
Abstract
Interleukin-12 (IL-12), a potent inducer of interferon gamma (IFNγ), is a heterodimeric protein consisting of p40 and p35 subunits whose expression is regulated independently. IL-12 is part of a cytokine family (currently consisting of IL-12, IL-23, IL-27, and IL-35) that can have profoundly different immunologic effects, despite sharing subunits. In constructing a single-chain fusion of p40 and p35, we discovered an insert corresponding to an intron in the gene encoding the p35 subunit that would result in a truncated form of p35 if translated. To test its possible role, we constructed, expressed, and analyzed fusions of p40 with the full-length or the truncated form of p35. The fusion protein containing the truncated p35 did not stimulate the proliferation of the IL-12-responsive cell line CTLL-2 nor did it induce IFNγ or the chemokine IFNγ-inducible protein 10 (IP-10, CXCL10) or monokine induced by IFNγ (MIG, CXCL9) from spleen cells. In striking contrast, the full-length IL-12 p40/p35 fusion induced robust responses in both assays. Moreover, the truncated IL-12 fusion protein inhibited the action of the full-length IL-12 p40/p35 fusion in the proliferation assay and also blocked the induction of IFNγ. These findings raise the possibility that alternative splicing may provide an additional regulatory mechanism for IL-12.
Introduction
I
IL-12 and other related members of its family, such as IL-23, IL-27, and IL-35, are unusual cytokines since they are heterodimeric molecules that can share subunits with other members of the family (Vignali and Kuchroo 2012). The receptors for these cytokines show similar complexity, with some receptors being shared among family members, whereas others are unique (Vignali and Kuchroo 2012; Ngiow and others 2013). Various combinations of these family members can be inflammatory and enhance immune responses (eg, IL-12), and in some cases, promote autoimmune responses (eg, IL-23) or be immunosuppressive (eg, IL-35) and downregulate immune responses (Trinchieri and others 2003; Vignali and Kuchroo 2012; Ngiow and others 2013). The function of these powerful cytokines thus needs to be carefully controlled. The p40 and p35 subunits of IL-12 are independently regulated and are often not produced stoichiometrically with respect to one another (Cooper and Khader 2007).
The p40 protein may have additional functions independent of other subunits (Gillessen and others 1995; Abdi 2002). Also, the p40 subunit can exist as a monomer or as a homodimer when expressed at high levels, and these forms may have different and even opposing functional activities (Gillessen and others 1995; Abdi 2002). Recently, it has been proposed that the p40 subunit might also be able to complex with partners outside the cell (Abdi and others 2014) and, as a consequence, might have yet undiscovered and even opposing functions, adding additional complexity to the biological action of this cytokine family.
We initially set out to construct a fusion protein of p40 and p35 to have a defined reagent that minimizes these potentially confounding aspects of its function. By constructing a single-chain fusion, the subunits are automatically produced in stoichiometrically equivalent amounts. Moreover, their biochemical linkage in a fusion protein also ensures p40 and p35 bind to each other and not to other family members. In the course of assembling a DNA construct encoding a single polypeptide fusion consisting of p40 connected to p35, we unexpectedly found an additional sequence in the p35 component. Further analyses showed that the unexpected sequence corresponded to an intron, which could result from alternative mRNA splicing. The in-frame translation of this intron would ultimately result in a truncated version of the p35 subunit of IL-12. The current work used a fusion containing the putative truncated p35 component and compared it with a p40/p35 fusion containing the complete p35 sequence to investigate the properties of this molecule. In this study, we show that it has the potential to block 2 critical functions of IL-12.
Materials and Methods
Construction of murine IL-12 single-chain fusion proteins
For our initial studies, the murine p40 and p35 subunits were polymerase chain reaction (PCR) amplified from the pWRG3169 plasmid (Rakhmilevich and others 1996; Tan and others 1996) using the following primers: p40 forward 5′-CCGTCCCACCATCGGGCGCGGATCAGATCTATGT GTCCTCAGAAGCTAACCATCTCCTGG-3′, p40 reverse 5′-GAGGAACGCACCTTTCTGGTTACACCCCTC-3′, p35 forward 5′-GAAGACATCGATCATGAAGACATCACA CGG-3′, and p35 reverse 5′-GGAAAGGATCCCGGGTAC CTTCTAGAATTCTCATTAGTGGTGGTGGTGGTGGTG GGCGGAGCTCAGATAGCCCAT-3′. The geneblock DNA fragment (corresponding to the carboxy-terminal region of p40, the repeating glycine/serine linker, and the sequence corresponding to the N-terminal region of p35) was purchased from Integrated DNA Technologies (IDT). The Gibson reaction (Gibson and others 2009) was performed to knit together the 4 individual fragments [p40, geneblock (linker), p35, and the linearized pVL1392 vector]. This single-chain IL-12 construct was sequenced and found to have an insert that corresponds to an intronic region (intron 4) of p35.
Using PCR and the Gibson reaction, the intronic region was removed and a single-chain IL-12 consisting of the p40 linked to full-length p35 was assembled in the pVL1392 vector with the addition of a carboxy-terminus 6x Histidine (His) tag. To construct a single chain of p40 linked to truncated p35 with a 6x-His tag, PCR amplification of the full-length IL-12 single chain was performed using the following primers: p40 forward (listed above) and truncated reverse 5′-AAGGGTTCCTTCCCCATGACGGATTCACA CCACCACCACCACCACTAATGAGAATTCTAGAAGG TACCCGGG-3′. The p40/p35 truncated IL-12 product was then directionally cloned into the pVL1392 plasmid vector using the BglII/EcoRI restriction sites.
Baculovirus production of IL-12 fusion proteins
The generation of recombinant baculoviruses for the expression of proteins in insect cells was performed as described previously (Rose and others 1990, 1994; Puskas and others 2011). Recombinant viruses were created using the pVL1392 transfer vector and the BD BaculoGold transfer vector system (BD Biosciences) basically as described by the manufacturer. Initial virus production was performed in Spodoptera frugiperda (Sf-9) cells cultured in Sf-900 II serum-free media (SFM; Gibco). After 2 passages (P0 and P1) of Sf-9, a high-titer stock was obtained (P1). For final protein production, Trichoplusia ni (Tni) cells were cultured in Express Five SFM (Gibco) plus 2 mM
Western blotting
Immunoblot analysis was performed on the full-length IL-12 and the truncated IL-12 as described previously with minor modifications (Puskas and others 2011). A monoclonal penta-His primary antibody (Qiagen) was used followed by detection with a goat anti-mouse HRP-conjugated secondary antibody (Jackson Immuno Research) and developed using the Amersham ECL Plus Western blotting detection reagent (GE Healthcare) as per the manufacturer's instructions.
IL-12 functional assay
Functional IL-12 was measured using CTLL-2 cells (ATCC) as described with minor modifications (Khatri and others 2007; Puskas and others 2011). In brief, Ni-NTA-purified proteins were serially diluted 1:5 in 50 μL of medium, then 4×104 CTLL-2 cells in 100 μL of medium were added per well to a 96-well plate and incubated at 37°C in 5% CO2 for 18–22 h. At the end of this period, 75 μg/well of Thiazolyl Blue Tetrazolium Bromide (MTT; Sigma-Aldrich) was added and the plate was incubated for 8 h at 37°C in 5% CO2. Cells were lysed with 100 μL/well of 10% SDS (Gibco) acidified with HCl, incubated at 37°C in 5% CO2 overnight, and absorbance was read at 570 nm. For the competition assay, we again used the CTLL-2 assay with minor modifications. In brief, full-length IL-12 was serially diluted 1:5 in 80 μL of medium alone or with the addition of a constant amount of truncated IL-12 per well in the 96-well plate, then 4×104 CTLL-2 were added per well, and the plate was developed as indicated above.
Cytokine production by spleen cell cultures
BALB/c spleen cells were plated at 5×106 per well in a 24-well plate. Equal molar amounts of either full-length IL-12 or truncated IL-12 were added to the cultures, which were then incubated at 37°C in 5% CO2 for 24 h. Supernatants were collected and assayed for cytokine expression (IFNγ, MIG, IP-10) using the Luminex assay (Millipore) according to the manufacturer's instructions. For the continuous culture assay, the full-length IL-12 was serially diluted 1:2 in 50 μL of medium alone to obtain a maximum signal or with the addition of a constant amount of truncated mIL-12 per well in the 96-well plate. Spleen cells were added at 1×106 per well and the plate was then incubated at 37°C in 5% CO2 for 24 h. Supernatants were collected and an IFNγ enzyme-linked immunosorbent assay (ELISA; eBioscience) was performed as per the manufacturer's recommendations. For the pulse assay, spleen cells were pulsed for 10 min with serial dilutions of IL-12 alone or in the presence of a constant concentration of truncated IL-12 followed by extensive washing. Supernatants were collected at 24 h and analyzed for IFNγ production by ELISA.
Results
Identification of a novel form of the p35 subunit of IL-12
In the course of constructing a p40/p35 fusion protein, we discovered that the p35 component, which had been originally derived from a cDNA library (Rakhmilevich and others 1996), had an apparent insert. A blast search of GenBank revealed that this insert was 100% homologous to intron 4 in the gene encoding the p35 subunit (Fig. 1) suggesting that this form might be derived from an alternatively spliced variant of p35. In-frame translation of this sequence would result in a truncated form of the p35 subunit due to the presence of a stop codon in the retained intron (Fig. 1). To characterize this novel form of p35 further, we created a fusion protein of p40 joined to the truncated p35 molecule with the carboxy-terminal 6x-His tag (Fig. 2). Using such a fusion reduces potential complexities due to the unequal production of the different subunits. This ensures that the p35 subunit will pair exclusively with the p40 subunit by being physically connected in the same molecule. This construct and the corresponding p40/p35 full-length fusion were created using a combination of traditional cloning techniques and the Gibson reaction (Gibson and others 2009) and were subsequently expressed using the baculovirus system and purified by affinity chromatography through the 6x-His tag using the NTA agarose. This purified material was examined by Western blot analysis using an anti-6x-His detecting antibody. As expected, the IL-12 fusion protein containing the truncated form of p35 had a somewhat lower apparent molecular weight than the full-length IL-12 fusion protein (Fig. 2), indicating that we had successfully constructed and expressed a single-chain fusion protein containing these alternate forms of the p35 subunit.
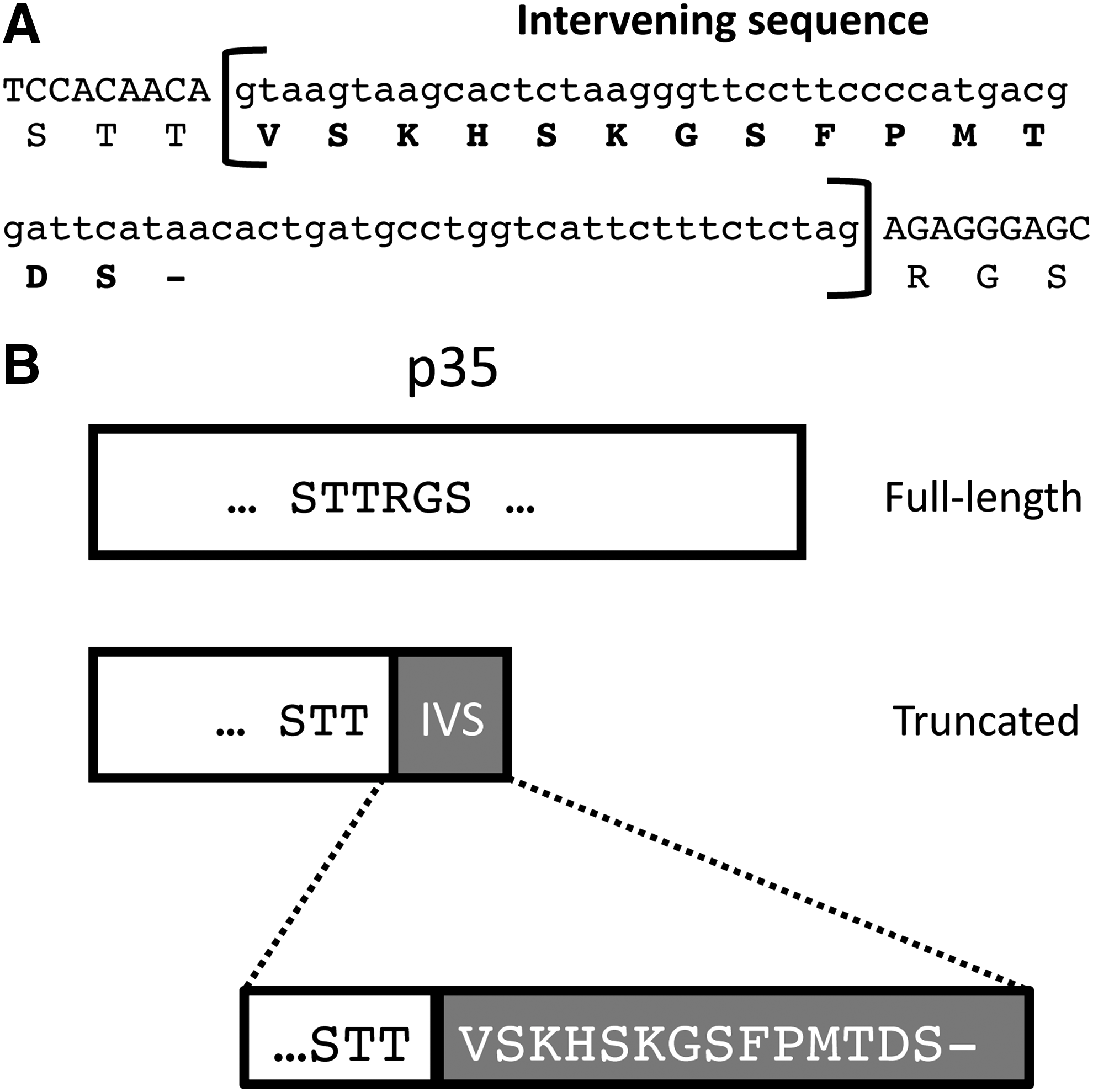
Intervening sequence (IVS) identified in the p35 subunit.

Interleukin (IL)-12 full-length and truncated proteins are produced in Tni insect cells.
A fusion protein encoding the p40/p35 truncated form of IL-12 is not stimulatory
As a first test of the functional properties, we examined the ability of the fusion proteins to stimulate the IL-12-responsive cell line, CTLL-2. While this cell line, which was derived from a cytotoxic cell line, is best known for its requirement and ability to respond to IL-2 (Gillis and Smith 1977; Baker and others 1979), it can also respond to IL-12 (Khatri and others, 2007; our unpublished data). As shown in Fig. 3A, the full-length p40/p35 fusion protein stimulates the proliferation of this cell line. These data indicate that the fusion protein consisting of p40 fused to p35 was functional. In contrast, the p40/p35 truncated form of IL-12 did not result in detectable proliferation of these cells at any concentration tested (Fig. 3A). Another major function of IL-12 is induction of IFNγ production by T cells and NK cells. We therefore tested the ability of the full-length p40/p35 fusion or the p40/p35 truncated form to induce IFNγ from spleen cell cultures. As can be seen in Fig. 3B, the full-length p40/p35 fusion results in the dramatic stimulation of IFNγ, whereas the p40/p35 truncated form did not. Consistent with this finding, the truncated fusion protein also did not stimulate IP-10 (CXCL10) or MIG (CXCL9) (Fig. 3C, D), which are induced by IFNγ.

Truncated IL-12 does not show the functional activity compared with the full-length IL-12.
The p40/p35 truncated form blocks the ability of the p40/p35 full-length fusion protein to stimulate CTLL cell line
The above data showed that the truncated IL-12 fusion protein was not functional in a standard proliferation assay using an IL-12-responsive cell line. However, although the truncated IL-12 was not stimulatory, we hypothesized that it might bind to the IL-12 receptor but not to signal effectively. A prediction of such a model is that the p40/p35 truncated form might serve as an antagonist with respect to the stimulatory ability of the full-length IL-12 fusion protein. To test this concept, we performed a competition experiment in which we titrated the amount of full-length fusion protein, in the presence or absence of a fixed amount of IL-12 truncated fusion protein. As can be seen in Fig. 4, the IL-12 truncated fusion protein inhibits the ability of the full-length IL-12 fusion protein to stimulate the CTLL cell line. The stimulatory ability of the full-length IL-12 fusion protein is inhibited ∼20- to 30-fold under these conditions.

Truncated IL-12 blocks the ability of the full-length IL-12 to stimulate CTLL-2 cell line.
The IL-12 p40/p35 truncated fusion protein inhibits the IFNγ response of spleen cells by full-length IL-12
To further characterize the potential for the IL-12 truncated form to act as an antagonist, we examined the ability of the p40/p35 truncated form to inhibit the production of IFNγ by spleen cells when incubated with the full-length IL-12 fusion protein. The full-length IL-12 was diluted serially by itself or in the presence of a fixed amount of truncated IL-12, and the amount of IFNγ is determined by ELISA. As shown in Fig. 5A, the action of p40/p35 full-length IL-12 is inhibited in the presence of p40/p35 truncated protein. To minimize potential complexities inherent in the continuous culture method and to obtain a more quantitative analysis that is likely more directly related to IL-12 binding, we used a short-term pulse assay. Spleen cells were pulsed for 10 min with varying concentrations of full-length IL-12 in the absence or presence of a set of concentrations of truncated IL-12. The cells were extensively washed and the amount of IFNγ at 24 h was then determined by ELISA. As shown in Fig. 5B, under the conditions of this assay, the concentration of full-length IL-12 required to achieve 50% maximum signal increased by 33-fold in the presence of the p40/p35 truncated protein (3 pM versus 100 pM). Thus, the p40/p35 truncated form not only is not stimulatory for cytokine secretion on its own but also appears that it can competitively inhibit the cytokine-inducing activity of full-length IL-12.
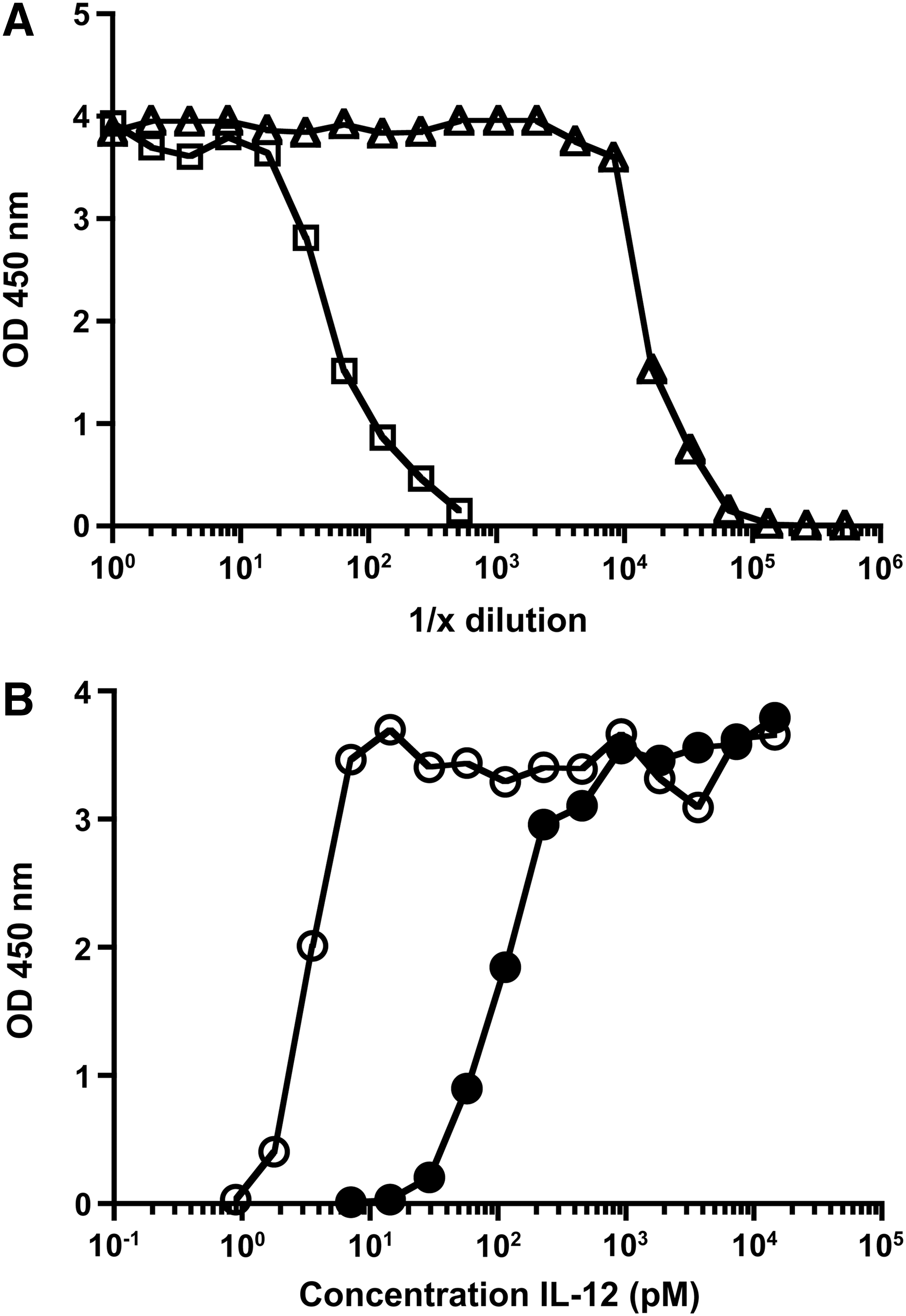
Truncated IL-12 competitively inhibits IFNγ production by the full-length IL-12 in spleen cultures.
Discussion
An IL-12 fusion protein containing a truncated version of p35 has unexpected function
We have shown that a fusion protein consisting of the p40 subunit and a truncated version of p35 exhibits some noteworthy properties. Our results using a proliferation assay show that the p40/p35 truncated fusion protein does not stimulate CTLL-2, an IL-12-responsive cell line. It also does not engender the production of IFNγ, IP-10, or MIG by spleen cells. Importantly, it appears to act as an antagonist in that it inhibits the action of full-length IL-12. Our data showed that truncated IL-12 not only inhibits proliferation of the cytokine-dependent cell line CTLL-2 but also blocks IFNγ production normally generated in spleen cell cultures by full-length IL-12. This inhibition is consistent with previous structural and mutational analyses of IL-12. Prior work using chimeric molecules, constructed from portions of human and mouse p35, took advantage of the species specificity that is dependent on the p35 subunit to suggest that binding could be differentially uncoupled from signaling (Zou and others 1995). Using chimeric mouse–human molecules as well as combinatorial mutational approaches, it was reported that 5 amino acids (K57, H58, S60, H146, and P184) in p35 were important for IL-12 signaling in mouse cells (Zou and others 1995). The truncated version of p35 lacks residues H146 and P184, yet retains amino acids K57, H58, and S60 and thus might be expected to have limited signaling potential consistent with the results shown here.
The truncated p35 subunit could arise from alternative RNA splicing
Our initial analysis of the unexpected form of p35 suggested that the truncated form could be the result of an alternatively spliced mRNA such that an intron was not removed. This alternatively spliced variant would result in a truncated version of p35 due to an in-frame stop codon. Our search of GenBank revealed that this insert corresponded to an intron and that it was a predicted isoform of the p35 subunit (ENSEMBL Transcript IL-12A-001 ENST00000305579). Blast analysis also revealed that a cDNA corresponding to the form of p35 employed in this article was found independently in a cDNA library from B10.SJL mice (GenBank accession number AF128219). Additionally, the p35 component that we utilized was originally derived from a cDNA library of mouse lymphocytes (Rakhmilevich and others 1996), also consistent with the idea that it is a relevant alternatively spliced form of p35. Interestingly, the p35 containing the intron was present in a mammalian expression vector that produces functional IL-12 (Rakhmilevich and others 1996; Tan and others 1996) presumably by preferentially splicing out the intervening intronic sequence. It is likely that it was initially unrecognized due to the small size of the intron and the fact that the plasmid expressed functional IL-12 after transfection (Rakhmilevich and others 1996; Tan and others 1996; Moran and others 2003). Further work is needed to establish how the splicing is regulated and to definitively determine the biological significance of the truncated p35 subunit of IL-12 in vivo. However, our observations raise the intriguing possibility that it may have a biological role either alone or by interacting with either p40 or Epstein–Barr virus induced gene 3 (EBI3) (affecting IL-12 or IL-35, respectively) or even possibly interact with additional yet unidentified proteins.
Importantly, it is now being appreciated that the “transcriptome” that can arise from alternative splicing is an important means of increasing the diversity from a finite number of genetic elements and can play an important role in almost all cellular processes (Kelemen and others 2013). One of the best-characterized examples in immunology is immunoglobulin M (IgM), in which alternative splicing can convert IgM from a membrane-bound signaling molecule to a secreted effector molecule. Alternative splicing that has functional significance has also been reported for cytokines, such as IL-4 (Arinobu and others 1999; Luzina and others 2012). Interestingly, an alternatively spliced form of the human growth hormone-releasing hormone receptor that results in intron retention generates a dominant-negative form of the receptor (McElvaine and Mayo 2006), thereby inhibiting signaling in this pathway. Moreover, intron retention in the gastric inhibitory polypeptide receptor also results in a truncated protein with a dominant-negative effect (Harada and others 2008). Thus, there is a precedent for alternative splicing and intron retention also being used to block important cellular pathways.
The truncated version of p35 might provide another level of IL-12 regulation
It is becoming increasingly evident that IL-12 and other family members of this axis (IL-23, IL-27, and IL-35) with shared and unique subunits is one of the most complex cytokine families. It is not only structurally more complex than other cytokine families (Wang and others 2009; Ngiow and others 2013) but also the actions of these cytokines can be dramatically different and result in robust inflammation or immune suppression depending on the family member and the exact combination of subunits (Vignali and Kuchroo 2012). The precise control of the expression of these components is therefore essential. It is intriguing to speculate that this alternate form of p35 might give yet another level of control to this cytokine family. For example, alternative mRNA splicing of p35 could create a rapid means of attenuating immune responses even in the presence of already existing IL-12. Interestingly, alternatively spliced forms of p19, which like p35 can pair with p40, have been described, which would also result in a truncated protein (Waibler and others 2007; Sender and others 2010). However, the precise roles of the p19 or p35 splice variants are not yet fully understood and indeed may even have yet undiscovered biological functions.
Several lines of evidence indicate that the precise control of IL-12 is critical. As one might predict given its important role, deletions affecting the cytokine or its receptor can have significant immunological effects (Altare and others 1998a, 1998b; Decken and others 1998; Cooper and others 2002; Lichtenauer-Kaligis and others 2003; MacLennan and others 2004; Rosenzweig and Holland 2005). However, even more subtle changes in this system can have important effects. For example, a polymorphism in the 5′ flanking region of the IL-12 receptor β2 subunit in humans appears to reduce its level of expression and is correlated with clinical types of leprosy (tuberculoid versus lepromatous) manifested in patients (Ohyama and others 2005). Other polymorphisms in the untranslated regions of IL-12 receptor β1 or in noncoding regions of the IL12B gene encoding p40 result in the increased susceptibility to tuberculosis in certain ethnic groups (reviewed in Azad and others, 2012). While outside the scope of the current work, it will be crucial in the future studies to determine when and under what circumstances this putative form of p35 is expressed. Importantly, our results show that this molecule when in complex with p40 has the potential to act as an antagonist to IL-12. This single-chain form of truncated IL-12 may also be a useful experimental reagent and could even be useful as a potential therapeutic.
Footnotes
Acknowledgments
The authors thank Dr. Abigail Sedlacek for helpful comments to the article. We also acknowledge generous support from Steven and Alison Krausz and from F.C. Blodgett. D.S. was supported in part by the National Institutes of Health Training Grant AI007285. S.M. was supported by GM064133 from the National Institutes of Health. The project described in this publication was supported in part by the University of Rochester CTSA award number UL1 TR000042 from the National Center for Advancing Translational Sciences of the National Institutes of Health. The content is solely the responsibility of the authors and does not necessarily represent the official views of the National Institutes of Health.
Author Disclosure Statement
No competing financial interests exist.