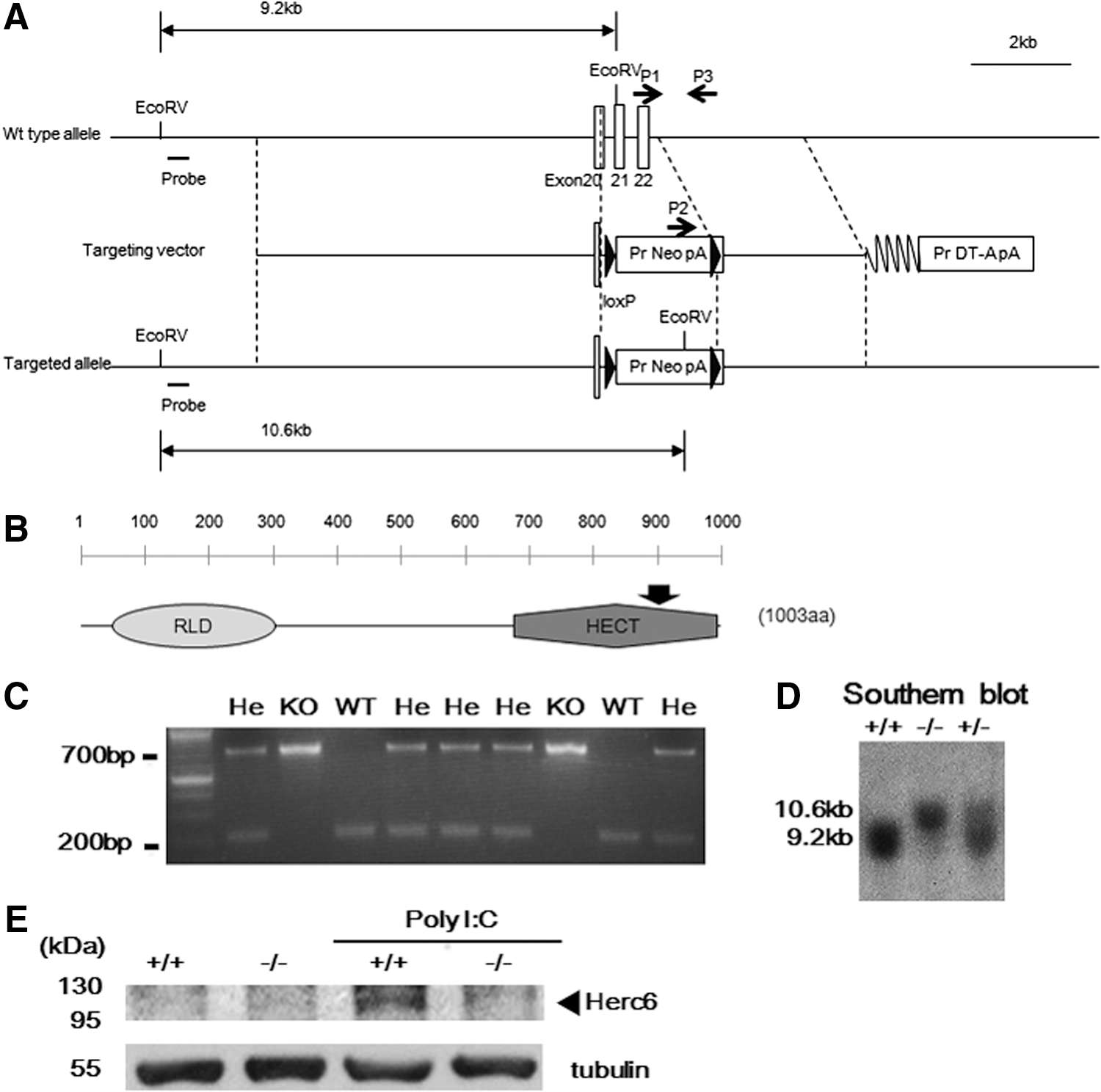
Generation of Herc6 knockout mice.
Defective protein ISGylation in Herc6 knockout mice
To determine whether lack of Herc6 correlated with decreased ISGylation in vivo, we injected WT and knockout mice with PBS or poly I:C. Twenty-four hours later, spleens and livers were harvested, and protein ISGylation was detected by Western blotting. Protein ISGylation was readily detected in the liver and spleen from WT but was barely detected from Herc6 knockout mice (Fig. 2A and B, respectively). Similar results were observed in IFN-β-treated MEFs and BMDM from WT and Herc6 knockout mice (Fig. 2C and D, respectively). Herc6-deficient tissues or cells showed an increased amount of free ISG15 as a result of the lack of conjugation (Fig. 2A–D).

Analysis of protein ISGylation in Herc6 knockout mice.
These results indicate that Herc6 knockout mice are defective in protein ISGylation but not in free ISG15 expression.
Normal IFN responses of Herc6-deficient cells and mice
A previous report has shown that human HERC5 positively regulates the IFN-β promoter via enhancing IRF3 function and so confers antiviral activity (Shi and others 2010). In addition, mouse Herc6 also enhanced IFN-β promoter activity similar to its human Herc5 counterpart (Oudshoorn and others 2012). To explore whether Herc6 regulates IFN signal transduction, we examined the IFN-response of Herc6-deficient cells. Macrophages derived from bone marrow cells of WT and Herc6 knockout mice were cultured in vitro and treated with 100 U/mL of IFN-β, and STAT1 phosphorylation was detected as an indication of the activation of the signaling pathway. Increased phosphorylation of STAT1 on IFN treatment was observed. However, there was no difference of STAT1 phosphorylation between WT and Herc6-deficient cells (Fig. 3A).
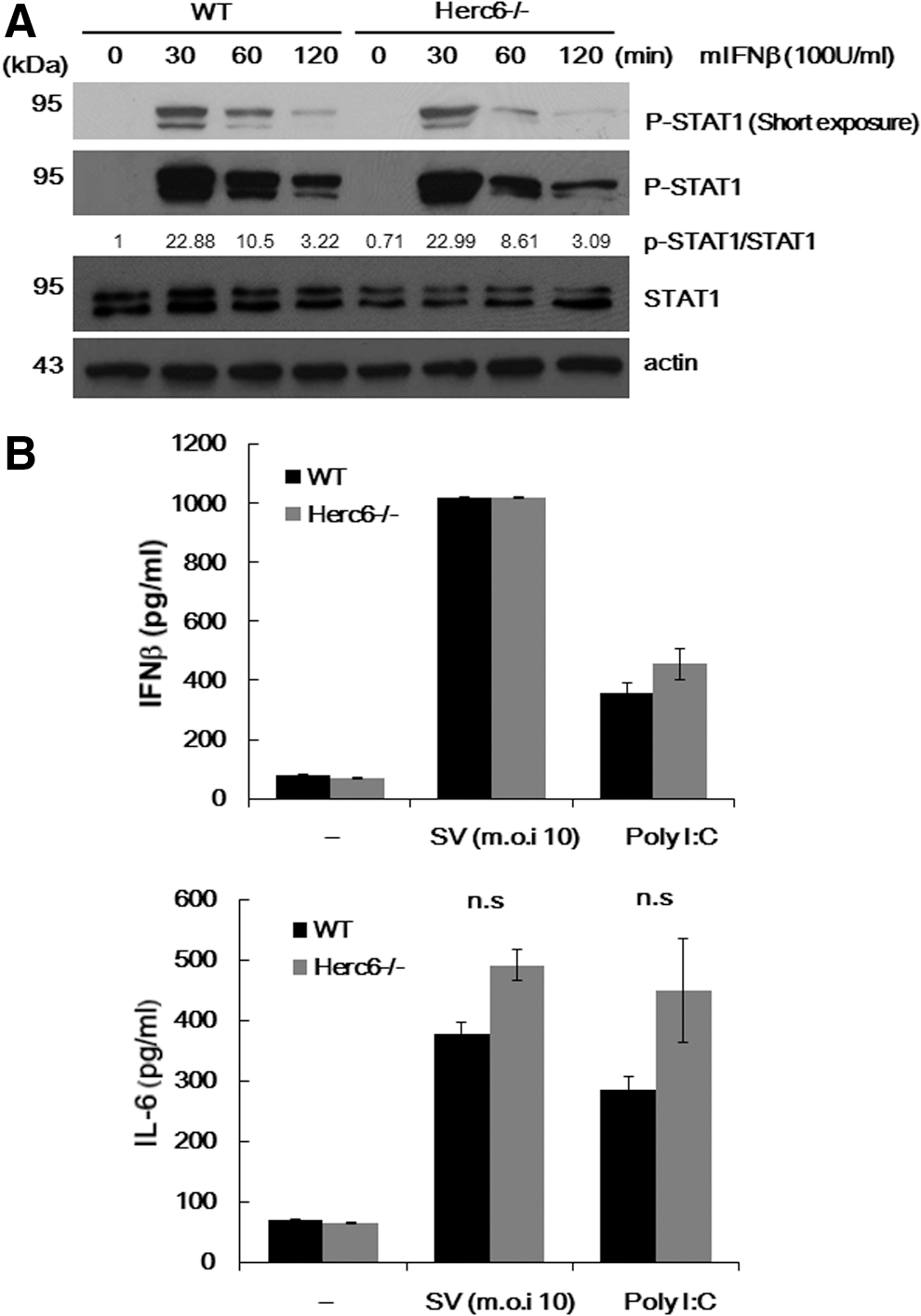
Analysis of IFN and inflammatory response in Herc6 knockout cells.
To investigate whether the mouse Herc6 enhances an innate immune signal, we examined the IFN-β and IL-6 production in WT and Herc6−/− MEFs on Sendai virus infection and poly I:C stimulation. No significant differences in WT and Herc6 cells were observed (Fig. 3B top and bottom, respectively). Although it was not significant, a modest increase of IL-6 production in Herc6−/− MEFs with SV infection or poly I:C stimulation was observed (Fig. 3B bottom). This should be investigated with greater detail in the future.
Since lipopolysaccharide (LPS) activates the expression of these genes via the IFN signaling pathway, we also examined the peripheral blood cell count and other blood parameters. However, no significant differences were observed between WT and Herc6 knockout mice with or without LPS stimulation (Table 1).
Blood samples were collected via retro-orbital bleeding from wild-type and Herc6 knockout mice at 1 day after with or without LPS (15 mg/kg body weight) injections. Values are means±standard deviation.
WBC, white blood corpuscles; RBC, red blood corpuscles; Hb, hemoglobin; Ht, hematocrit; MCV, mean corpuscular volume; MCH, mean corpuscular hemoglobin; MCHC, mean corpuscular hemoglobin concentration; PLT, platelet; LPS, lipopolysaccharide; WT, wild type.
In summary, Herc6-deficient cells do not show any detectable differences from WT cells in their responses to IFN-related treatments.
Deletion of Herc6 did not affect the antiviral response against VSV infection
Besides the IFN response (Shi and others 2010), a recent report indicated that Herc5 mainly conjugates ISG15 to newly synthesized proteins in tissue culture and may by this mechanism largely target de novo synthesized viral proteins during infection (Durfee and others 2010).
To investigate whether Herc6 affects the proliferation of virus, we examined the consequences of reduction of protein ISGylation on the antiviral effects of IFN in MEFs. Treatment with increasing concentrations of IFN-β correlated positively with the antiviral stage of both WT and Herc6 knockout MEFs on infection with VSV, with no detectable differences in the response between the 2 genotypes (Fig. 4A). In addition, there was no significant difference of viral titer between WT and Herc6 knockout MEFs after infection with VSV (Fig. 4B). These experiments demonstrate that protein ISGylation via Herc6 in mice is not involved in the antiviral response against VSV.
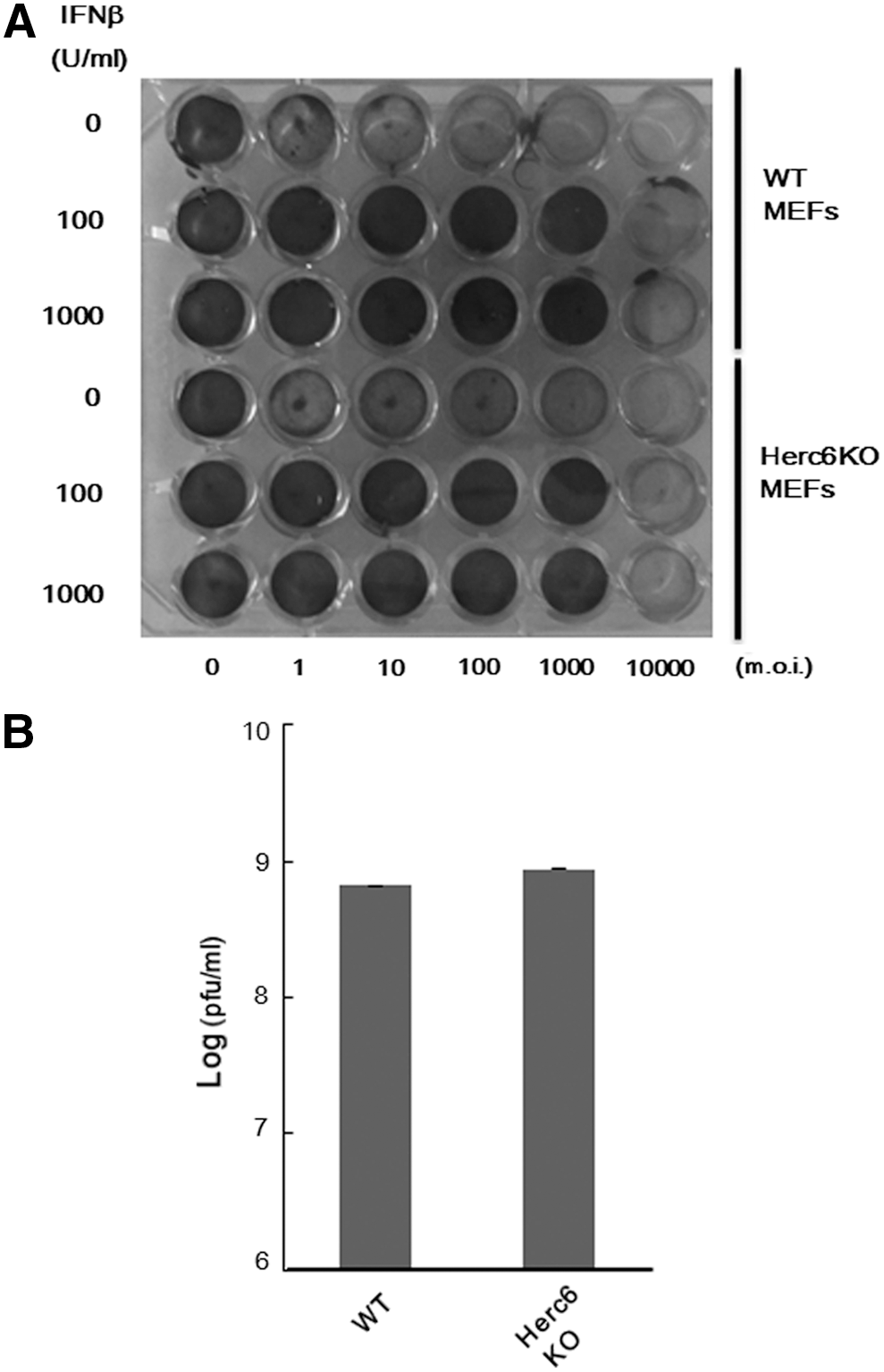
VSV protection assay.
Male Herc6 null mice showed hypertrophy of seminal vesicle
During the experiments, we noticed that mix-background Herc6 knockout male mice began showing abdominal distension at 28 weeks. Severe enlargement seminal vesicles were observed in Herc6 knockout mice (Fig. 5A). However, only slight epithelial hyperplasia with cystic dilatation of seminal vesicles was observed in Herc6 knockout mice (Fig. 5B), suggesting that the enlargement of seminal vesicles was from increased seminal fluid. Glandular hypoplasia and benign hypertrophy of the prostate, which are commonly found in elderly men, were not observed in Herc6 knockout mice (data not shown). Further examination using C57BL6 background mice showed a higher frequency of enlargement of seminal vesicles in Herc6 knockout mice compared with WT or UBE1L knockout mice (Table 2 and Supplementary Fig. S2). These results demonstrate that mouse Herc6 has an ISG15-independent function in regulating the seminal component.

The phenotype of Herc6 knockout mice.
One-side anomalistic seminal hypertrophy.
Seminal hypertrophy is defined as above at least 0.04 g seminal vesicle/g body weight.
Discussion
In this article, we described the generation and analysis of the first Herc6 knockout mouse model. The results of our studies demonstrate that (1) Mouse Herc6 is the major E3 ligase for ISG15 conjugation in vivo. (2) ISG15 conjugation via mouse Herc6 does not affect type I IFN response and antiviral response against SV and VSV infection. (3) Mouse Herc6 possesses an ISG15 conjugation-independent role in regulating sperm sac morphology.
Previous reports indicate that human Herc5 knockdown in 293T cells and mouse Herc6 knockdown in L929 cells showed reduced ISG15 conjugation to a broad group of proteins after IFN treatment (Dastur and others 2006; Wong and others 2006; Oudshoorn and others 2012). In line with this result, the level of ISG15 conjugation in tissues of Herc6-deficient mice showed substantial reduction compared with that of WT mice after poly I:C injection. These findings clearly indicate that Herc6 is the major mouse ISG15 E3 ligases in vivo. Furthermore, our report supports that humans and mice developed different Herc proteins to facilitate global ISG15 conjugation during evolution.
A recent report indicates that HERC5 is mainly associated with poly-ribosome, and ISGylation targets newly synthesized proteins in human tissue culture (Durfee and others 2010). Furthermore, in this same report, the authors showed that HPV16 L1 capsid protein was ISGylated and this modification inhibited HPV pseudovirus production in transfection experiments (Durfee and others 2010). In addition, previous reports showed that antiviral effects exerted by human HERC5 (Shi and others 2010) are shared by Herc6 in mouse cells (Oudshoorn and others 2012). ISG15-activating enzyme Ube1L knockout mice also showed no protein ISGylation and no difference in IFN responses and anti-VSV and LCMV defense (Osiak and others 2005; Kim and others 2006). However, further studies of Ube1L knockout mice revealed a critical role of Ube1L in control of influenza B virus infection (Lai and others 2009). These discrepancies of revealed functions of protein ISGylation are likely due to the differences in species (humans and mice), lines, and viruses, and in vitro versus in vivo experiments. These questions need to be addressed in the future.
Herc6 null mice showed hypertrophy of seminal vesicles in both CBA/C56BL6 mixed background and C57BL6 background. However, Ube1L−/− mice did not show similarly enlarged sperm sacs although protein ISGylation is lost in both Herc6 and Ube1L knockout mice (Kim and others 2006). This difference suggests that mouse Herc6 has an ISG15-independent function in seminal fluid production during aging. However, the phenotype of Herc6−/− ISG15−/− also should be investigated in the future to examine whether this seminal hypertrophy is truly Herc6 dependent and ISG15 independent. Since mouse Herc6 belongs to the Herc family of ubiquitin E3 ligases, it is possible that ISG15 conjugation is regulated by the ISG15 E3 ligase activity of Herc6, and seminal component secretion is regulated by its ubiquitin E3 ligase activity. Human Herc5 may have evolved to exclusively function as an ISG15 E3 ligase, while human Herc6 may function as a ubiquitin E3 ligase that is involved in the seminal component secretion.
While we could not see the difference in male fertility of Herc6 knockout mice (∼35 weeks) (data not shown), we cannot rule out this possibility because it could be affected by the conditions associated with older age.
The function of seminal fluid is largely unknown except for the involvement of normal conception. Interestingly, it has recently been reported that paternal seminal fluid composition affects the epigenome of male offspring and that its impact on the periconception environment involves not only sperm protection but also indirect effects on various female factors regulating embryo development, which suggests that offspring of Herc6 null mice may have interesting epigenomic changes by the hypertrophied seminal vesicles of Herc6 male mice (Bromfield and others 2014). Future studies using this mouse model may facilitate to address this question.
In humans, symptoms of enlarged seminal vesicles are frequently seen in patients with ejaculatory duct obstructions (EDO) (Pryor and Hendry 1991). EDO is a congenital or acquired pathological condition that is characterized by the obstruction of one or both ejaculatory ducts, and causes 1%–5% of male infertility (Philip and others 2007). In addition to the congenital form that is often caused by cysts of the müllerian duct, the obstruction can be acquired due to an inflammation caused by chlamydia, prostatitis, tuberculosis of the prostate, and other pathogens (Philip and others 2007). However, in many patients, there is no history of inflammation and the underlying cause simply remains unknown (Philip and others 2007). The finding that the suppression of Herc6 caused the hypertrophied seminal vesicles in this report may be involved in the acquired enlarged seminal vesicle symptoms, and this should be investigated in the future.
Footnotes
Acknowledgments
The authors thank all lab members for helpful discussion. They especially thank Dr. Kentson Lam (School of Medicine, University of California San Diego) and Samuel Stoner (Moores Cancer Center, University of California San Diego) for discussion and a critical reading of this article. They also thank Shuichiro Ogawa (Kyoto University) for several experiments and discussion. This study was supported by Grants-in-Aid for Scientific Research from the Ministry of Education, Culture, Sports, Science, and Technology (22114004 & 22249012), funding from National Institutes of Health USA (R01CA177305 and R01HL091549), and JSPS KAKENHI Grant (No. 10J00577); Kei-ichiro Arimoto is a JSPS Postdoctoral Fellow for Research Abroad.
Author Disclosure Statement
There is no financial interest to disclose.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.