
Abstract
Interleukin (IL)-37 is a newly discovered member of the cytokine IL-1 family. Recent evidence suggests that IL-37, an anti-inflammatory factor, may have a role in atherosclerosis. In this study we used apoE-deficient diabetic mice, an established animal model, to examine the effects of IL-37 on the progression of vascular calcification and atherosclerosis. Compared with the control groups, IL-37-treated (with injection of recombinant protein for 16 weeks) animals had significantly less calcification areas detected by both von Kossa and Alizarin Red staining, and much smaller plaque size of the atherosclerotic lesions and lower plaque vulnerability scores detected by hematoxylin–eosin staining in the aorta root. Our data also showed that IL-37 treatment caused elevated concentrations of osteoprotegerin (OPG) in serum. We detected that the group that received additional anti-OPG antibody reduced the effect of IL-37 treatment. The group that received both IL-37 and anti-OPG had significant larger percentage area of calcified lesion and atherosclerotic plaque size than the IL-37-treated group. Significant changes in disease-relevant cytokines (eg, ALP, BMP-2, TNF-α, IL-18, and IL-10) were also elicited. This is the first report that IL-37 could attenuate not only atherosclerosis, but also vascular calcification. This study may offer a therapeutic potential for the prevention and treatment of calcification and atherosclerotic disease.
Introduction
A
For several decades, vascular calcification has been a known consequence of aging. Recent studies have confirmed that vascular calcification is an actively regulated process and shares many features with bone development and metabolism (Johnson and others 2006); the process is closely related with chronic inflammation in mice and human (Abdelbaky and others 2013). Macrophages, lymphocytes, and dendritic cells infiltrate plaque and release cytokines that regulate calcification (Li and others 2010). Perivascular adipose inflammation (Takaoka and others 2009) and systemic inflammation (Shroff and others 2008) may also contribute to both neointimal and medial calcification. Moreover, several studies have confirmed that proinflammatory factors, such as tumor necrosis factor (TNF) (Al-Aly and others 2007) and leptin (Zeadin and others 2009), increase vascular calcification by promoting osteogenic differentiation, while anti-inflammatory cytokine-like omentin-1 (Xie and others 2011) and adiponectin (Luo and others 2009) ameliorates vascular calcification.
The novel anti-inflammatory cytokine interleukin (IL)-37 is a newly discovered member of the IL-1 family (Nold and others 2010; Boraschi and others 2011). Recent evidence has shown that IL-37 plays a protective role in inflammatory and autoimmune diseases in animal models through the inhibition of proinflammatory cytokine generation and the activation of macrophage and dendritic cells (Nold and others 2010; McNamee and others 2011; Sakai and others 2012). IL-37 protein has been detected in the foam-like cells of human atherosclerotic coronary and carotid plaque (Boraschi and others 2011), suggesting that IL-37 is involved in the development of atherosclerotic disease. More recently, IL-37 has been shown to play also a protective role against mouse myocardial ischemia/reperfusion (I/R) injury (Wu and others 2014). As calcification typically follows the development of atherosclerotic lesions and inflammation is involved in the progression of arterial calcification, we hypothesized that IL-37 may regulate the process of atherosclerosis and may be involved in the progression of arterial calcification.
Thus, we used an established animal model to examine the effect of IL-37 on vascular calcification and atherosclerosis. During preliminary experiment, we noticed that the serum of IL-37-treated mice had unusually high concentrations of osteoprotegerin (OPG), which has been shown to be able to inhibit vascular calcification in mice. OPG is a cytokine of the TNF receptor superfamily and is classified as an osteoclastogenesis inhibitory factor. Many animal studies have suggested that OPG has a beneficial effect on vascular calcification and atherosclerosis (Bennett and others 2006). Therefore, we considered that elevated levels of IL-37 may inhibit vascular calcification and atherosclerosis in vivo and included anti-OPG antibody in one of our control experiments to investigate whether the effect of IL-37 on vascular calcification and atherosclerosis is through OPG.
Materials and Methods
Animals
Twenty-eight male C57BL/6 Apolipoprotein E-deficient (ApoE−/−) mice (8 weeks old) were obtained from Beijing University (Beijing, China). All of the mice were housed inside a specific pathogen-free environment with a12-h light–12-h dark cycle. The animal care and experimental protocols were in compliance with the Animal Management Rule of the Ministry of Health, People's Republic of China.
Experimental design
All of the mice were given intraperitoneal (i.p.) injections of streptozotocin (STZ) for 5 days, 40 mg/kg in citrate buffer (0.05 M, pH4.5). Mice with blood glucose levels of >300 mg/dL at 2 weeks after the initial STZ administration were considered diabetic and were included in the current study. All the mice were fed on regular chow until 2 weeks after the start of the STZ administration. The mice were then switched to a semisynthetic high-fat diet (21% fat, 0.15% cholesterol, and other contents, the same with standard chow). Nɛ-carboxymethyl-lysine (CML), a major immunogen of advanced glycation end products (AGEs) that affects the progression of atherosclerotic calcification in diabetes, was administered intraperitoneally in normal saline at the daily dose of 60 μg for 16 weeks (Menini and others 2007; Wang and others 2012).
After a change in diet, all of the mice were randomly distributed into 4 groups, with 7 animals per group. The experimental group received weekly i.p. injections of 2 μg (Wu and others 2014) recombinant human IL-37 (Adipogen, Liestal, Switzerland) suspended in 200 μL of normal saline. The control group received an equivalent volume of saline (200 μL i.p./mouse) only. The third group received both IL-37 (2 μg) and anti-OPG antibody (200 μg; R&D Systems) weekly i.p. The fourth group was similarly treated as the third group, but with an isotype mAb administered intraperitoneally on a weekly basis. Sixteen weeks after the diet-switch (Wang and others 2012), blood was collected and the heart, including the thoracic aorta, was removed after general anesthesia for histological evaluation.
Serum analysis
After fasting for 4 h, the mice were placed under general anesthesia (a tactile stimulus failed to induce a response, and the forelimb or hindlimb pedal withdrawal reflex response was absent or delayed) with pentobarbital sodium (50 mg/kg i.p.); the mice remained alive. Blood was collected from the retro-orbital venous plexus using a capillary tube. Serum concentrations of plasma lipid levels (total cholesterol, triglycerides, low and high density lipoprotein) were measured using colorimetric diagnostic kits (Crystal Chem). Blood glucose levels were measured using a glucometer (Bayer). Plasma OPG level was determined by ELISA using commercially available kits (R&D Systems). TNF-α concentration was determined by radioimmunoassay (Linco).
Tissue sample preparation and histology
After the animals were euthanized, the heart and thoracic aorta were removed. The aortic root was separated from the aorta and embedded in optimal cutting temperature gel before storage at −80°C. Serial cross-sections beginning at the aortic valves were collected and measurements of lesion sizes were performed at the same point within the aortic root for each animal. Six serial sections per mouse were analyzed.
Quantification of vascular calcification
Calcification of the aortic root was detected by von Kossa staining and Alizarin Red staining. The cryosections were placed briefly in 5% silver nitrate solution for 30 min in the dark. The sections were then treated with film developer for 5 min and finally fixed in 5% sodium thiosulfate. Calcification was visualized as distinct black deposits of calcium using an epifluorescent microscope coupled with an IBM computer. All images were captured using a 3CCD video camera module and analyzed using the Northern Eclipse imaging software system (Empix Imaging). For Alizarin Red staining, sections were dissected and stained with 0.005% Alizarin Red S in 0.5% KOH for 24 h. For histological analysis, aortas were fixed in 4% formaldehyde overnight. For paraffin sections, fixed aortas were dehydrated and embedded in paraffin and cut into sections of 5 μm thickness. Measurements of the calcified areas were expressed as a percentage of a lesion's total size.
Measurement of atherosclerosis
Hematoxylin–Eosin (HE) staining was used to measure the morphological changes in atherosclerotic lesions of the aorta as measured by histological section. Briefly, the aorta was cut into transversal sections of 5 μm and fixed in 10% formalin for 24 h. The tissue slices were processed and embedded in paraffin. Measurement of the atherosclerotic area was classified as the lesion area (Phan and others 2005).
Histopathologically stained slides were scanned by microscope and analyzed by a computer-assisted morphometric analysis system (Image-Pro Plus 5.0, Media Cybernetics). The vulnerability index was calculated using the following relationship: (macrophage staining% +lipid staining%)/(vascular smooth muscle cells% +collagen fiber%) (Shiomi and others 2001).
Immunohistochemistry analysis
Staining of OPG, alkaline phosphatase (ALP), and bone morphogenetic proteins 2 (BMP-2) of the aortic root were used to identify osteoblast-like cells present in the atherosclerotic lesions of the 4 groups of mice. IL-10, TNF-α, and IL-18 staining of the aortic root were also used to identify inflammatory mediators present in the atherosclerotic lesions of the 4 groups of mice.
Anti-Osteoprotegerin antibody (9986; Abcam), anti-ALP antibody [4H1] (54778; Abcam), Anti-BMP2 antibody (14933; Abcam), Anti-IL-10 antibody (JES5-2A5, 189392; Abcam), Anti-TNF alpha antibody (52B83, 1793; Abcam), and Anti-IL-18 antibody (71495; Abcam) were used as the primary antibodies. The slides were incubated with the secondary antibody (biotin-labeled rabbit anti-mouse IgG antibody; Nichirei Co) at room temperature for 30 min, followed by peroxidase-conjugated streptavidin (Nichirei) at 37°C for 30 min. After washing with PBS, peroxidase activity was detected by reaction with 3,3-diaminobenzidine tetrahydrochloride (DAB tablet, 30 mg/mL, containing 0.03% H2O2; Merck) as the chromogen. Finally, sections were counterstained by 1% methylene green, dehydrated, and mounted. Staining without the primary antibody was used as a negative control.
Statistics
The data were presented as the means±SD. Comparisons were made using a 1-way analysis of variance (post hoc). The GraphPad Prism (Version 6.0) software was used for statistical analyses. P values less than 0.05 were considered significant.
Results
Body weight and serum biochemical analyses
Among the 4 experimental mouse groups, including those treated with IL-37, IL-37+anti-OPG antibody, IL-37+an isotype mAb, and the untreated control, there were no major difference in body weight, serum glucose, or plasma lipid levels, including total cholesterol, triglycerides, and low- and high-density lipoprotein (Supplementary Table S1; Supplementary Data are available online at
TNF-α is a proinflammatory factor and it was reduced in IL-37-treated mice compared with the control group (1.81±0.22 vs. 2.47±0.33 ng/mL; P=0.001; Supplementary Table S1). However, there were no significant statistical difference (P=0.296) between groups treated with IL-37, IL-37+anti-OPG, and IL-37+isotype anti-OPG.
Interestingly, plasma OPG level was significantly elevated in IL-37-treated mice (3.02±0.60 ng/mL) compared with the control group (1.73±0.52 ng/mL). This result was similar to the preliminary experiment that led us to the prediction that OPG may play an important role in the effect of IL-37 on calcification and on atherosclerosis. Understandably, the group treated with IL-37 and anti-OPG antibody showed lower plasma OPG levels compared with IL-37-treated mice (P<0.001), but the group treated with IL-37 and isotype antibody showed a similar level of OPG as in the IL-37-treated mice (P=0.436, Supplementary Table S1).
Effect of IL-37 on the vascular calcifications
To determine the effect of IL-37 on vascular calcification, we compared the percentage area of calcium deposit in the aortic root. The calcified area was stained with 2 staining reagents separately to ensure the reliability of the results: von Kossa (Fig. 1) and Alizarin Red staining (Supplementary Fig. S1). A large black area together with small punctate spots (von Kossa staining: 1.56%±0.35%, Fig. 1A; Alizarin Red S: 1.42%±0.42%; Supplementary Table S2) were detected in the mice of the untreated control group. However, in the aortic root of the IL-37-treated animals, only very small amounts of scattered punctate areas of calcification were observed (von Kossa staining: 0.29%±0.30%; Fig. 1B; and Alizarin Red S: 0.21%±0.22% Supplementary Fig. S1 and Table S2). The values are of statistical significance with P<0.001 and the data clearly showed that IL-37-treated animals have statistically significantly less calcification in areas within the aortic root compared with the untreated mice (Fig. 1E).
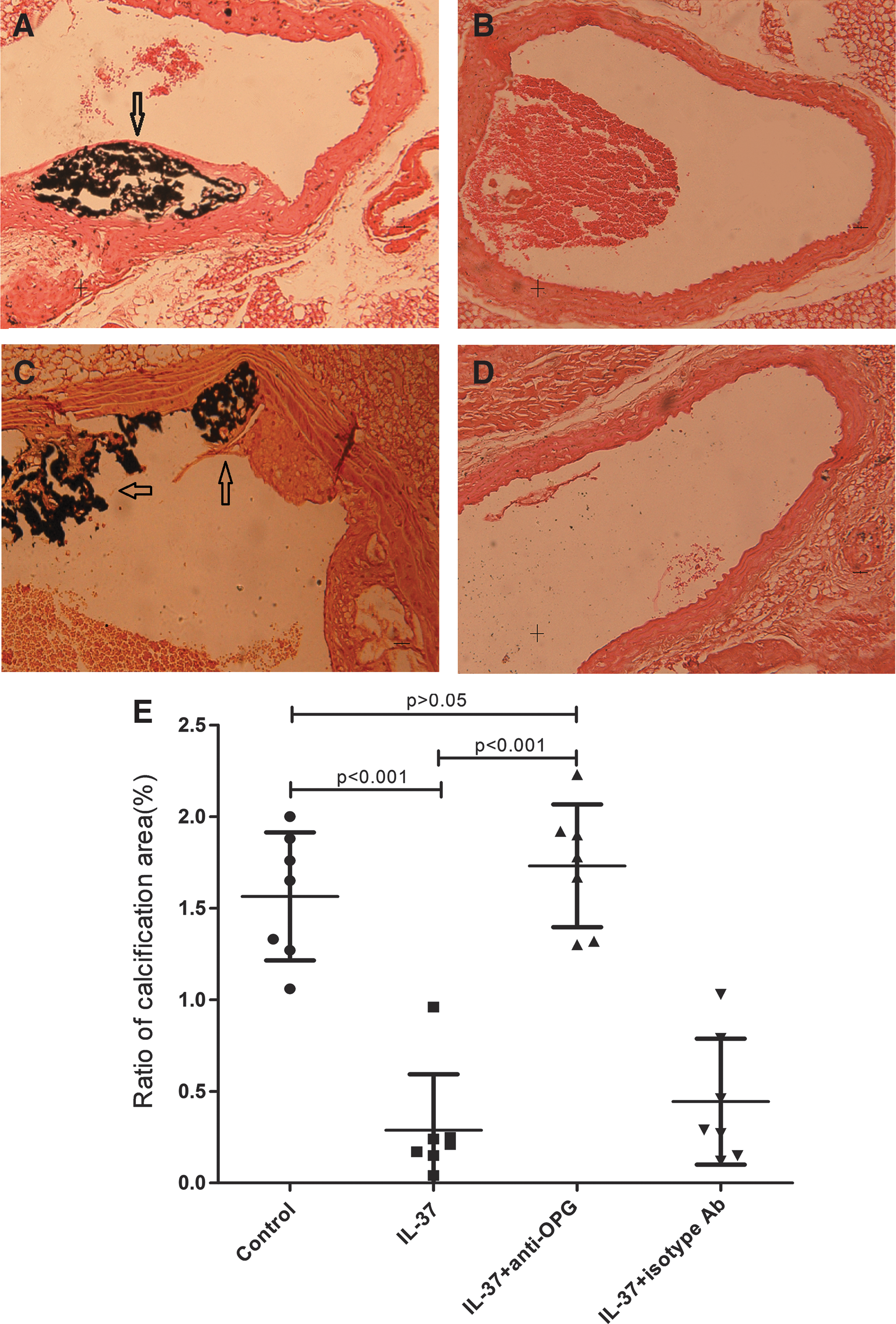
Effect of lL-37 on the calcification of atherosclerotic lesions by von Kossa staining. Vascular calcification in groups of untreated
We also observed an elevated level of OPG in the IL-37-treated ApoE−/− mice. To minimize the potential OPG effect, we used a specific anti-OPG antibody to block OPG signaling pathway in one of the experimental groups while using an isotype antibody as a control. As shown by von Kossa staining, treatment with IL-37 and anti-OPG had much larger percentage area of calcified lesion than the IL-37-treated group (1.73%±0.34% vs. 0.29%±0.30%, P<0.001; Fig. 1C). The anti-OPG control group (treated with both IL-37 and isotype antibody) did not show the same effect as the calcified area was remained roughly the same as the IL-37-treated group (0.44%±0.34%; Fig. 1D, E). Similar results were obtained with Alizarin Red staining (Supplementary Table S2). These data clearly indicated that the decrease of the calcification after treatment with IL-37 is closely related with the increase of OPG.
Effects of IL-37 on atherosclerosis
To determine whether IL-37 has an effect on atherosclerosis, we assessed the morphological changes of atherosclerotic lesions by comparing the plaque size of the lesions in the aorta root. The comparison was made by measuring the area of plague based on HE staining, before and after IL-37 treatment. As shown in Fig. 2, atherosclerotic area significantly decreased in the IL-37-treated group (0.20±0.17 mm2; Fig. 2B and Supplementary Table S2), compared with the control mice (2.90±0.24 mm2; Fig. 2A). To explore the relationship between plaque vulnerability and IL-37 treatment, we calculated the vulnerability index, a parameter for determination of the destabilization of vulnerable plaques, which ranged from 0.5 to 1.5 in our study. Again, we found a similar trend to the effect of IL-37 on the calcification of atherosclerotic lesion, although the effect was not as profound as plague area measurement. The plaque was stable in the IL-37 treatment group (index=0.76±0.26) whereas in the control group the plaque was vulnerable (index=1.24±0.24) (P=0.003).
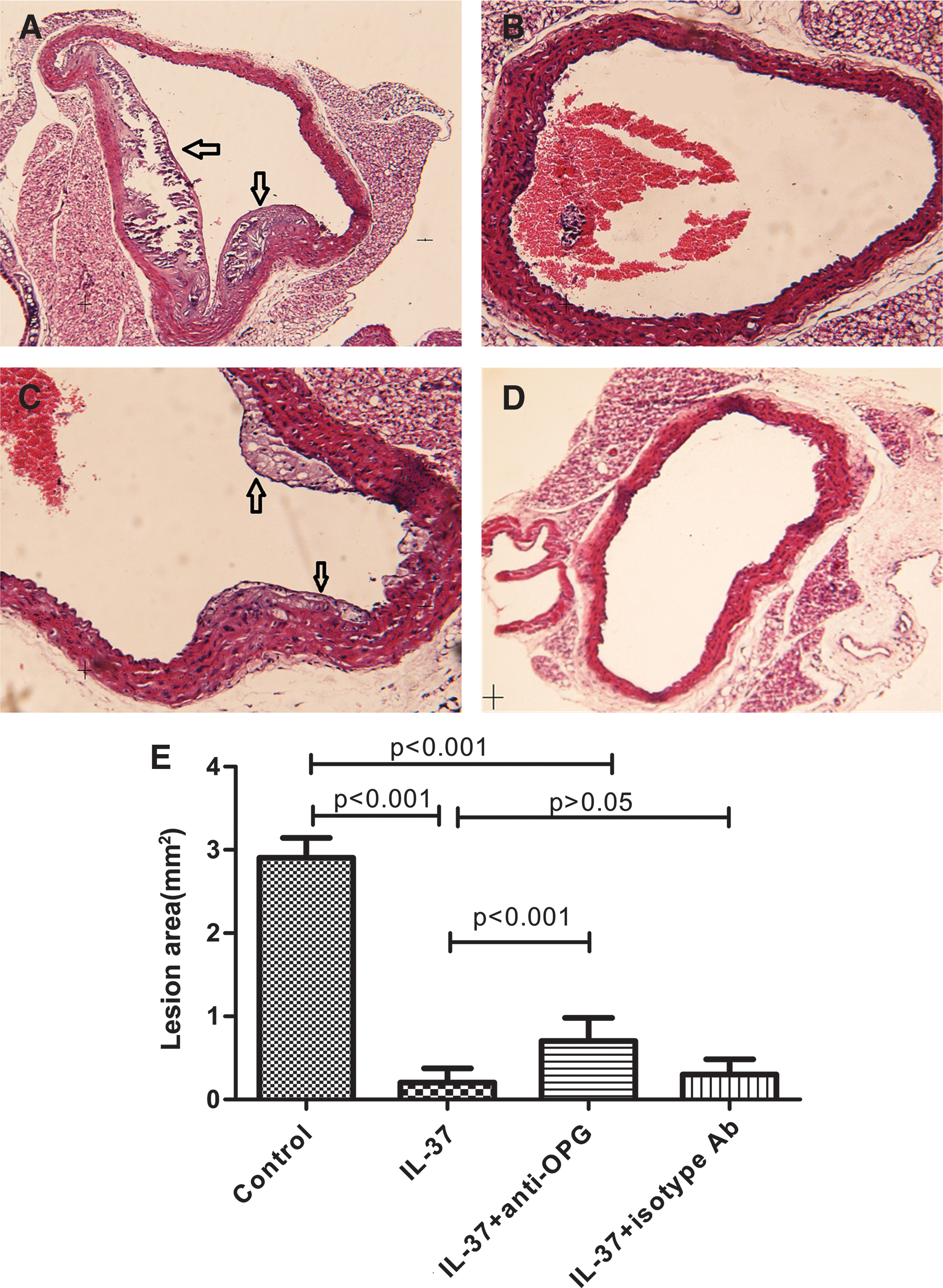
Effect of lL-37 on atherosclerosis. The lesion in groups of untreated
Furthermore, both treatments with IL-37+anti-OPG and IL-37+isotype antibody were found to have a smaller atherosclerotic area and a lower vulnerability index (P<0.001; Fig. 2) compared with the control. However, compared with the IL-37-treated group, treatment with IL-37 and anti-OPG increased both the plaque size of the atherosclerotic lesions (0.71±0.28, P<0.001; Fig. 2E) and the vulnerability index (1.15±0.23, P<0.001; Supplementary Table S2), whereas treatment with IL-37 and isotype antibody could not. With these results, we are able to conclude that the treatment with anti-OPG could only partially prevent the protective effects of IL-37 on atherosclerosis and the inhibitory effect of IL-37 on calcification seems more profound than on arteriosclerotic lesions.
Immunohistochemical staining of OPG
As shown in Fig. 3B, our study indicated that IL-37-treated animals had larger OPG-positive areas (brown staining, 1.65%±0.48%, P<0.001), expressed by endothelial cells and vascular smooth muscle cells (VSMCs), compared with the control animals (0.66%±0.14%; Fig. 3A). We also included an anti-OPG antibody to minimize the potential role of OPG. As shown in Fig. 3C, treatment with both IL-37 and anti-OPG indeed suppressed the production of OPG, as the OPG-positive staining was greatly reduced compared with IL-37-treated group (0.44%±0.23% vs. 1.65%±0.48%, P<0.001; Fig. 3C, B and Supplementary Table S3). To confirm the role of OPG, an isotype antibody was included as control. As shown in Fig. 3D, the OPG-positive area (1.58%±0.31%) was increased compared with the group treated with IL-37 and anti-OPG, but was similar to the IL-37 only treated group indicating that the OPG cannot be suppressed by this isotype antibody.
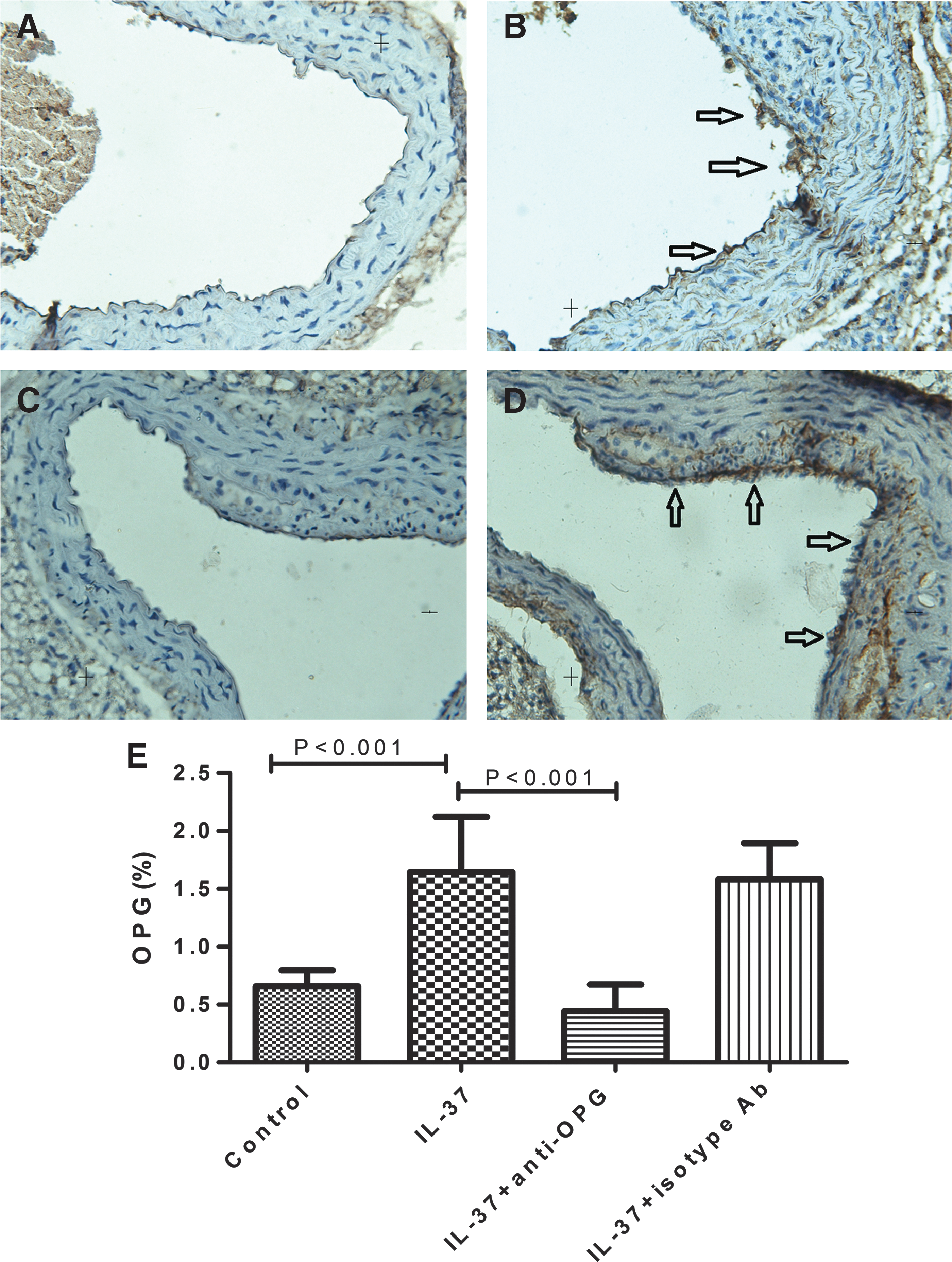
Immunohistochemistry of OPG in groups of untreated
Effect of IL-37 on the osteoblast differentiation markers ALP and BMP-2
ALP and BMP-2 are regulatory factors of vascular calcification (Sage and others 2010). The IL-37 effect on their expression was examined by immunohistochemical method. We found that the IL-37-treated animals had less areas that stained positive for ALP and BMP-2 compared with control animals (0.85%±0.37% vs. 2.42%±1.05%, 0.87%±0.57% vs. 1.64%±0.43%, respectively, P<0.05; Fig. 4A, B and Supplementary Table S3). The result indicated that IL-37 can reduce the expression of ALP and BMP-2, consistent with the degree of vascular calcification described above. Moreover, the area with positive staining for ALP and BMP-2 was significantly increased in IL-37+anti-OPG-treated animals (2.55%±0.76% and 1.57%±0.47%, respectively; P<0.05) compared with the IL-37-treated group, indicating that anti-OPG may affect the expression of ALP and BMP-2 which in turn regulate vascular calcification.
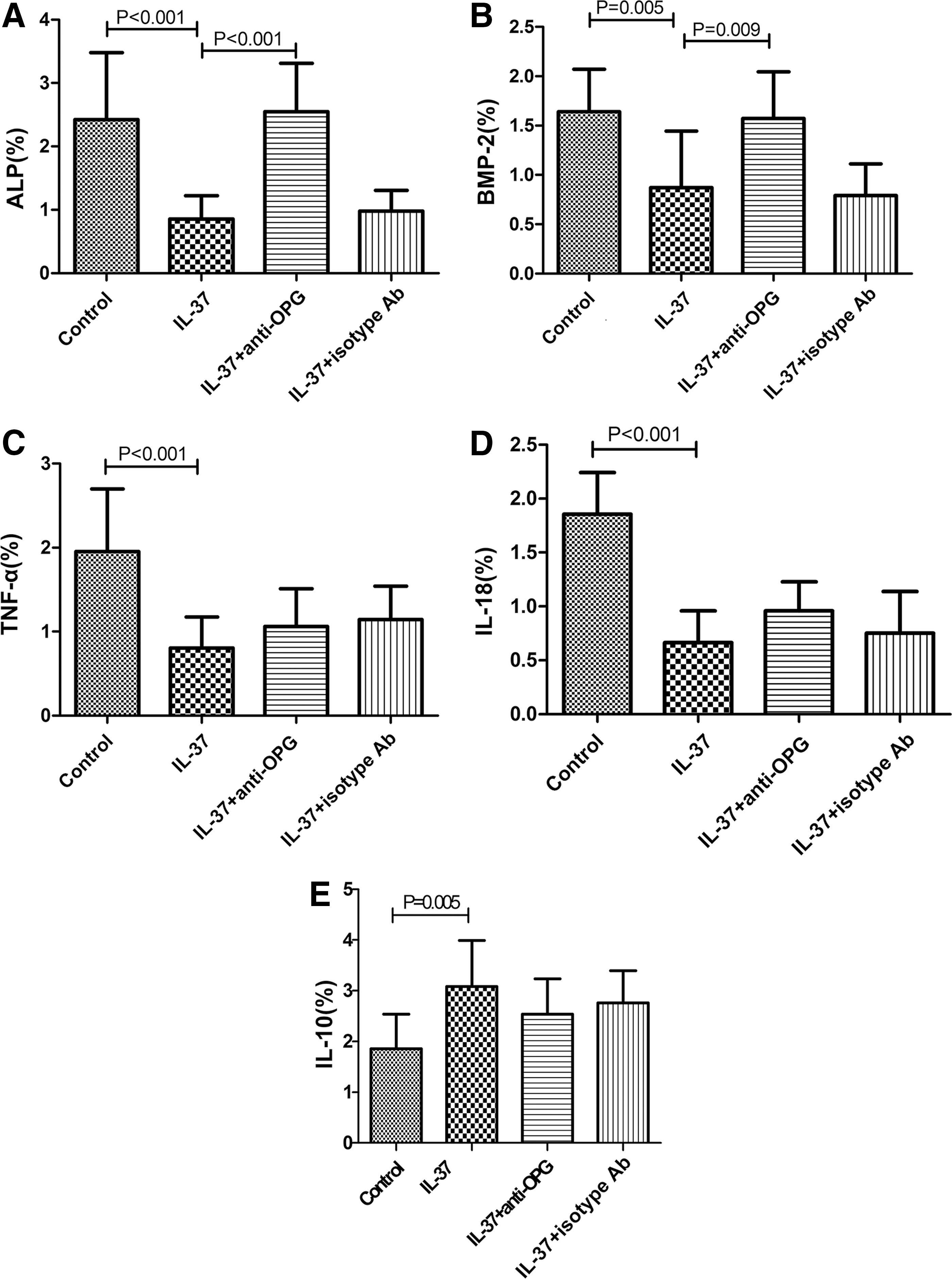
Markers of osteoblast differentiation and atherosclerosis. Immunohistochemical staining of ALP
Effect of IL-37 on the atherosclerotic inflammatory and anti-inflammatory mediators
As inflammatory and anti-inflammatory mediators TNF-α, IL-18, and IL-10 can regulate the process of atherosclerosis, in this study these were examined by staining with immunohistochemical method. We observed that the IL-37-treated group had less areas that stained positive for TNF-α and IL-18 and larger areas staining positive for IL-10, compared with control animals (P<0.05; Fig. 4C–E and Supplementary Table S3). This latter staining showed that IL-37 may inhibit the inflammatory procedure that occurs in atherosclerosis. However, treatment with both IL-37 and anti-OPG could not reverse the percentage change of TNF-α-, IL-18-, and IL-10-positive staining (P>0.05; Fig. 4C–E and Supplementary Table S3), suggesting that anti-OPG may not affect their expression.
Discussion
IL-1 cytokines play a key role in mediating the activation of innate immunity. IL-37, as a newly identified member of the IL-1 family, is present in several organs and tissues, including the heart, brain, kidney, bone marrow, and testis. IL-37 is also expressed in peripheral blood mononuclear cells, monocytes, dendritic cells, and epithelial cell, acting as a natural inhibitor of innate inflammatory and immune responses (Nold and others 2010; Tete and others 2012). Evidence shows that IL-37 plays a protective role in inflammatory and autoimmune diseases in animal models through suppression of the generation of proinflammatory cytokines and the activation of macrophage and dendritic cells (Nold and others 2010; McNamee and others 2011; Sakai and others 2012). However, knowledge of IL-37 in human diseases is rather limited. Nold and others (2010) found that the IL-37 protein is highly expressed in synovial cells of patients with rheumatoid arthritis, but expressed at low levels in healthy human synovial cells. Recently, Boraschi and others (2011) found that IL-37 was expressed in the foam-like cells of atherosclerotic coronary and carotid artery plaques, suggesting that IL-37 is related to the activation of macrophages and the shift in macrophages to foam cells in atherosclerosis. Activated macrophages accumulate in atherosclerotic lesions and play an essential role throughout the whole process of atherosclerosis (Boyle 2005). These studies demonstrated that IL-37 is indeed involved in the disease process of humans. However, the exact role of IL-37 in human diseases remains largely unclear and deserves thorough investigation. Our study presented at this point is the first report that IL-37 can attenuate not only atherosclerosis, but also vascular calcification in mice.
Using an animal model of ApoE−/− mice of diabetes, we were able to demonstrate that IL-37 treatment can decrease atherosclerotic area and lower plaque vulnerability and that exogenous IL-37 plays a protective role in the progression of atherosclerosis leading to increased plaque stability. These findings can be corroborated with several data obtained from Boraschi and others (2011) and our recent publication (Wu and others 2014) that reveals the protective role of IL-37 against mouse myocardial I/R injury.
Our study is also the first to report that IL-37 can have a role in the attenuation of the vascular calcification in the in vivo animal model. Several working hypotheses have been proposed in an attempt to explain how vascular calcification occurs. These include the presence of disturbed Ca/Pi balance, inducing factors (Pi, lipids, inflammatory cytokines, and others), cell death, bone remodeling, and loss of inhibition (Speer and Giachelli 2004). Among these, chronic inflammation seems to be an important factor in aberrant soft tissue calcification as the sites of chronic inflammation in the vasculature were shown to become sites of atherosclerotic calcification in mice (Aikawa and others 2007). Proinflammatory factor TNF-α can induce mineralization of calcifying vascular cells in vitro (Tintut and others 2002). In a human clinical trial, the result is similar. Abdelbaky and others (2013) evaluated imaging data obtained from 137 patients aged 1 to 5 years and showed that the sites of focal aortic inflammation are closely associated with subsequent calcification detected by computed tomographic scanning. Several other studies have also demonstrated that inflammatory cytokines are associated with increased coronary artery calcification in patients (Oh and others 2002; Stompór and others 2005). In the present study, we were able to corroborate the essential role of inflammation in atherosclerotic calcification. Moreover, we discovered that IL-37-treated mice had less calcified area than the control group and this indicates that the anti-inflammatory factor IL-37 may have an important protective role in the progress of vascular calcification.
By measuring the expression of the osteoblast differentiation markers within the lesions of IL-37-treated mice, we were also able to show that the decrease in vascular calcification was associated with a decrease of ALP and BMP-2. ALP is a functional phenotypic marker of osteoblasts, and ALP activity is usually used as a molecular indicator for vascular calcification (Whyte 1994; Johnson and others 2006). ALP is expressed highly in medial smooth muscle cells (SMCs) in medial calcification (Shanahan and others 1999), and the classic stimuli of atherosclerotic vascular calcification, including BMP-2 and oxidized LDL, increases ALP activity in cultured VSMCs. BMP-2 is an important molecule in the regulation of bone formation, as well as in vascular calcification. Treatment with BMP-2 resulted in increased calcification of calcifying vascular cells or bovine SMCs in vitro (Chen and others 2006). In addition, as atherosclerotic lesions evolve, BMP is increasingly expressed by a variety of cells in atherosclerotic lesions (Csiszar and others 2005). Li and others (2008) were able to demonstrate that BMP-2 acts through runx2, which induces the expression of type I collagen and ALP. In our study, after treatment with IL-37 the mice were found to reduce calcification accompanied by reduced areas that stain positive with ALP and BMP-2. We speculate that IL-37 may regulate the expression of ALP and BMP-2, although the exact mechanism is not clear.
Moreover, our study also showed that IL-37 may affect the expression of IL-10, IL-18, and TNF-α. IL-10 is a pleiotropic cytokine produced by a variety of immune cells that can inhibit a broad array of immune and inflammatory responses (Liu and others 2006). IL-10 has an anti-inflammatory property and is thought to play a potentially protective role in the process of atherosclerosis. IL-18 is also a member of IL-1 family, and recent animal experiments support the concept that IL-18 participates in the pathogenesis of atherosclerosis (Elhage and others 2003). TNF-α is a pleiotropic cytokine that exerts potent proinflammatory effects in atherosclerosis and other metabolic and inflammatory disorders. The involvement of TNF-α in the pathogenesis of atherosclerosis is supported by many animal studies (Kober and others 2007). In our present work, exogenous IL-37 upregulates the expression of IL-10, but decreases the expression of IL-18 and TNF-α. Thus, we can speculate that IL-37 may participate in regulating inflammatory and anti-inflammatory mediators, which lead to the attenuation of atherosclerosis and vascular calcification.
Interestingly, in our present study we observed a significant increase of plasma OPG level in the IL-37-treated mice. We subsequently included OPG in a study group to confirm our hypothesis that the protective effect of IL-37 is closely related with the increase of OPG. Indeed, when an anti-OPG antibody is administered, the protective effect of IL-37 in mice with vascular calcification was significantly reduced, accompanied by increased expression of ALP and BMP-2. Compared with the IL-37-treated group, the addition of anti-OPG resulted in increase of both the plaque size of the atherosclerotic lesions and the vulnerability index. However, there were no significant changes in the expression of TNF-α, IL-18, and IL-10. The IL-37+anti-OPG group also showed attenuated atherosclerotic lesions compared with control, indicating that anti-OPG may only partially, not completely, prevent the protective effect of IL-37 on atherosclerosis. This may indicate that the well-known OPG/receptor activator of NF-kB (RANK)/RANK ligand (RANKL) system, which influences bone remodeling and vascular calcification, may have more effect on calcium formation than on atherosclerosis. As we found previously that the levels of IL-37 correlated positively with the increased IL-18 levels in patients with acute coronary syndrome (Ji and others 2014), the protective role of IL-37 may be also related, in part, to other factors such as IL-18.
Several other studies have shown that selective deletion of OPG in mice lead to early onset severe osteoporosis as well as significant calcification of the aorta and renal arteries (Price and others 2001). In addition, inactivation of OPG in apoE knockout mice resulted in increased size of atherosclerotic lesions compared with OPG+/+apoE−/− mice (Bennett and others 2006). Whereas in humans, a large body of epidemiological evidence has suggested a positive association between serum OPG and cardiovascular events such as atherosclerosis and arterial calcification (Jono and others 2002). It is likely that the elevated serum concentration of OPG is simply a result of the calcification through some unknown mechanism, or the serum concentrations of OPG might not reflect OPG levels in the vessel wall at all. Although recently the anti-RANKL antibody failed to have any treatment outcomes of arterial calcification and other cardiovascular disease in human (Samelson and others 2014), IL-37 therapy may have a chance to challenge this clinical difficulty through OPG.
In conclusion, our findings clarify the protective effect of IL-37 in the process of the vascular calcification and atherosclerosis. Although the mechanism of IL-37 is yet not clear and needs further investigation, we believe that the protective effect of IL-37 partly dependents on the activity of OPG as shown in our present study. In addition, significant changes in disease-relevant cytokines (eg, ALP, BMP-2, TNFa, IL-18, and IL-10) were also found after IL-37 treatment. Therefore, it is reasonable to hypothesize that IL-37 may offer a new therapeutic route to solve the difficulty of clinical therapy of calcification and atherosclerotic disease.
Footnotes
Acknowledgments
This work was supported by the Beijing Municipal High-Level Talent Foundation of Health System (No. 2011-1-5), Beijing Municipal Administration of Hospitals Clinical Medicine Development of Special Funding Support (ZY201303), the National Key Clinical Specialty Construction Project, the National Natural Science Foundation of China (No. 81160045 and 81270285), and the Chinese Postdoctoral Science Foundation Grant (No. 2013 M540987).
Author Disclosure Statement
The authors declare that they have no competing interests.