
Abstract
Many malignant tumors express high levels of the chemokine receptor CXCR4, and the interaction between CXCR4 and its ligand, SDF-1, promotes migration, invasion, and metastasis of breast cancer cells. Therefore, blocking the interaction between CXCR4 and SDF-1 could alter the tumor's metastatic phenotype and control the development and progression of cancers. We used a cellular phenotypic knockout strategy and developed a novel recombinant gene, AdSDF-1α/54/KDEL, which contains an adenovirus vector, a mutant form of SDF-1 that lacks a C-terminal α-helix, and a KDEL tetrapeptide sequence that promotes retention at the endoplasmic reticulum (ER). We hypothesized that SDF-1α/54/KDEL could efficiently block metastasis of breast cancer cells with less inflammatory side effects than SDF-1α/KDEL. Using the MCF-7 cell line, which expresses a stable, high level of CXCR4, we found that SDF-1α/54/KDEL efficiently becomes localized at the ER of tumor cells, where it specifically binds to newly synthesized CXCR4 and prevents it from reaching the cell surface. Chemotaxis and invasion assays revealed that the cells treated with SDF-1α/54/KDEL failed to migrate toward SDF-1. We also found that SDF-1α/54/KDEL impaired lung metastasis of metastatic breast cancer by decreasing CXCR4 on the cell surface. The novel recombinant gene, SDF-1α/54/KDEL, played an instrumental role in blocking SDF-1/CXCR4-mediated cell migration, and we found that this gene-based strategy for targeting the SDF-1/CXCR4 axis offers a very effective alternative method for preventing metastasis of breast cancer and other cancers expressing high levels of CXCR4.
Introduction
E
We previously developed a human mutant SDF-1α/54 protein, which contains a defective C-terminal alpha helix, a normal N-terminus, and a normal central beta-strand scaffold structure, that retains a physiological binding affinity to CXCR4, induces CXCR4 internalization, and prevents CXCR4-mediated cell signaling and chemotaxis. It was confirmed in vitro that this novel mutant of SDF-1 quickly promoted CXCR4 internalization and eliminated CXCR4 from the cell surface without activating downstream signals, which, in turn, inhibited tumor cell migration (Tan and others 2006). However, this protein level inhibitory effect of SDF-1α/54 appeared to be transient due to the subsequent transport of internalized CXCR4 to the cell membrane (Ma and others 2009b). Thus, it is necessary to develop strategies to prolong this inhibition of CXCR4 expression on the tumor cell surface. CXCR4 phenotype knockout strategies have been widely used to inhibit tumor metastasis mediated by the CXCR4/SDF-1 axis (Wheeler and others 2003; Bjorndahl and others 2004; Hu and others 2005; Meijer and others 2006). CXCR4 expression and function in CT-26 colon carcinoma cells were inhibited by transduction of SDF-1 extended with a KDEL sequence, and this strategy blocked metastasis to the liver and lung (Zeelenberg and others 2003). KDEL is a tetrapeptide sequence that, when inserted at the C-terminus of SDF-1, transports CXCR4 into the lumen of the endoplasmic reticulum (ER), where it is unable to be secreted due to constant recycling at the Golgi apparatus (Yamamoto and others 2001; Capitani and Sallese 2009; Alanen and others 2011).
The use of an intracellular antibody (intrabody) or intracellular chemokine (intrakine) to inhibit the expression of target proteins offers potential applications for AIDS prevention and treatment, hematopoietic stem cells, immunology, embryonic stem cells, and cancer research (Wheeler and others 2003; Bjorndahl and others 2004; Hu and others 2005; Tan and others 2005; Meijer and others 2006; Zhang and others 2009; Vallera and others 2010). This strategy prolongs the transport inhibition of newly synthesized target proteins to the cell surface. Studies have shown that a lentivirus-mediated technique for intrakine gene transfer targeting RANTES protected human T-lymphocytes from infection by a variety of R5-tropic HIV-1 strains (Schroers and others 2002). The viral vector provides an effective means for intrakine gene transfer; however, the SDF-1α/KDEL intrakine is partly secreted and downregulates the CXCR4 receptor on nontransduced cells (Engel and others 2000). In addition, the wild-type SDF-1 chemokine functions as an autocrine agent, acting as a chemoattractant for leukocytes, macrophages, and tumor cells. These chemotactic effects produce an inflammatory response in the tumor microenvironment and promote tumorigenesis. Therefore, to avoid these harmful effects, we investigated the use of the SDF-1α/54 mutant as a more appropriate intrakine.
We hypothesized that the C-terminus of SDF-1α plays an important role in the chemotaxis of breast cancer cells and that we could prevent the metastasis of breast cancer by deleting the alpha helical domain to produce a more efficient, high-affinity intracellular chemokine. SDF-1α/54 was linked to the KDEL ER retention signal to form an intrakine to which a recombinant adenovirus vector could bind and inactivate newly synthesized CXCR4 within the tumor cell. This inactivation of CXCR4 prevents its transport to the cell surface, and the SDF-1α/54/KDEL-CXCR4 protein complex is degraded in the ER lumen. Therefore, this strategy prevents tumor metastasis by producing a more effective and prolonged neutralization of CXCR4. We used the MCF-7 and MDA-MB-231 cell lines, which both express stable, high levels of CXCR4, to investigate the ability of AdSDF-1α/54/KDEL to efficiently transport newly synthesized CXCR4 into the ER lumen rather than to the cell surface of tumor cells. Thus, if CXCR4 fails to reach the cell surface, these tumor cells fail to migrate toward the gradient concentration of SDF-1 to signal proliferation and metastasis. Furthermore, we explored the systemic inhibitory effects in vivo of AdSDF-1α/54/KDEL on lung metastasis with CXCR4-positive breast cancer cells in the transplanted tumor mouse model.
Materials and Methods
Reagents
The TRIzol and Lipofectamine 2000 kit (Invitrogen), Taq polymerase, the polymerase chain reaction (PCR) fragment recovery kit, the DNA ligation kit, XhoI, XbaI, PacI, PmeI, alkaline phosphatase (CIAP), DL2000+DL15000 DNA Marker, the E.Z.N.A. Plasmid Miniprep Kit (Omega Bio-tech), protein marker (Fermentas Life Sciences), recombinant human SDF-1α, matrigel, and fibronectin (Becton Dickinson), the transwell chamber (Kurabo), goat anti-hSDF-1 antibody, alkaline phosphatase rabbit anti-goat IgG (H+L) (Zhongshan), phycoerythrin (PE) mouse anti-human CXCR4 (fusin) monoclonal antibody (Clone:12G5; R&D), and goat anti-mouse IgG-fluorescein isothiocyanate (FITC) (BOSTER).
Plasmids and Escherichia coli
Adenovirus backbone plasmid pAdeasy-1, pAdtrack-CMV, and BJ5183 were purchased from Stratagene. DH5α competent cells were purchased from Sun Yat-sen University (Guangzhou, China).
Mice and cell lines
Four- to six-week-old female BALB/c nude mice were obtained from the Experimental Animal Center of Sun Yat-sen University, People's Republic of China (Certification No. 2004A084). Our protocol conforms with the Guide for the Care and Use of Laboratory Animals published by the US National Institutes of Health (NIH Publication No. 85-23, revised 1996) and was approved by the Ethics Committee of Guangdong Pharmaceutical University Animal Center. MCF-7 cells were maintained in Dulbecco's modified Eagle's medium (DMEM) supplemented with 4 mM
AdSDF-1α/54/KDEL construction and expression
The SDF-1α/54/KDEL gene was amplified by PCR from a pcDNA3.1/SDF-1α/54/KDEL vector (Ma and others 2009b). The sense strand for the SDF-1α/54/KDEL primer was 5′-ACACTCGAGATAAGCCCGTCAGCCTGAGCTACAGAT-3′, and the anti-sense strand was 5′-GCTCTAGAGAGCTCCTTCTT CTTCGGGTCAATGCACAC-3′. The sense strand for the SDF-1α/KDEL primer was 5′-ACACTCGAGATAAGCCCGTCAGCCTGAGCTACAGAT-3′, and the anti-sense strand was 5′-GCTCTAGATCCTAGAGCTCGTCCTTCATCTTGAACCTCTTGTT-3′. The PCR products were 174 and 228 bp, respectively. The shuttle plasmid, pAdTrack-CMV, was cut by XbaI and XhoI, and SDF-1α/54/KDEL was subcloned into the plasmid vector to construct pAdTrack-CMV-SDF-1α/54/KDEL. It was then homologously recombined with pAdEasy-1 to construct AdSDF-1α/54/KDEL inside the host bacterium. AdSDF-1α/54/KDEL was then transfected with Lipofectamine 2000 into HEK-293 cells, and the viral plaque appeared 7 days after transfection.
Cell proliferation assay
The effects of AdSDF-1α/54/KDEL on growth inhibition were evaluated by MTT assays. MCF-7 or MDA-MB-231 cells were plated at a density of 1×104 per well in 96-well microplates and incubated overnight, and recombinant adenovirus-encoding intrakines with multiplicity of infections (MOIs) ranging from 1 to 200 were added to the wells. After 48 h, each well was treated with 20 μL of 5 mg/mL MTT solution, and the cells were further incubated at 37°C for 4 h. The untransformed MTT was then removed and 150 μL dimethyl sulfoxide (DMSO) was added. After culture absorbance, the absorption (A) was determined at 490 nm. The cell growth inhibition rate was calculated according to the following formula: cell growth inhibition rate (%)=1−(mean cells of treatment group/mean cells of control group)×100. The mean of 3 wells was calculated, the measurements were repeated 3 times, and the results were averaged.
CXCR4 receptor expression analysis
MCF-7 or MDA-MB-231 cells (1.5×106) were infected with 50 MOI of AdSDF-1α/54/KDEL, control SDF-1α/KDEL, or no insert AdNull (Mock). Cells were analyzed for CXCR4 surface expression at 72 h after infection by flow cytometry. Cells were stained with PE-conjugated mouse anti-human CXCR4 monoclonal antibody or PE-conjugated mouse IgG2a isotype control.
Chemotaxis assay
Migration of tumor cells toward SDF-1 was determined by a transwell migration assay as previously described (Smith and others 2004) with modifications. MCF-7 or MDA-MB-231 cells were assessed in disposable transwell trays with 6-mm diameter chambers and a membrane pore size of 8 or 5 mm. Cells were preincubated with various concentrations of AdSDF-1α/54/KDEL and AdSDF-1α/KDEL at 37°C for 12 h. Two hundred microliters of preincubated cells was added to the upper wells, and 700 μL of diluted SDF-1α (0.1 μmol/L) was added to the lower wells. RPMI containing 0.25% human serum albumin with 300 ng/mL recombinant human SDF-1α served as a control. After a 2-h incubation, the cells that migrated to the lower wells were counted. Migrating cells without any treatment were used as negative controls. CXCR4 neutralization studies were performed with anti-human CXCR4 monoclonal antibody (10 pg/mL). Each experiment was performed in triplicates, and the data were averaged for statistical analysis.
Invasion inhibition assay
Cells were washed twice and suspended at 2×106 cells/mL in a medium containing DMEM with 0.1% BSA. A 24-well plate containing matrigel and fibronectin was used for this experiment. Cells were pretreated with a control (DMSO) or different concentrations of inhibitors for 24 h. One hundred microliters of cells from each sample was loaded into the upper well, and the lower well contained 0.6 mL of medium with SDF-1α (30–300 n
Effectiveness of AdSDF-1α/54/KDEL on tumors in vivo
To study the effects of AdSDF-1α/54/KDEL on tumor volume and weight, mice were inoculated with 3×105 MDA-MB-231 cells. When a palpable tumor was observed (at day 9), the mice were divided randomly into 4 groups of 6 mice each that were controls or treated with AdSDF-1α/54/KDEL or SDF-1α/KDEL. Eight days later, AdNull, AdSDF-1α/54/KDEL, or SDF-1α/KDEL (at 5×108 pfu in 100 μL) was injected into the tumor, and tumor volume and weight were calculated at 3-day intervals from days 8 to 30 after grafting. Results are expressed as mean tumor volume and mean tumor weight±SD in mm3 and grams, respectively. We also used this protocol to study the effects of AdSDF-1α/54/KDEL on tumor metastasis and survival. Tumor size was measured 2 times per week, and survival was monitored for 12 weeks. If a mouse appeared moribund or if the diameter of the tumor reached 20 mm, the animal was sacrificed, and this was recorded as the date of death for survival studies. Survival of the animals was assessed using the standard methodology. Liver and lung tissues were removed sterilely from the tumor-bearing mice for RT-PCR testing. The mRNA was extracted with TRIzol, and gene-specific primers were used to detect the expression of CXCR4 and SDF-1α. The sense strand for the CXCR4 primer was 5′ -ATAAGAATGCGGCCGCTATGGAGGGGATCAGTATATACACTT-3′, and the anti-sense strand was 5′-CGCTCTAGATTAGCTGGAGTGAAAACTTGAAGACT- 3′. The sense strand for the SDF-1α primer was 5′-GCAAGCCCGTCAGCCTGAGCTACAGAT-3′, and the anti-sense strand was 5′-CTTCGGGTCAATGCACACTTGTC-3′. The sense strand for the β-actin primer was 5′-GAGACCTTCAACACCCCAGC-3′, and the anti-sense strand was 5′-ATGTCACGCACGATTTCCC-3′. The cDNA of the sample was taken as the template for RCR. The programe is 94°C denature for 4 min, 94°C for 3s, 55°C for 10s, 72°C for 10s. followed by 30 cycles and final extension final of 72°C for 10 min. The PCR products were detected by 1.5% agarose gel electrophoresis. Syngene gel imaging analysis system was used to the gray values of CXCR4, SDF-1 and β-actin band.
Results
Expression of AdSDF-1α/54/KDEL
HEK-293 cells were infected with AdSDF-1α/54/KDEL, and obvious cytopathic effects appeared 2 to 3 days after infection. Western blotting revealed SDF-1α/54/KDEL protein expression near 11 kDa from the first generation of packaged virus supernatant (Fig. 1A), demonstrating that the novel intrakine mutant SDF-1α/54/KDEL is effectively expressed in HEK-293 cells. The cells were collected by centrifugation, and we calculated end point titers of 6×109 pfu/mL. The MOI of amplified recombinant adenoviruses was 0–200, and its infection efficiency for MCF-7 cells was 100% when MOI was 100 after 72 h (Fig. 1B).

In vitro results following intrakine treatment.
Effects of AdSDF-1α/54/KDEL on cell proliferation and viability
Survival and proliferation of cells infected with AdSDF-1α/54/KDEL were similar to cells infected with the control intrakine (Fig. 1C, D). We found that cell numbers were reduced by 5% and 3% in MCF-7 and MDA-MB-231 cells, respectively, at an MOI of 60 when compared with uninfected cells (n=3, P<0.01). Therefore, accumulation of AdSDF-1α/54/KDEL in the ER did not significantly affect cell viability relative to the control.
Depletion of surface CXCR4 with AdSDF-1α/54/KDEL in vitro
To determine the effect of AdSDF-1α/54/KDEL on the cell surface expression of CXCR4, MCF-7 and MDA-MB-231 cells were infected with adenovirus-encoding intrakines with an MOI of 60 and analyzed by flow cytometry at 3 days after infection. For the MCF-7 and MDA-MB-231 cells infected with AdSDF-1α/54/KDEL, expression of CXCR4 was dramatically reduced by 98.6% and 92.5%, respectively (Fig. 2). These results demonstrate the remarkable effectiveness of this functional, ER-specific intrakine.
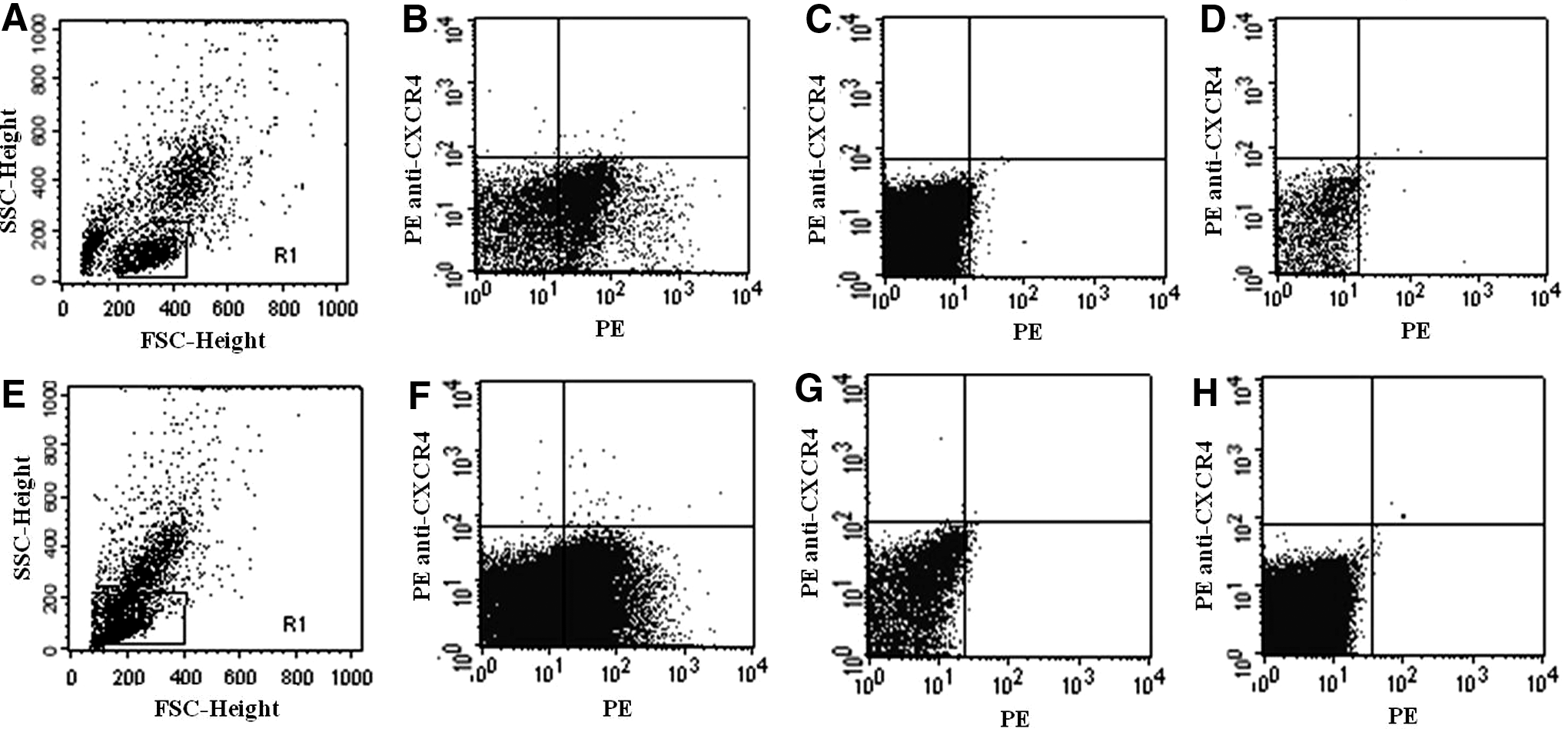
CXCR4 expression in vitro following intrakine treatment. Downregulation of CXCR4 in
AdSDF-1α/54/KDEL attenuates migration and invasion in vitro
To determine the effects of CXCR4 downregulation on migration in vitro, we performed transwell assays with MCF-7 and MDA-MB-231 cells with controls (RPMI and AdNull) and treatments (AdSDF-1α/KDEL and AdSDF-1α/54/KDEL). We observed that the intrakine mutant caused a dramatic decrease in cell migration and chemotaxis to SDF-1α (Fig. 3A, B). To determine the effects of the intrakine mutant on cell invasion in vitro, we used the aforementioned controls and treatments in MCF-7 and MDA-MB-231 cells and used Matrigel to measure the percentage of cell invasion. The intrakine mutant caused a profound reduction in cellular invasion in these 2 metastatic breast cancer cell lines (Fig. 3C, D).

Cell migration, chemotaxis to SDF-1α, and cell invasion following intrakine treatment. Downregulation of CXCR4 expression with our intrakine mutant attenuated cell migration, chemotaxis to SDF-1α, and cell invasion in
In vivo effects of AdSDF-1α/54/KDEL
Cultured MDA-MB-231 breast cancer cells were inoculated subcutaneously into BALB/c mice, and they were subsequently treated with RPMI, AdNull, AdSDF-1α/KDEL, or AdSDF-1α/54/KDEL. We found that the intrakine mutant caused a marked reduction in tumor volume and weight in vivo (Fig. 4A, B). We also observed that AdSDF-1α/54/KDEL dramatically reduced metastasis to the lung to 23% and increased survival to 81% for mice 8 weeks after treatment (Fig. 4C, D). These results demonstrate the remarkable effectiveness of this intrakine mutant in vivo on tumor growth, metastasis, and survival.
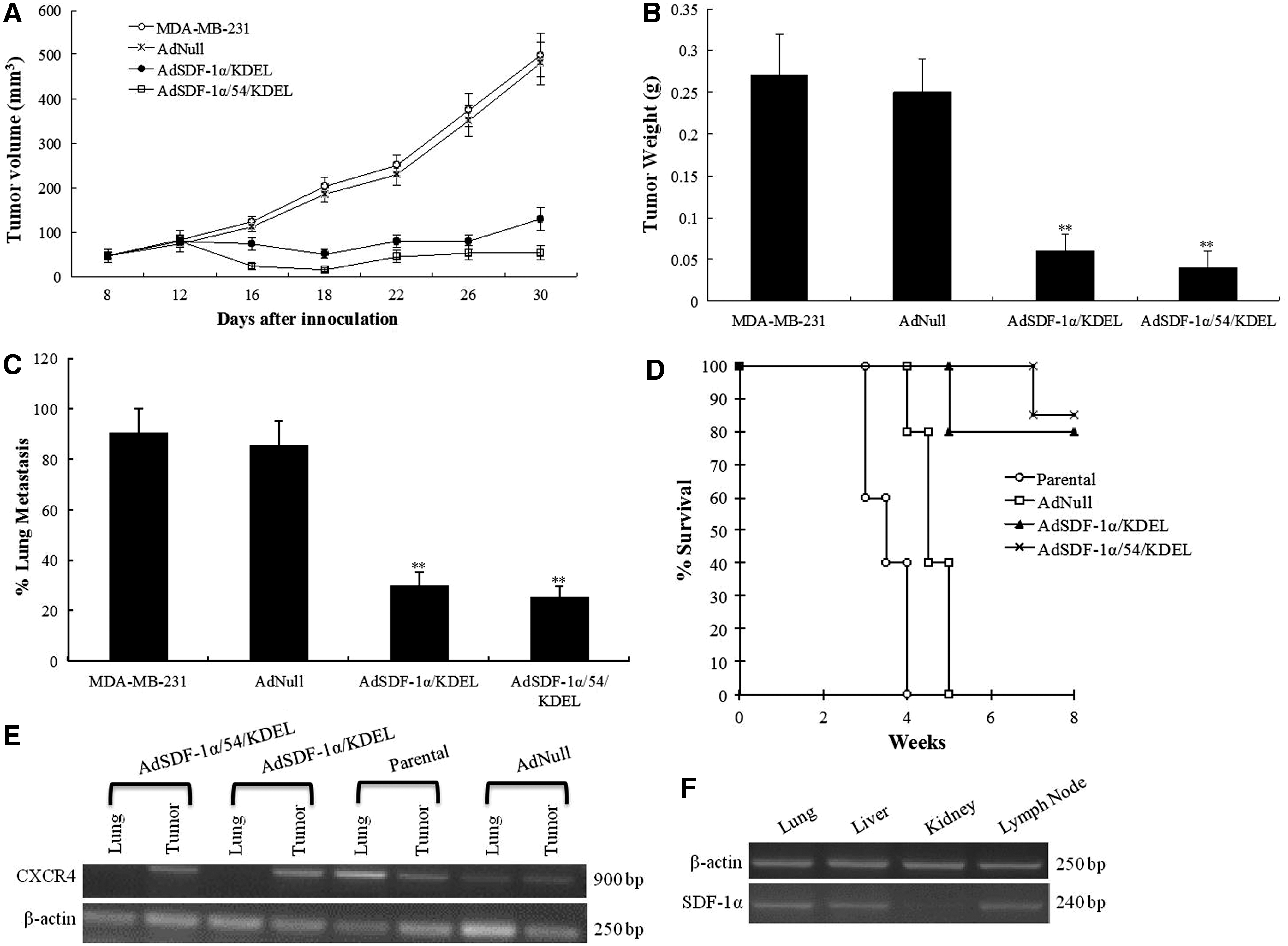
In vivo results following intrakine treatment. Our AdSDF-1α/54/KDEL intrakine mutant decreased
Primary tumors and lung metastases in tumor-bearing mice possessed high expression of CXCR4, while the mice infected with AdSDF-1α/54/KDEL possessed CXCR4 expression in tumor tissue, but not in lung tissue (Fig. 4E). We observed SDF-1α expression in the lung, liver, and lymph node, but not in the kidney (Fig. 4F). The lungs typically express CXCR4 and SDF-1α, making this organ highly susceptible to metastasis from breast cancer. We found that the intrakine mutant attenuated CXCR4 expression in the lungs, which reduced the ability of CXCR4-positive breast cancer to metastasize to the lungs. These results suggest that the lung metastasis of MDA-MB-231 cells is closely related to the high expression of SDF-1α and CXCR4. Our CXCR4 phenotype knockout strategy blocks the interaction between CXCR4 and SDF-1α and inhibits the lung metastasis of breast cancer.
Discussion
In comparison to current cancer treatment strategies, intrakine gene therapy techniques offer unique advantages for potential clinical applications due to their ability to be manipulated and their relative effectiveness and safety (Fernandis and others 2004). The only specific ligand of CXCR4 is SDF-1; however, using native SDF-1 in CXCR4 phenotypic knockout strategies may present numerous risks, such as widespread inflammatory activation, intratumoral inflammation, chemotaxis, and metastasis through its activation of CXCR4, that are avoided by using the SDF-1α/54 mutant (Fernandis and others 2004). Cancer cells also secrete chemokines to attract host cells that promote factors for angiogenesis and growth, and CXC chemokines, such as SDF-1, are known to play a pivotal role in the control of angiogenesis with CXCR4 (Chen and others 2003; Liang and others 2005). Therefore, the SDF-1α/54 mutant, which internalizes CXCR4 from the cell membrane, is a more practical component for developing a recombinant protein for CXCR4 phenotypic knockout.
We previously performed site-directed mutagenesis for SDF-1α to obtain protein mutants using the prokaryotic expression vector pET30a(+). However, the proteins functioned as antagonists to CXCR4. Removing the C-terminal α-helix, forming the SDF-1/54 mutant, eliminated the CXCR4 phenotype. This SDF-1/54 mutant functions similarly to SDF-1α, which plays a critical role in CXCR4 activation and signaling; however, the SDF-1/54 mutant induces CXCR4 internalization and decreases SDF-1α-mediated signaling and chemotaxis (Cai and others 2004; Tan and others 2006; Ma and others 2008, 2009a, 2009b). Here, the novel intrakine mutant, AdSDF-1α/54/KDEL, contains an adenovirus vector, a mutant form of SDF-1 with CXCR4 antagonism, and an ER-directing KDEL tetrapeptide sequence. These 3 components function synergistically to penetrate tumor cells and produce a phenotypic knockout of CXCR4. The adenovirus vector transports the intrakine across the cell membrane, and the SDF-1α/54 mutant induces internalization of the CXCR4 receptor. The KDEL sequence promotes the localization of our exogenous intrakine at the ER, where endogenous proteins, such as CXCR4, are synthesized, assembled, and secreted. By interacting with its receptors in the intermediate compartment and the Golgi apparatus, KDEL plays a key role in the ER retention of type I transmembrane proteins and many soluble proteins in the cisternal lumen (Yamamoto and others 2001; Wheeler and others 2003; Meijer and others 2006; Capitani and Sallese 2009; Alanen and others 2011). Our previous data revealed that SDF-1α/54/KDEL expressed by the pcDNA3.1 eukaryotic expression vector was located at the ER of the mammalian cells (Chen and others 2008). This unique localization of AdSDF-1α/54/KDEL at the ER provided us with an effective means of capturing newly synthesized CXCR4 at the ER.
It is important to consider whether treatment with SDF-1α/54/KDEL in vivo could cause certain side effects due to adenovirus-mediated nonspecific cell penetration as well as CXCR4 expression in noncancer cells; however, we did not observe any noticeable toxic effects in the heart, spleen, or kidney of mice injected with AdSDF-1α/54/KDEL at primary tumor sites (Smith and others 2004). Thus, to be as safe and effective as possible for future clinical applications, we need to optimize the targeting of SDF-1α/54/KDEL to selectively penetrate tumor cells. One potential method to increase the selective targeting of our intrakine mutant to tumor cells is the use of a folate-conjugated liposome complex. Most tumor cell types possess high specific expression of folate receptor, and our previous research has shown that using a folate-conjugated liposome complex is highly effective in selectively targeting breast cancer cells (Chen and others 2010). Therefore, folate-conjugated liposome complexes may offer a highly effective method for optimizing the targeting and selectivity of the intrakine to tumor cells.
The SDF-1/CXCR4 axis has been shown to be instrumental in tumor growth (Scotton and others 2002; Smith and others 2004; Kollmar and others 2007; Marlow and others 2008; Kollmar and others 2010). Downregulation of CXCR4 expression through negative regulators of SDF-1 and CXCR4, such as Slits, has been shown to cause a dramatic suppression of tumor growth in breast cancer (Marlow and others 2008). Our research revealed that the novel intrakine mutant, AdSDF-1α/54/KDEL, is effectively transfected and becomes localized in the ER, blocking the transport of newly synthesized CXCR4 to the cell membrane of breast cancer cells. These components function synergistically to downregulate CXCR4 expression in breast cancer cells and attenuate migration, chemotaxis, and cell invasion in vitro. We observed that our intrakine mutant also exhibited remarkable effectiveness in vivo in downregulating CXCR4 expression to cause a dramatic decrease in tumor volume, weight, metastasis and an increased survival in BALB/c mice. Thus, the novel intrakine mutant inhibits tumor metastasis mediated by the CXCR4/SDF-1 axis. Further research needs to be conducted with regard to the intrakine's in vivo effects and its role in the signal transduction pathway for breast tumor growth. It will also be prudent to investigate possible cytokine induction, such as IFN and other inflammatory factors, following intrakine injection. Further studies are required to optimize and develop this treatment strategy for clinical applications and to selectively target tumor cells; however, combination therapies with this intrakine strategy and classic cancer treatments may offer unique opportunities for enhanced cancer treatment outcomes in the near future.
Footnotes
Acknowledgments
This research was prompted and supported by the 54th China Postdoctoral Science Foundation No. 2013M542234, the Science and Technology Project of Guangdong Province, China (No. 2013B021800085), and the Science and Technology Project Guangzhou City Grant (No. 2014J4100058). We would also like to thank Professors Jun Du and Shao-Hui Cai for their excellent recommendations and edits with this manuscript.
Author Disclosure Statement
The authors declare that there are no conflicts of interest.