
Abstract
The cationic antimicrobial peptide, LL37, forms electrostatic complexes with DNA (LL37-DNA), which are potent activators of circulating plasmacytoid predendritic cells (ppDCs) and monocytes. However, the effects of LL37-DNA on other immune cell types, such as NK cells, are poorly characterized. In this study, we show that complexes of human genomic DNA (hgDNA) or synthetic double-stranded oligodeoxynucleotides with LL37 strongly enhance natural cytotoxicity of human peripheral blood mononuclear cells (PBMCs) upon an overnight culture, whereas hgDNA alone has no effect, and LL37 alone is moderately active. LL37-DNA complexes potentiate degranulation of, and interferon (IFN)-γ production by, NK cells upon subsequent encounter of K562 target cells. The complexes do not influence percentages of NK cells among PBMCs or the expression of cytotoxic proteins by NK cells. Using neutralizing anticytokine antibodies and immunomagnetic depletion of different subpopulations of PBMCs, we found that the effect of LL37-DNA on NK cells is indirect and mediated by type I IFNs produced by monocytes and, to a lesser extent, by ppDCs. We discuss possible roles of LL37-DNA complexes in the regulation of NK cell functions and in the treatment of cancer.
Introduction
L
LL37 electrostatically interacts with eukaryotic, bacterial, or viral DNA and RNA, as well as with synthetic CpG-oligodeoxynucleotides (CpG-ODNs) and synthetic double-stranded RNA (poly[I:C]). The resulting complexes are immunostimulatory; in particular, they induce the production of interferon (IFN)-α, tumor necrosis factor (TNF), and interleukin (IL)-6 by innate immune cells (Lande and others 2007; Chuang and others 2009; Ganguly and others 2009; Lai and others 2011; Singh and others 2013). Within these complexes, LL37 protects DNA or RNA from endonucleases and facilitates the delivery of nucleic acids into the endosomal and cytosolic compartments (Lande and others 2007, 2011; Ganguly and others 2009; Chamilos and others 2012).
The complexes of LL37 and DNA (LL37-DNA) target primarily monocytes and plasmacytoid predendritic cells (ppDCs) (Lande and others 2007; Chamilos and others 2012). Importantly, monocytes take up LL37-DNA much more efficiently than ppDCs (Chamilos and others 2012). In both cell types, LL37-DNA first accumulate in the endosomes. In monocytes, LL37-DNA subsequently translocate through the endosomal membrane and in 24 h are found diffusely in the cytosol. In ppDCs, on the contrary, LL37-DNA are contained within the early endosomes (Chamilos and others 2012). Virtually no uptake of LL37-DNA is observed in NK cells, T cells, B cells, keratinocytes, or fibroblasts (Chamilos and others 2012).
In ppDCs, LL37-DNA activate the endosomal receptor toll-like receptor 9 (TLR9), which triggers the production of type I IFNs (Lande and others 2007). In monocytes, LL37-DNA activate cytosolic DNA sensor(s) that signal through the stimulator of interferon genes (STING) and also induce the production of type I IFNs, although the intensity and kinetics of IFN expression in monocytes differ from those in ppDCs (Chamilos and others 2012).
The emergence of LL37-DNA complexes in healthy individuals has not been directly demonstrated. However, it is clear that such complexes are present in pathological conditions, eg, in the psoriatic skin in sera from patients with systemic lupus erythematosus (Lande and others 2007, 2011). Auto-DNA complexed with LL37 is a component of neutrophil extracellular traps, which are a product of neutrophil activation in different inflammatory and infectious diseases (Brinkmann and others 2004; Lande and others 2011).
Altogether, these data indicate that LL37-DNA complexes probably play an important role in the regulation of immune processes in the body. However, little is known about the effects of LL37-DNA on cells other than monocytes and ppDCs, eg, on NK cells. A first step in this direction was made by Chuang and others (2009), who showed that complexes of CpG-ODNs with LL37 are more potent activators of murine NK cells than CpG-ODNs alone. However, mechanisms of action of LL37-DNA on NK cells have not been examined.
Materials and Methods
Donors
Heparinized venous blood was obtained from healthy male and female donors aged from 20 to 60 years. The study was approved by the local Ethics Committee, and informed consent was obtained from all subjects.
Reagents
The peptide, LL37 (LLGDFFRKSKEKIGKEFKRIVQRIKDFLRNLVPRTES) (Lande and others 2007), was synthesized at the Institute of Bio-organic Chemistry (Moscow, Russia) using a standard solid-phase protocol (Wellings and Atherton 1997). The molecular weight (m.w.) of the peptide was verified using MALDI-TOF (calculated m.w., 4490.57; experimental m.w., 4491.69).
Human genomic DNA (hgDNA) was isolated from pooled peripheral blood mononuclear cells (PBMCs) of healthy donors using TriReagent (Sigma, St-Louis, MO) according to the manufacturer's instructions. Pairs of single-stranded complementary ODN were synthesized at Syntol (Moscow, Russia) (Table 1). Pair 1, with all cytosines 5-methylated, corresponded to a randomly selected region of human chromosome 1 (base pairs from 7460666 to 7460695). To obtain double-stranded ODNs (dsODNs), pairs of single-stranded ODNs were annealed in phosphate-buffered saline (PBS) by heating to 95°C and slowly cooling to room temperature. To prepare LL37-DNA, 10 μg hgDNA or synthetic dsODNs were mixed with 50 μg LL37 in a total volume of 60 μL PBS and incubated during a period of 30 min at room temperature (Lande and others 2007); the complexes were always used immediately and not stored. Unless otherwise specified, the term “LL37-DNA” refers to the complex of LL37 and hgDNA.
All cytosine residues are 5-methylated.
ODNs, oligodeoxynucleotides.
Recombinant human IFN-α2b was from Bioprocess (Moscow, Russia). Polyclonal goat neutralizing antibodies against IL-1α, IL-1β, and TNF, as well as normal goat IgG were purchased from R&D Systems (Minneapolis, MN); polyclonal neutralizing antisera against IFN-α and IFN-β were from Life Technologies (Paisley, UK).
Cell isolation and culture
PBMCs were isolated from venous blood by density gradient centrifugation. NK cells were isolated from PBMC suspensions using an NK cell-negative isolation kit in combination with a MidiMACS magnet and LS columns (all from Miltenyi Biotech, Bergisch Gladbach, Germany). Immunomagnetic depletion of NK cells from PBMC suspensions was done using anti-CD56 microbeads (Miltenyi Biotech). To immunomagnetically deplete CD14+ monocytes or ppDCs, suspensions of PBMCs were first labeled with fluorescein isothiocyanate (FITC)-conjugated mAbs against CD14 (Beckman Coulter, Brea, CA) or blood dendritic cell antigen (BDCA-2; Miltenyi Biotech) and subsequently incubated with anti-FITC microbeads (Miltenyi Biotech), whereafter they were passed through LS columns. Control aliquots of PBMCs were subjected to the same manipulations, except that a primary FITC-labeled mAb was omitted (mock depletion). In some experiments, positively selected CD14+ monocytes were also used.
PBMCs or their fractions were adjusted to 2×106 per 1 mL in complete culture medium [RPMI-1640 (PAA, Pasching, Austria) supplemented with 2 mM
Natural cytotoxicity assay
The assay was carried out as described (Murugin and others 2011). Briefly, K562 target cells were labeled with carboxyfluorescein succinimidyl ester (CFSE) and dispensed in 96-well plates at 8×103 per well. Effector cells (PBMCs or their fractions) were added so as to obtain E:T ratios ranging from 3:1 to 50:1. Control wells received the medium instead of effector cells. Plates were spun down (200 g, 1 min) and incubated at 37°C for 4 h, whereafter cells were resuspended in the staining buffer [PBS with 1 μg/mL propidium iodide (PI, Sigma)] and analyzed using a Cytomics FC500 flow cytometer equipped with CXP software (all from Beckman Coulter). Dead target cells were identified as CFSE+PI++ events. Specific killing and the amount of LU20 per 105 PBMCs were calculated as described (Murugin and others 2011).
NK cell degranulation assay
Degranulation of NK cells was measured by a flow cytometric assay based on externalization of CD107a after coculture with K562 target cells (Alter and others 2004; Murugin and others 2011). Briefly, cultured PBMCs or their fractions (5×105) were mixed with an equal number of K562 cells in the presence of a PC5-labeled mAb against CD107a (BD Biosciences, San Jose, CA). From each cell sample, a negative control well was prepared in parallel, which received the medium instead of K562 cells (spontaneous degranulation). After a 4-h incubation at 37°C, cells were treated with PBS containing 0.02% EDTA and 0.02% sodium azide to disrupt cell–cell aggregates and stained with ECD-anti-CD3 and PC7-anti-CD56 mAbs (both from Beckman Coulter). Externalization of CD107a (ie, percentages of CD107a+ events within the CD3−CD56+ NK cell population) was determined by Cytomics FC500. All results were expressed as ΔCD107a, ie, the difference between K562-triggered and spontaneous externalization of CD107a in the given sample.
In case of simultaneous detection of degranulation and IFN-γ, the 4-h coincubation with K562 was done in the presence of monensin (10 μM, Sigma). After surface staining with anti-CD3 and anti-CD56, cells were fixed in 4% paraformaldehyde (Sigma), permeabilized with 0.1% saponin (Sigma), and stained intracellularly with an FITC-labeled mAb against IFN-γ (Life Technologies) or with a respective isotype control. Results were expressed as percentages of CD107a+ and/or IFN-γ+ events among the gated CD56+CD3− NK cells.
Immunofluorescent detection of perforin and granzymes in NK cells
PBMCs were surface stained, fixed, and permeabilized as described above and stained intracellularly with phycoerythrin (PE)-anti-perforin (eBioscience, San Diego, CA), PE-anti-granzyme A (R&D Systems), or FITC-anti-granzyme B (BD Biosciences) mAbs. Cells were analyzed using Cytomics FC500. Levels of intracellular molecules were expressed as MFI of CD3−CD56+ NK cells in the respective channel (FITC or PE).
Enzyme-linked immunosorbent assay
Levels of TNF, IL-1β, IFN-α, and IL-6 in cell culture supernatants were measured by sandwich ELISA using reagent kits from Biosource/Life Technologies; the human IFN-β ELISA kit was from USCN Life Sciences (Wuhan, China).
Real-time polymerase chain reaction
Total RNA was isolated from TriReagent lysates according to the manufacturer's instructions and treated with RNase-free DNase I (Fermentas, Vilnius, Lithuania) to remove traces of DNA; 0.5 μg total RNA was reverse transcribed using the Revertaid reagent kit (Fermentas). Real-time polymerase chain reaction (RT-PCR) was performed using the SYBR Green approach under the following conditions: initial denaturation at 95°C, 5 min, followed by 40 cycles consisting of 15 s at 95°C and 45 s at 60°C. The following primers were used: IFNB forward, 5′-AGCAGTCTGCACCTGAAAAG-3′; IFNB reverse, 5′-CTGACTATGGTCCAGGCACAG-3′; GAPDH forward, 5′-CAGCCTCCCGCTTCGCTCTC-3′; GAPDH reverse, 5′-ACCAGGCGCCCAATACGACC-3′. All PCR reagents were from Syntol. Cycles of threshold (Ct) were determined in each reaction. The specificity of amplifications was confirmed by analyzing melting curves in each sample. Relative expression of IFNB mRNA was calculated according to the 2–ΔΔCt method, where unstimulated PBMC samples from each donor served as reference samples, and GAPDH (a housekeeping gene) was used to normalize IFNB expression.
Statistics
Multiple comparisons were done using analysis of variance (ANOVA) with Bonferroni correction. Paired comparisons were performed using paired t-test. Throughout the text, data are expressed as mean±SD.
Results
Complexes of hgDNA and LL37 (LL37-DNA) enhance effector functions of NK cells
To investigate the effects of LL37-DNA on human NK cells, we cultured healthy donor PBMCs with the medium, hgDNA alone (10 μg/mL), LL37 alone (50 μg/mL), or with an equivalent concentration of LL37-DNA complexes, whereafter we measured NK cell degranulation upon a 4-h coincubation with K562 target cells. Treatment with LL37-DNA caused more than a 4-fold increase in degranulation of NK cells compared with medium treatment (Fig. 1A). hgDNA had no effect, while LL37 alone caused a modest increase in degranulation, which was not statistically significant. Thus, the strong enhancement of NK cell degranulation induced by LL37-DNA is mediated by the entire complex and not by one of its components.

The LL37-DNA complexes enhance degranulation and natural cytotoxicity of NK cells.
We also measured natural cytotoxicity of PBMCs toward K562 target cells after the same treatments. The changes of cytotoxicity (at an E:T ratio of 50:1) closely paralleled changes of degranulation (Fig. 1B). At lower E:T ratios (down to 6:1), LL37-DNA-treated PBMCs were still superior to medium-treated PBMCs at killing K562 target cells (Fig. 1C). LL37-DNA-treated PBMCs contained a 6-fold greater amount of lytic units 20 (LU20) per 105 cells compared with medium-treated PBMCs (2.4±1.43 versus 0.36±0.26, P<0.05 in paired t-test).
To make sure that the increased killing of K562 targets by LL37-DNA-treated PBMCs was mediated by NK cells, we depleted NK cells from these PBMCs using CD56 microbeads. The percentage of NK cells among CD56-depleted PBMCs was 1.1%±0.9% versus 17.7%±0.3% among mock-depleted PBMCs. As expected, this manipulation virtually abrogated the killing (Fig. 1D). In addition, LL37-DNA did not induce any degranulation responses in the CD3+CD56− or CD3+CD56+ subpopulations of PBMCs (not shown).
Treatment with LL37-DNA did not influence percentages of NK cells among PBMCs, nor the intracellular expression of perforin and granzymes by NK cells (Fig. 1E–H), excluding the role for these factors in the increased natural cytotoxicity of PBMCs.
To analyze the effect of LL37-DNA on the production of IFN-γ by NK cells, PBMCs were treated overnight with LL37-DNA or medium alone, washed, and cultured for 4 h with or without K562 targets in the presence of monensin. As shown in Table 2, treatment with LL37-DNA yielded a nearly 4-fold increase in the percentages of IFN-γ+ NK cells upon subsequent culture with K562 cells (12.7%±4.5% compared with 3.6%±2.0% in the medium-treated NK cells, n=3). Most of the IFN-γ-producing NK cells coexternalized CD107a (Fig. 2, Table 2). Thus, treatment with LL37-DNA enhances not only degranulation but also IFN-γ production by NK cells upon subsequent encounter with target cells.

The influence of LL37-DNA on interferon (IFN)-γ production by NK cells. PBMCs were treated overnight with medium or LL37-DNA, then incubated for 4 h without or with K562 cells, followed by a simultaneous analysis of NK cell degranulation and intracellular IFN-γ (see the Materials and Methods section). Shown are gated CD3−CD56+ NK cells (1 representative experiment of 3).
All percentages were calculated in relation to gated CD3−CD56+ NK cells. See the Materials and Methods section for experimental details. n=3 values are expressed in mean±SD.
IFN, interferon.
The effect of LL37-DNA on NK cells is indirect and mediated by type I IFNs
Next, we asked whether LL37-DNA activate NK cells directly or indirectly. NK cells were isolated from PBMC suspensions by negative immunomagnetic sorting (with a purity of >95%) and treated overnight with LL37-DNA or medium alone, whereafter they were tested for degranulation upon coincubation with K562 targets. LL37-DNA strongly increased degranulation of NK cells within unfractionated PBMCs, but did not influence degranulation of purified NK cells from the same donors (Fig. 3A). We concluded that LL37-DNA complexes act upon (a)other subpopulation(s) of cells within PBMCs, which in turn release NK-activating factors, probably cytokines. Interestingly, purified NK cells, irrespective of LL37-DNA treatment, degranulated somewhat stronger than NK cells within untreated PBMCs, which might be due to improved NK-target cell contacts or to removal of a degranulation-suppressing subpopulation of PBMCs.
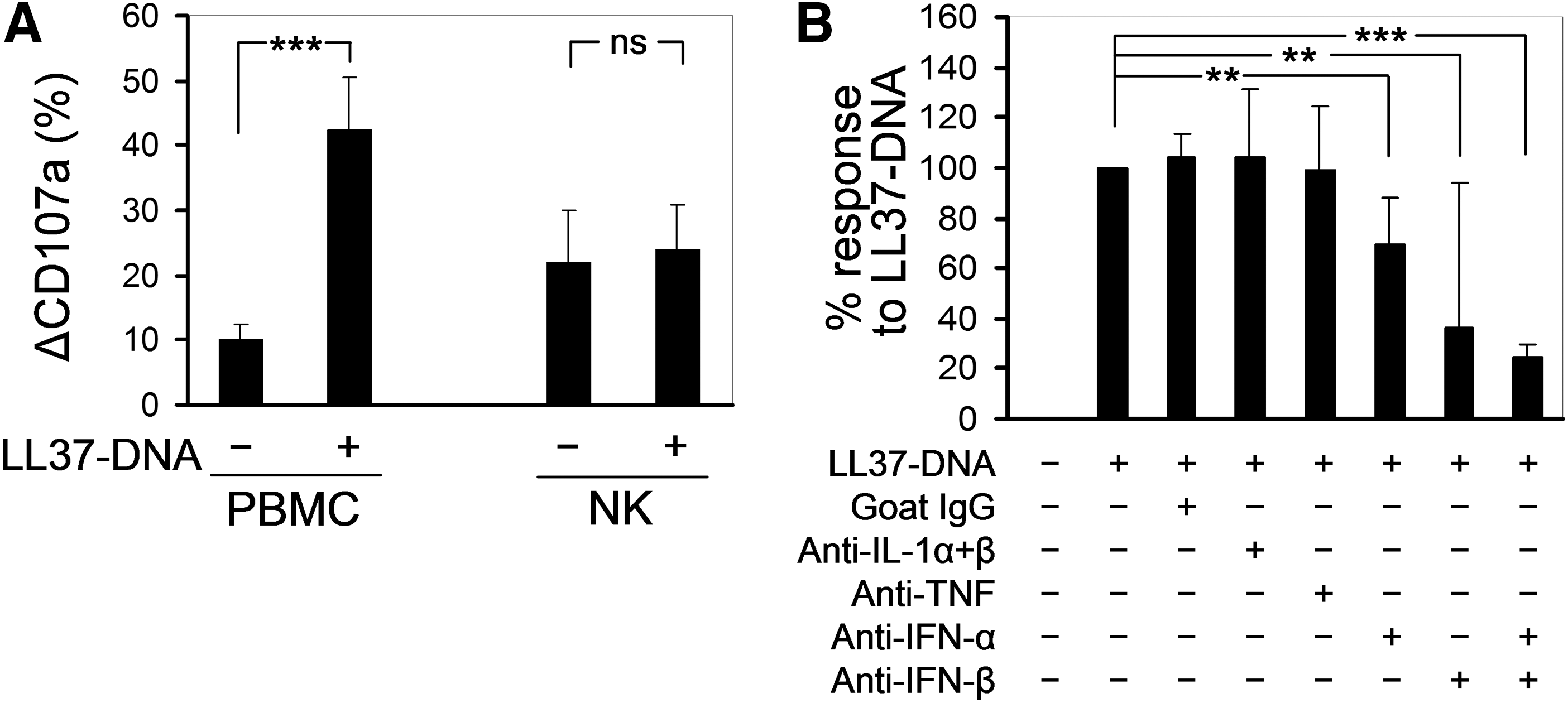
LL37-DNA activate NK cells through an indirect mechanism mediated by type I IFNs.
Earlier studies have shown that LL37-DNA complexes target at least 2 types of cells in human blood: ppDCs and monocytes; both respond to LL37-DNA by producing type I IFNs, although with different kinetics (Chamilos and others 2012). We found that PBMCs stimulated with LL37-DNA secreted IFN-α and, to a lesser extent, TNF, but not IL-1β and IL-6 (Table 3). None of the 4 cytokines was induced by LL37-DNA in purified NK cells (data not shown).
n=7 for IL-1β, IL-6, TNF, and IFN-α; n=6 for IFNB. Values are expressed in mean±SD.
P<0.05 compared with medium-treated PBMCs.
P<0.01 compared with medium-treated PBMCs.
IL, Interleukin; PBMC, peripheral blood mononuclear cell; RE, relative expression; TNF, tumor necrosis factor.
We could not reliably measure IFN-β in cell culture supernatants because the commercial ELISA kit we used showed a significant cross-reactivity with LL37 present in the samples. We therefore measured IFNB mRNA expression by RT-PCR and found that the IFNB gene was significantly induced in PBMCs by LL37-DNA, whereas LL37 and DNA separately had no significant effect (Table 3).
It is well known that type I IFNs are potent activators of NK cells (Theofilopoulos and others 2005). In our hands, recombinant human IFN-α2b displayed an activity that was qualitatively similar to that of LL37-DNA, namely the enhancement of NK cell degranulation with no effect on percentages of NK cells or expression of perforin (data not shown). We then used neutralizing antibodies against several cytokines, including type I IFNs, to block the enhancing effect of LL37-DNA on NK cell degranulation within unfractionated PBMCs. Antibodies against IL-1α, IL-1β, or TNF had no effect, whereas antibodies against IFN-α and IFN-β significantly downregulated the LL37-DNA-induced enhancement of NK cell degranulation (Fig. 3B). Interestingly, relative contributions of IFN-α and especially IFN-β seemed to vary from donor to donor, which is depicted by large standard deviations (Fig. 3B); however, simultaneous neutralization of both cytokines almost completely canceled the enhancing effect of LL37-DNA on NK cell degranulation in all donors studied (Fig. 3B).
The effect of LL37-DNA on NK cells is mediated by monocytes and ppDCs
To establish cellular sources of the type I IFNs that activate NK cells in our system, we depleted ppDCs or monocytes from PBMC suspensions using mAbs against BDCA-2 or CD14, respectively, then treated the depleted PBMCs with LL37-DNA and measured degranulation of NK cells in response to K562 targets. The BDCA-2 mAb depleted 92%–100% ppDCs; however, this resulted in only a minor reduction of the enhancing effect of LL37-DNA on degranulation compared with mock-depleted cultures (Fig. 4A). The CD14 mAb depleted all CD14high monocytes and a fraction of the CD14dim monocytes. In the CD14-depleted PBMCs, the enhancing effect of LL37-DNA on NK cell degranulation was significantly reduced (Fig. 4A) and in some donors was completely absent.
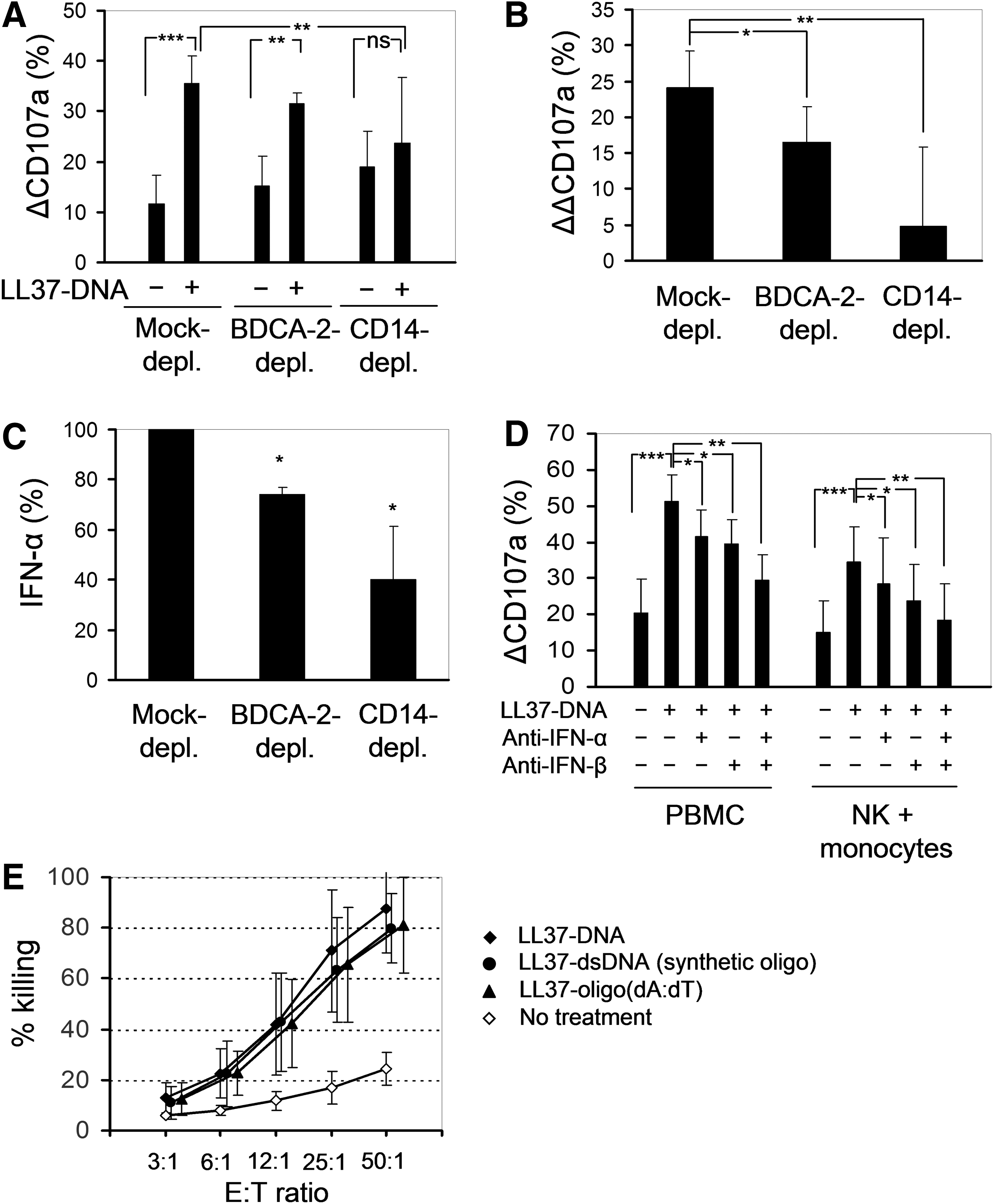
Activation of NK cells in LL37-DNA-stimulated PBMC cultures is mediated mainly by monocytes.
We observed that depletion of ppDCs and especially of monocytes resulted in an increase of NK cell degranulation even in the absence of LL37-DNA treatment (Fig. 4A, see also Fig. 3A), which partly obscured the effect of LL37-DNA. It is known that freshly isolated monocytes suppress NK activity (Jondal and others 1981; Yang and Zucker-Franklin 1984; Pedersen and others 1986), so monocyte depletion apparently reverses this suppressive effect. Therefore, for each mock-depleted BDCA-2-depleted and CD14-depleted sample, we calculated a ΔΔCD107a, which is ΔCD107a after LL37-DNA treatment minus ΔCD107a after medium treatment. This parameter, which enables to distill the effect of LL37-DNA, was only modestly reduced in BDCA-2-depleted PBMCs and strongly reduced in the CD14-depleted PBMCs (Fig. 4B). We speculate that the remaining activity of LL37-DNA in the CD14-depleted fraction could be mediated by the remaining CD14dim monocytes, myeloid dendritic cells, or ppDCs (Chamilos and others 2012). Depletion of B cells using a mAb against CD19 did not impair the ability of LL37-DNA to enhance NK cell degranulation (data not shown).
To confirm the role of monocytes as chief activators of NK cells in our system, we performed 3 series of experiments. First, we measured IFN-α secretion by mock-depleted BDCA-2-depleted and CD14-depleted PBMCs upon an overnight treatment with LL37-DNA. In the CD14-depleted fraction, secretion of IFN-α was reduced by about 60% compared with only 26% in the BDCA-2-depleted fraction (Fig. 4C).
Second, we performed overnight cocultures of purified NK cells and monocytes at a ratio of 1:1 in the presence of LL37-DNA or the medium, whereafter we measured degranulation of NK cells in response to K562 (Fig. 4D). The fraction of positively selected monocytes used in these experiments consisted mainly of CD14high cells (89.0%±0.6%), with some admixture of CD14low monocytes (8.0%±0.7%). In these monocyte-rich cocultures, degranulation responses of NK cells were generally lower than in PBMC suspensions (Fig. 4D), which was the reverse of what had been observed in monocyte depletion experiments (Fig. 4A). Nonetheless, in the NK-monocyte cocultures, LL37-DNA caused a 2.5-fold increase of NK cell degranulation (comparable with the effect in unfractionated PBMCs), which was partially blocked by Abs against IFN-β and, to a lesser extent, by Abs against IFN-α, whereas the 2 Abs together had an additive effect, neutralizing more than 80% of the activating effect of LL37-DNA toward NK cells (Fig. 4D).
Third, it has been shown that monocytes respond to oligo-(dA:dT) complexed with LL37, whereas ppDCs do not (Chamilos and others 2012); furthermore, ppDCs poorly respond to methylated DNA complexed with LL37 (Chamilos and others 2012). Therefore, we treated PBMCs by LL37 complexed with hgDNA used in all above experiments or with oligo-(dA:dT) or with a synthetic fully methylated dsODN. All 3 types of complexes similarly enhanced natural cytotoxicity of PBMCs (Fig. 4E). Thus, we conclude that the enhancing effect of LL37-DNA on NK cell degranulation is mediated, for the most part, by monocytes producing type I IFNs.
Discussion
In this study, we confirm earlier reports, which demonstrated that LL37 converts nonstimulatory DNA (of either genomic or synthetic origin) into a potent activator of innate immune responses (Lande and others 2007; Chamilos and others 2012). We show that complexes of LL37 with hgDNA or synthetic dsODN enhance degranulation and natural cytotoxicity of human NK cells in vitro (Fig. 1). The enhanced production of IFN-γ by LL37-DNA-treated NK cells (Fig. 2) may contribute to their increased cytotoxicity (Berthou and others 2000). Finally, we show that the effect of LL37-DNA on NK cells is indirect and mediated by type I IFNs, which are produced by LL37-DNA-activated monocytes and, to a lesser extent, by other IFN-producing cells. Our data argue against a direct activation of NK cells by LL37-DNA.
Contrary to expectation, it was monocytes and not ppDCs that played the chief role in the production of type I IFNs and activation of NK cells upon LL37-DNA treatment. The role of monocytes as the predominant source of type I IFNs in our system can be explained by comparing the amounts of IFNs produced by monocytes and ppDCs (per cell) and their abundance in blood. While each individual monocyte stimulated with LL37-DNA produces around 10 times less IFNs than a ppDC (Chamilos and others 2012), monocytes are 20–30 times more numerous in blood than ppDCs. For example, in the experimental series shown in Fig. 4C, CD14bright monocytes constituted 14.71%±2.76% of PBMCs and ppDCs only 0.55%±0.22% (a 26.7 times difference). Therefore, upon LL37-DNA treatment, the population of monocytes makes more type I IFNs than the population of ppDCs, even though each individual monocyte produces much less type I IFN than each ppDC. The role of monocytes is further supported by the fact that NK cytotoxicity was augmented by complexes of LL37 with oligo-(dA:dT), to which ppDCs do not respond, as well as by complexes of LL37 with a fully methylated dsODN, to which ppDCs respond poorly (Chamilos and others 2012). However, we do not exclude that in tissue environments where ppDCs are more abundant, they will have a stronger relative impact on NK cell activation.
Interestingly, monocytes that had not been treated with LL37-DNA appeared to suppress, rather than to activate NK cells, because (1) depletion of monocytes resulted in an increased degranulation of NK cells upon encounter of target cells (Fig. 4A) and (2) adding excess of monocytes to NK cells resulted in a reduced degranulation (Fig. 4D). These data fit very well with works from 1980s, which documented the suppressive activity of freshly isolated monocytes toward NK cells and linked this suppressive effect to monocyte-derived prostaglandins (Jondal and others 1981; Yang and Zucker-Franklin 1984; Pedersen and others 1986). Thus, it can be concluded that treatment with LL37-DNA transforms monocytes from an NK-suppressing into an NK-activating population.
We also found that even LL37 alone enhanced NK cell degranulation and cytotoxicity, although to a lesser extent than LL37-DNA (Fig. 1A, B). These data are reminiscent of those by Lande and others (2007), who reported that LL37 alone can induce type I IFN production in the cultures of ppDCs. We agree with the suggestion that LL37 may bind to endogenous nucleic acids released from dying cells (Lande and others 2007), so the LL37-DNA complexes may be formed directly in the culture wells, although DNA-independent stimulatory effects of LL37 cannot be excluded (Vandamme and others 2012).
Our findings are relevant both to the regulation of immune processes and to the therapeutic manipulation of the immune system. It is known that small constitutively produced amounts of IFN-β are key factors required for the proper functioning of diverse immune cells (Gough and others 2012), including NK cells (Lee and others 2000). However, factors regulating the constitutive IFN-β production itself have not been identified. It is tempting to speculate that one such factor could be the LL37-DNA complexes that emerge during the steady-state cell turnover. Targets of LL37-DNA in tissues could be macrophages, which are the progeny of monocytes, or keratinocytes, which can respond to LL37-DNA under certain conditions (Chi and others 2014). However, the existence of such a steady-state mechanism involving LL37 as a regulator of NK cell homeostasis awaits direct demonstration. So far, only one study has examined NK cell functions in CAMP–/–mice, which lack LL37 (Buchau and others 2010). NK cells from these mice display reduced cytotoxicity (which is not restored by treatment with IL-2) as well as altered expression of perforin and granzyme B. However, since LL37 is stored in NK cells (Buchau and others 2010) and can be directly cytotoxic at high concentrations (Zhang and others 2008), these results do not allow to distinguish whether LL37 acts as a cytotoxic effector molecule utilized by NK cells or it is a regulatory factor that alone or in combination with nucleic acids regulates NK cell homeostasis. In inflammatory conditions, the production of LL37-DNA complexes has been clearly documented (Lande and others 2007, 2011), so they are much more likely to affect NK cells both in the inflamed tissue and systemically.
Second, complexes of LL37 with synthetic DNA might be viewed as potential pharmacological agents to enhance NK cell cytotoxicity in relevant clinical situations, such as the treatment of cancer. At the first glance, the mechanism of action of LL37-DNA would be similar to that of CpG-ODNs, which have been extensively studied as antitumor agents in animals and in clinical trials (Krieg 2012). However, the single-stranded CpG-ODN molecules are sensed by ppDCs through TLR9, and the choice of an optimal TLR9-activating ODN sequence has always been an issue (Krieg 2012). In the case of LL37-DNA complexes, this issue would be solved because even oligo-(dA:dT) complexed with LL37 can boost NK activity (Fig. 4E). This is because LL37-DNA complexes efficiently target monocytes, which recognize DNA by means of cytosolic DNA sensors (Chamilos and others 2012). As shown earlier, LL37 can also enhance the antitumor activity of traditional CpG-ODNs (Chuang and others 2009); this can be explained both by the ability of LL37 to improve CpG-ODN recognition by ppDCs (Hurtado and Peh 2010) and by targeting CpG-ODNs to monocytes, which altogether results in enhanced type I IFN production and augmented activation of NK cells. Importantly, since monocytes/macrophages are more numerous in the tumor tissue than ppDCs, they may have a stronger impact on the local milieu, resulting in a stronger activation of NK cells within the tumor and thus in a better tumor rejection.
In all, we have dissected a mechanism by which LL37-DNA complexes activate human NK cells. Further studies are warranted to clarify mechanisms of action of LL37-DNA complexes as well as their utility as therapeutic agents.
Footnotes
Acknowledgment
The authors thank Dr. A. Kalashnikov for his assistance in preparation of dsODNs.
Author Disclosure Statement
No competing financial interests exist.