
Abstract
Determining to what extent biophysical characteristics of aggregates affect immunogenicity of therapeutic interferon beta-1b. Three recombinant human interferon beta-1b (rhIFNβ-1b) samples with different levels of aggregates generated by copper oxidation, thermal stress, or left untreated, as well as Avonex® drug substance and Betaferon® drug product, were injected intraperitoneally in nontransgenic and interferon beta transgenic FVB/N mice 5 times per week for 3 weeks. Antibodies against interferon beta were measured using enzyme-linked immunosorbent assay. UV and fluorescence spectroscopy, dynamic light scattering, size exclusion chromatography, reversed-phase high-performance liquid chromatography (RP-HPLC), fluid imaging microscopy, and resonant mass measurement, as well as sodium dodecyl sulfate–polyacrylamide gel electrophoresis and Western blotting, were used to characterize and quantitate aggregates in the 3 rhIFNβ preparations, to correlate biophysical characteristics with immunogenicity. In immune-tolerant interferon beta transgenic FVB/N mice, Betaferon drug product showed the highest immunogenicity, while Avonex drug substance showed the lowest level of immunogenicity. Of the 3 forms of rhIFNβ-1b, copper-oxidized rhIFNβ-1b showed lower immunogenicity than thermally stressed rhIFNβ-1b, despite containing larger aggregates. Both copper-oxidized rhIFNβ-1b and thermally stressed rhIFNβ-1b exhibited changes in protein structure as shown using fluorescence spectroscopy and RP-HPLC. Nontransgenic, nonimmune-tolerant FVB/N mice generated high antibody titers against all interferon beta samples tested. The level of immunogenicity and the breaking of tolerance in FVB/N transgenic mice are not only related to the level of aggregation but also depend on the size and structure of the aggregates.
Introduction
D
Recombinant human interferon beta-1b (rhIFNβ-1b) is produced in Escherichia coli, is nonglycosylated, has a Cys to Ser mutation at position 17, and the N-terminal methionine is missing. Furthermore, rhIFNβ-1b contains ∼60% noncovalent aggregates (van Beers and others 2011).
rhIFNβ-1b has a molecular weight of 18.5 kDa and consists of a single polypeptide chain of 165 amino acids. The protein structure contains 5 α-helices and 1 intramolecular disulfide bond between Cys31 and Cys141 (Karpusas and others 1997, 1998). The absence of the attached carbohydrate chain that is found in rhIFNβ-1a and the subsequent reduction in solubility were proposed to result in aggregation and an increased immunogenicity of rhIFNβ-1b compared to rhIFNβ-1a (Karpusas and others 1997, 1998). The presence of aggregates not only enhances its immunogenicity in transgenic immune-tolerant mice but also as a result diminishes the biological activity of the protein in vivo (Hermeling and others 2005b; van Beers and others 2010b, 2010c). While there is relatively little known about the aggregation state of therapeutic proteins following parenteral administration, studies have suggested that aggregates can be modified after exposure to biological fluids (Filipe and others 2012).
The immunogenicity of a therapeutic protein can also be elevated by chemical alteration, such as hydrolysis, degradation, deamidation, or oxidation (Patro and others 2002; Sharma 2007). Oxidation of therapeutic proteins can occur following exposure to light, metal ions, or peroxide impurities during formulation, manufacturing processes, lyophilization, and storage (Lam and others 1997; Patro and others 2002; Sharma 2007). Bioactivity of functional proteins can be changed by oxidation (Shacter 2000). Novel epitopes can be formed by oxidation, and chemical modification may lead to alterations in immunogenicity (van Beers and others 2010a). Indeed, metal-catalyzed oxidized rhIFNα increased immunogenicity in transgenic mice due to aggregation (Hermeling and others 2006). Similarly, aggregation and precipitation of human relaxin occurred as a consequence of metal-catalyzed oxidation (Li and others 1995; van Beers and others 2010b).
In this study, we studied the effect of 2 different stress methods on biophysical characteristics of the resulting aggregates of rhIFNβ-1b and the immunogenicity of these forms in FVB/N immune-tolerant mouse model, which had been determined as a suitable model to study rhIFNβ products (Haji Abdolvahab and others 2014a), and compared to untreated rhIFNβ-1b. Untreated rhIFNβ-1b contained mostly monomers with low levels of aggregates and no quantifiable oxidized forms.
In addition, Avonex® drug substance (a low immunogenic control protein) (Hermeling and others 2005b; van Beers and others 2010a), and Betaferon® drug product (a high immunogenic control protein) (Hermeling and others 2005b; van Beers and others 2010a) were tested in FVB/N immune-tolerant mice as controls.
The results show that aggregation can enhance immune responses against rhIFNβ-1b associated with the shape, size, and structure of the aggregates.
Materials and Methods
rhIFNβ products
Avonex drug substance (bulk rhIFNβ-1a) was obtained from Biogen Idec, Inc. as a 0.27 mg/mL solution in 200 mM sodium chloride and 100 mM sodium phosphate buffer at pH 7.2. Betaferon drug product was purchased from BAYER as the lyophilized powder containing 0.25 mg/mL of rhIFNβ-1b, 12.5 mg/mL of mannitol, and 12.5 mg/mL of human serum albumin and reconstituted in phosphate-buffered saline (PBS) before use. rhIFNβ-1b was supplied by Zistdaru Danesh Co. Ltd. as a 1.96 mg/mL frozen solution in 40 mM sodium hydroxide (pH 10). Before testing, it was formulated in 10 mM sodium acetate buffer at pH 7.4 from Merck (64271), 200 mM
Physicochemical characterization
Visual inspection
Untreated, copper-oxidized, and thermally stressed rhIFNβ-1b solutions (0.25 mg/mL) were examined visually against a black background and compared with the sample buffer as a reference.
UV spectroscopy
UV spectra (λ = 250–360 nm) of 250 μg/mL IFNβ-1b solutions were recorded at room temperature in 1 cm path length quartz cuvettes using a UV-2450 UV/VIS spectrophotometer (Shimadzu). Ten millimolar sodium acetate buffer (pH 7.2) was used as a blank.
Dynamic light scattering
Dynamic light scattering (DLS) was used to measure an intensity-based average diameter (Z-ave) and polydispersity index (PDI) of particles in rhIFNβ-1b solutions. Two hundred fifty micrograms per milliliter rhIFNβ-1b was analyzed using a Malvern ALV CGS-3 goniometer (Malvern Instruments) equipped with a red laser source (λ = 632.8 nm) and a detector at a 90° angle. Samples were measured at 25°C in the presence or absence of 0.01% (w/v) sodium dodecyl sulfate (SDS).
The DLS time correlation of the samples was analyzed using ALV Correlator 3.0 software (ALV).
Fluorescence spectroscopy
rhIFNβ-1b samples (250 μg/mL) were excited at 295 nm and fluorescence emission spectra were recorded from 305 to 450 nm in 1 cm path length quartz cuvettes. A Horiba Fluorolog fluorometer (Horiba Jobin Yvon) was used. Integration time per data point was 1 s and slits were set at 5 nm.
Resonant mass measurement
Resonant mass measurement (RMM; Affinity Biosensors) equipped with a “micro” format resonant mass sensor (channel cross section 8 × 8 μm2, resonant frequency 400 kHz) was used to analyze particles smaller than 5 μm (Burg and others 2007). The sensor was flushed for 60 s with milli-q water before every run. Consequently, possible impurities in the system were removed by 2 “sneeze” operations followed by another flushing with milli-q water for 60 s. Then, 150 μL of sample filtered using 0.2 μm filter (GE Healthcare), was loaded into the sensor for 45 s, and each sample was measured in triplicate, obtaining values for 500 particles in each measurement. Particle Lab software version 1.8.510 (Affinity Biosensors) was used to discriminate silicon oil from protein aggregates and define the total number of protein aggregates.
Fluid imaging microscopy (MFI)
A fluid imaging microscopy (MFI) system (MFI5200; ProteinSimple) equipped with a 100 μm silane-coated flow cell and controlled by MFI View System Software (MVSS) version 2 was used. The system was flushed with 5 mL Milli-Q water at maximum flow rate and the cleanliness of the background was inspected visually. The background was zeroed by flowing PBS and performing the “optimize illumination” procedure. Samples of 0.5 mL with a prerun volume of 0.2 mL were analyzed at a flow rate of 0.17 mL/min and a fixed camera rate of 22 flashes per second.
The data recorded by the MVSS were analyzed with MFI View Analysis Suite (MVAS) version 1.2. For each sample, stuck, edge, and slow moving particles were removed by the software before data analysis. The equivalent circular diameter, which is the diameter of a circle that has an area equal to that of the particle imaged by MFI, was calculated and presented as a measure of the particle size.
Size-exclusion ultra-performance liquid chromatography
rhIFNβ-1b samples (7.5 μL, 250 μg/mL) were characterized with a Waters ACQUITY UPLC system (Waters). A flow rate of 0.25 mL/min was operated using a Waters AQUITY BEH450 SEC (150 × 4.6 mm, 2.5 μm) ultra-performance liquid chromatography (UPLC) column with a total run time of 14 min. The mobile phase contained 50 mM sodium acetate at pH 7.4, 200 mM
Reversed-phase high-performance liquid chromatography
A Jupiter 300 (5 μm, 250 × 4.6 mm) C4 column was used in combination with a Security Guard (4 × 3 mm) C4 guard column (Phenomenex) to analyze rhIFNβ-1b (20 μL, 250 μg/mL) preparations. A Waters Alliance 2695 system in line with a Waters 2475 Fluorescence detector (excitation at 295 nm/emission at 340 nm) was used. The sample temperature was 20°C and the column temperature was set at 30°C. The mobile phases were 10% acetonitrile, 0.1% trifluoroacetic acid (TFA) and 100% acetonitrile, 0.1% TFA, and protein was eluted with a gradient according to Geigert and others (1988) run at 1 mL/min.
Sodium dodecyl sulfate polyacrylamide gel electrophoresis
Five micrograms of sample solutions was mixed with 4× sample buffer under reducing and nonreducing conditions and applied to precast sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE) gels (Ready Gel, Tris-HCl, 4–12% NUPAGE®; Life Technologies) (10 wells × 1.5 mm) at 70 V for 30 min, then the voltage was increased to 150 V for 1 h. Prestained broad range molecular weight markers (Biorad) were run as standards. Visualization of protein bands was carried out using silver staining followed by densitometry using a Biorad GS-800 densitometer and Quantity One software.
Western blotting
Protein samples separated using SDS-PAGE were transferred to nitrocellulose using the iBlot® Dry Blotting System (Life Technologies). The transfer buffer consisted of 10 mM sodium hydrogen carbonate, 3 mM sodium carbonate, 0.1% SDS (w/v), and 20% (v/v) methanol at pH 10.0. Blots were blocked with 5% (w/v) nonfat milk powder (ELK; Campina Melkunie) in 0.005% (w/v) Tween 20 in PBS for 1 h with constant orbital shaking. After washing with Tris-buffered saline containing 0.005% (v/v) Tween 20 (TBS-T), blots were incubated with 0.1 μg/mL polyclonal rabbit anti-IFNβ antibody (Acris Antibodies) in 5% (w/v) bovine serum albumin and 0.005% (w/v) Tween 20 in PBS (dilution buffer) with constant orbital shaking for 2 h. Blots were washed with the TBS-T buffer and then incubated with peroxidase-labeled goat anti-rabbit immunoglobulin G (IgG; Sigma Aldrich), diluted 10,000-fold in a dilution buffer for 1 h with constant orbital shaking. Blots were washed and protein bands were visualized using SuperSignal West Femto Chemiluminescent Substrate (Thermo Scientific) and a Gel Doc Luminescent Imaging system (BIORAD Chemdox XRS). Data were analyzed with Quantity One software.
Immunogenicity study
Mouse breeding
Human IFNβ transgenic immune-tolerant FVB/N mice, developed previously (Haji Abdolvahab and others 2014a), were bred at the Central Laboratory Animal Institute (Utrecht University, The Netherlands). Transgenic mice were crossed with wild-type FVB/N mice (Janvier; Bioservices) to maintain the strain. The offspring was genotyped by polymerase chain reaction using chromosomal DNA of ear tissue to confirm the presence of the hIFNβ gene in their genome. As mice were not checked if they expressed the hIFNβ gene, they were genotypically transgenic, but not necessarily phenotypically transgenic. Both transgenic FVB/N mice and nontransgenic littermates were used.
Animal experiment
The animal experiments were permitted by the Institutional Ethics Committee (Utrecht University, The Netherlands). Water (acidified) and food (Hope Farms) were provided ad libitum. Fifty transgenic and 50 nontransgenic mice were injected intraperitoneally with 5 μg of Avonex drug substance, Betaferon drug product, untreated rhIFNβ, thermally stressed rhIFNβ, and copper-oxidized rhIFNβ using a schedule of 5 injections/week for 3 weeks (on days 1–5, days 8–12, and days 15–19) (Hermeling and others 2005b; van Beers and others 2010b, 2010c, 2011). Before and during the treatment, blood was drawn from a cheek puncture using an interval of at least 7 days between sampling (Hermeling and others 2005b; van Beers and others 2010c). At the end of the experiment, all mice were sacrificed by decapitation under isoflurane anesthesia and the final blood was collected. Blood samples were centrifuged for 10 min at 3,000 g in lithium heparin gel tubes and plasma was stored at −80°C.
Enzyme-linked immunosorbent assay
Plates were coated with 1 μg/mL Avonex drug substance in PBS (100 μL/well) overnight at 4°C. The plasma was analyzed using enzyme-linked immunosorbent assay based on a protocol designed by Hermeling and others (2005b) in Microlon 96-well plates (Greiner).
Plasma samples were screened and determined to be positive if the background absorbance values of 100-fold diluted samples were 10 times more than the value of the pretreatment plasma. The absorbance of serial dilutions against log dilution was plotted to a sigmoidal dose–response curve using GraphPad Prism version 4.02 (GraphPad software) to define titers of anti-hIFNβ IgG in positive samples. The equivalent of the dilution of the EC50 value was defined as the titer of the plasma. Statistical significance was calculated with a nonparametric Kruskal–Wallis one-way analysis of variance to compare different treatment groups and Dunn's multiple comparisons test was performed to compare the mean rank of each treatment group with every other group.
Results
Characteristics of rhIFNβ-1b
Visual inspection
The untreated and thermally stressed rhIFNβ-1b solutions were transparent without any noticeable aggregation or precipitation, while the copper-oxidized rhIFNβ-1b had an intense homogenous white haze with some visible particles detected.
UV spectroscopy
The UV spectra of all samples tested showed a peak at 280 nm, consistent with the presence of aromatic amino acid residues within the sequence of rhIFNβ-1b. An increase in light scattering due to protein aggregation can be evaluated using UV spectroscopy (Kueltzo and Middaugh 2005), the magnitude of which is determined by the shape, size, and the number of aggregate particles. To analyze the aggregation of rhIFNβ-1b, the optical density (OD) at 350 nm and the ratio OD 280 nm to OD 260 nm were determined (Table 1). A higher OD350 and a lower OD280:OD260 is indicative of aggregation (van Beers and others 2010b, 2011). The UV spectrum of copper-oxidized rhIFNβ-1b exhibited the highest light scattering due to the visible particles in the solution (Fig. 1). In the absence of SDS, both the copper-oxidized and thermally stressed samples showed increased OD350 values (0.78 and 0.18, respectively) compared to the untreated protein (0.004). The OD280:OD260 was 1.24 for thermally stressed rhIFNβ-1b, which was lower than the ratio for the untreated protein (2.32).
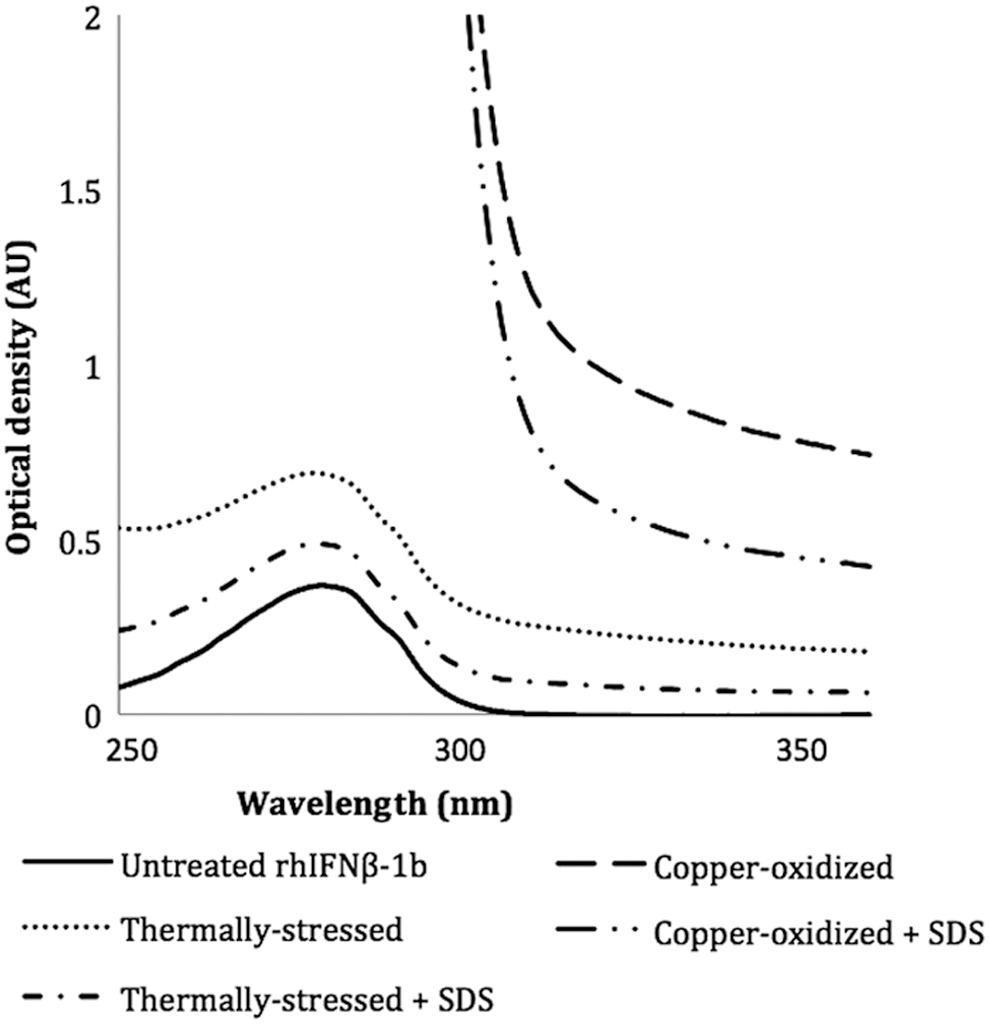
UV spectra of untreated, thermally stressed, and copper-oxidized recombinant human interferon beta-1b (rhIFNβ-1b) in the presence and absence of 0.01% (w/v) sodium dodecyl sulfate (SDS).
0.01% (w/v) SDS was added to the rhIFNβ-1b preparations before analysis.
SDS was not added to the untreated rhIFNβ-1b, which contained DDM.
OD, optical density; SDS, sodium dodecyl sulfate; DLS, dynamic light scattering; PDI, polydispersity index; MFI, fluid imaging microscopy; SE-UPLC, size-exclusion ultra-performance liquid chromatography; rhIFnβ-1b, recombinant human interferon beta-1b; DDM, N-dodecyl-β-
The thermally stressed preparation showed a decrease in OD350 from 0.18 to 0.06 and an increase in OD280:OD260 from 1.24 to 1.59 in the presence of 0.01% (w/v) SDS (Fig. 1; Table 1). Copper-oxidized rhIFNβ-1b also showed a decrease in OD350 in the presence of 0.01% (w/v) SDS; from 0.78 to 0.44 (Fig. 1; Table 1). The ability of SDS to reduce the OD350 values of the thermally stressed and copper-oxidized samples indicated the presence of noncovalent aggregates. However, as the UV scans of the thermally stressed and copper-oxidized samples in the presence of SDS did not overlay that of the untreated rhIFNβ-1b, covalent aggregates, possibly linked by disulfide bonds, are likely to also have been present. The copper-oxidized sample contained a higher level of covalent aggregates compared to the thermally stressed protein as shown by the higher OD350 in the presence of SDS (0.44) for copper-oxidized rhIFNβ-1b versus 0.06 for the thermally stressed rhIFNβ-1b). Since micellar solutions of mixed surfactants can display particularly different physicochemical behavior from those of single surfactants (Turro and others 1986), SDS was not added to the untreated rhIFNβ-1b which contained DDM.
Dynamic light scattering
DLS was used to measure the hydrodynamic size average of protein particles in the range of 1 nm to 5 μm (den Engelsman and others 2011). Samples were each measured 4 times and Z-ave and PDI values are shown in Table 1. Untreated rhIFNβ-1b showed the lowest Z-ave value (48 ± 2 nm) compared to the thermally stressed (521 ± 28 nm) and copper-oxidized (7,949 ± 525 nm) rhIFNβ-1b, reflecting the presence of aggregates in the treated samples. Untreated rhIFNβ-1b contained mostly monomeric protein as the size distribution of 95% of the species detected was ∼5 nm in diameter (Supplementary Fig. S1; Supplementary Data are available online at
The addition of 0.01% (w/v) SDS to thermally stressed rhIFNβ-1b resulted in a decrease in Z-ave (from 521 ± 28 to 185 ± 3 nm) and PDI (from 0.7 ± 0.1 to 0.5 ± 0.03), indicating the dissociation of noncovalent protein aggregates. Similarly, the aggregates present in copper-oxidized rhIFNβ-1b dissociated in the presence of 0.01% (w/v) SDS with a concomitant reduction in the Z-ave value from 7,949 ± 525 to 1,304 ± 161 nm; however, the PDI value increased from 0.3 ± 0.03 to 1 ± 0. This increase in polydispersity, despite the reduction of the Z-ave value, indicates that the aggregates in the copper-oxidized protein are more heterogeneous than following SDS treatment of thermally stressed protein.
Fluorescence spectroscopy
Fluorescence spectroscopy was used to probe the structure of the untreated, thermally stressed, and copper-oxidized sample of rhIFNβ-1b by measuring the fluorescence of the tryptophan residues located at positions 22, 79, and 143. Changes in fluorescence (intensity and wavelength of the peak) indicate changes in the microenvironment of these residues that are typically buried within protein structures (Karpusas and others 1998; Runkel and others 2000). Fluorescence spectra following excitation at 295 nm are shown in Fig. 2. The maximum emission intensity of untreated rhIFNβ-1b was set at 1.00, and the relative intensities of the thermally stressed and copper-oxidized samples are shown in Table 1. The untreated rhIFNβ-1b exhibited a maximum emission at 338 nm. The fluorescence intensity of the thermally stressed protein was 4-fold lower compared with the untreated sample, while the wavelength at maximal emission was unchanged. By contrast, the copper-oxidized protein had a relative fluorescence intensity that was 50-fold lower compared with the untreated sample and showed a red-shift in the wavelength at maximal fluorescence of 2 ± 0.5 nm indicating a slightly more hydrophilic environment of at least one of the tryptophans (Karpusas and others 1998; Runkel and others 2000) (Fig. 2). By contrast to the thermally stressed and copper-oxidized rhIFNβ-1b, the untreated protein likely maintained most of its native tertiary structure (Fig. 2).
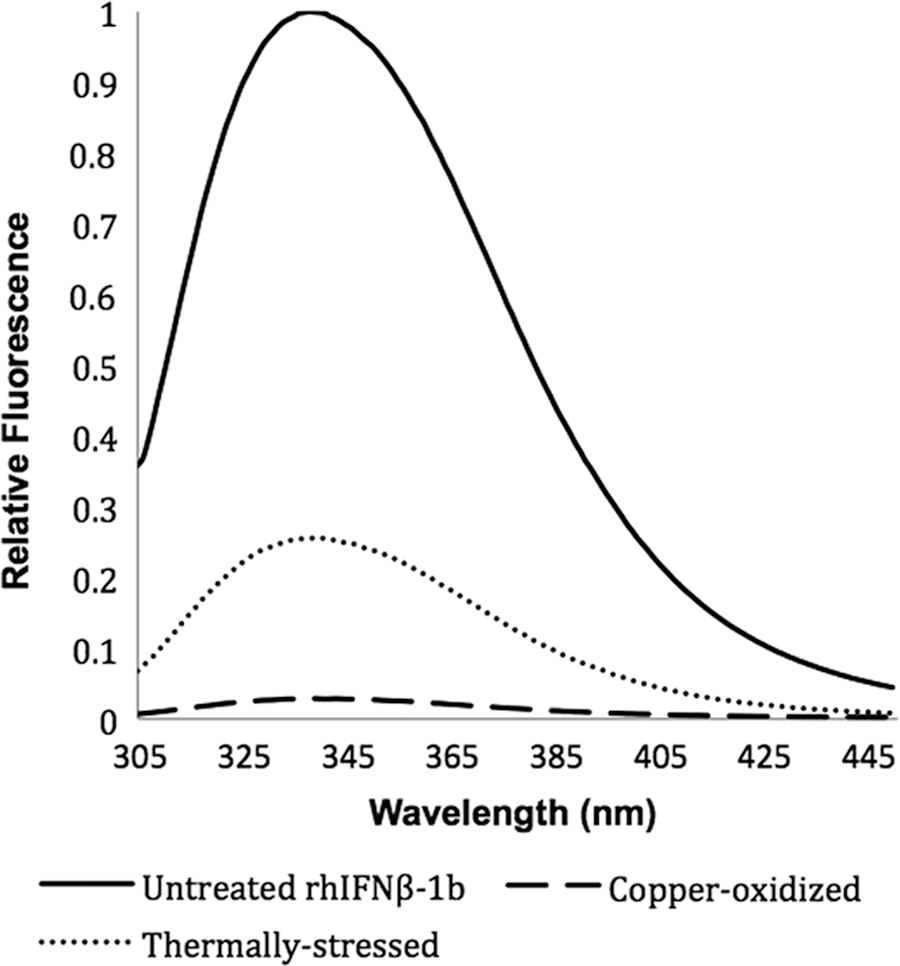
Fluorescence emission spectra of untreated, thermally stressed, and copper-oxidized rhIFNβ-1b.
Resonant mass measurement
Submicron and micron particles with a size range from 20 nm up to about 5 μm were measured in untreated, thermally stressed, and copper-oxidized rhIFNβ-1b using RMM. Untreated rhIFNβ-1b showed a low total particle concentration (7 × 105 particles/mL) that was all within the size range of 0.2–0.4 μm (Fig. 3). Thermally stressed protein contained heterogeneous particles ranging from 0.2 to larger than 2 μm, with the most prevalent being in the size range of 0.9–1.2 μm. The copper-oxidized protein showed a lower number of total particles in the 0.2–2 μm size range than the thermally stressed protein, with a particle concentration that was ∼7-fold lower compared to the thermally stressed protein; 55.2 × 105 particles/mL for copper-oxidized protein versus 374 × 105 particles/mL for the thermally stressed protein (Fig. 3).
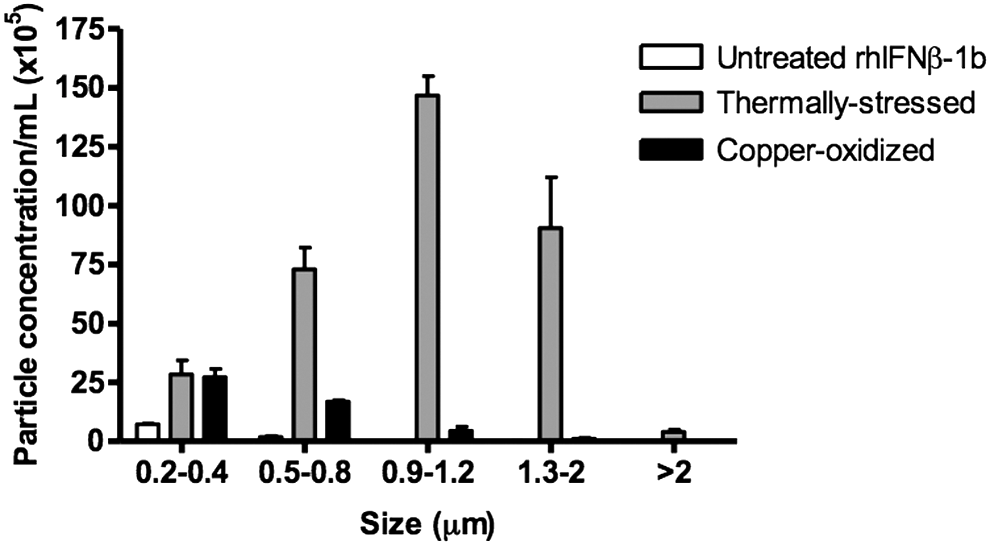
Particle size distribution ranging from 0.2 to >2 μm of untreated, thermally stressed, and copper-oxidized rhIFNβ-1b determined using resonant mass measurement. Error bars represent standard deviations from triplicate measurements.
Fluid imaging microscopy
MFI is a technique that can be used to determine the numbers of particles from 1 to 100 μm in size (Sharma and others 2010). Untreated rhIFNβ-1b showed a low particle concentration (0.18 × 105 particles/mL), slightly higher than the PBS control (0.05 × 105 particles/mL), whereas the particle concentrations of thermally stressed and copper-oxidized rhIFNβ-1b were significantly higher (160 × 105 and 30 × 105 particles/mL, respectively) (Table 1). The protein particles in the copper-oxidized sample were heterogeneous and ranged from 1 to >41 μm in size (Fig. 4) and in shape; small compact fibers to large ribbon-like aggregates (Fig. 5). The majority of particles in the thermally stressed rhIFNβ-1b were 1–2 μm in size, with the remainder being 3–12 μm (Fig. 4). A low number of particles were observed in untreated rhIFNβ-1b in the 1–2 μm size range, with minimal numbers in the 3–8 μm size range (Fig. 4). Particles in the untreated rhIFNβ-1b sample had a mean size of 2.02 ± 2.34 μm, while particles of thermally stressed and copper-oxidized protein had a mean size of 1.82 ± 1.15 μm and 5.36 ± 6.49 μm, respectively (Table 1). Thus, the copper-oxidized sample contained larger particles compared to the thermally stressed protein. Whole-screen images of untreated, thermally stressed, and copper-oxidized rhIFNβ-1b using MFI are shown in Supplementary Fig. S2. The copper-oxidized rhIFNβ-1b contained significantly larger aggregates compared to the thermally stressed protein.
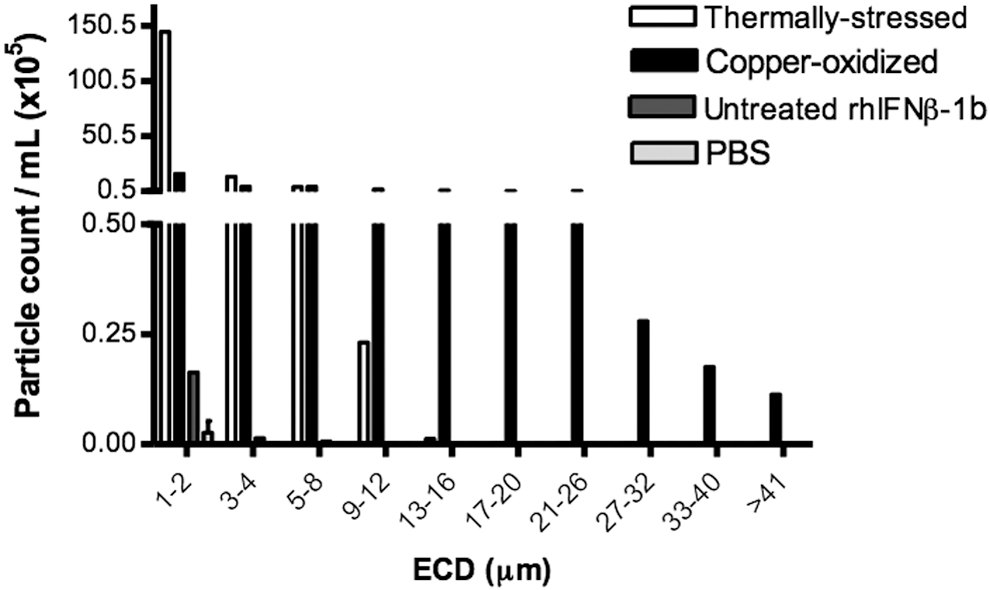
Protein particles (size range from 1 to >41 μm) detected using fluid imaging microscopy (MFI) in untreated, thermally stressed, and copper-oxidized rhIFNβ-1b. Phosphate-buffered saline (PBS) was analyzed as a control.

MFI images of protein particles detected in samples of untreated, thermally stressed, and copper-oxidized rhIFNβ-1b. PBS was analyzed as a control. The calculated dimensions refer to the equivalent circular diameter of each detected particle.
Size-exclusion ultra-performance liquid chromatography
Size-exclusion ultra-performance liquid chromatography (SE-UPLC) was carried out with 0.1% (w/v) DDM in the mobile phase to reduce nonspecific interactions of the protein with the column resin (Hawe and Friess 2008). The SE-UPLC method enabled monomers and covalent or tight noncovalent multimers of rhIFNβ-1b to be characterized (Fig. 6); weakly bound noncovalent aggregates may not be measurable using this method as they may dissociate in the presence of the DDM, which has surfactant properties. The chromatogram of the untreated rhIFNβ-1b showed 2 peaks, a dimeric peak (peak 1 of Fig. 6, mass ∼37 kDa obtained from calibration curve) representing 3% of the total area under the curve and a monomeric species (peak 2 of Fig. 6, mass ∼18.5 kDa) representing 97% of the total area under the curve (Fig. 6; Table 1). Thermally stressed rhIFNβ-1b also showed 2 peaks consistent with dimeric (52%) and monomeric (48%) forms, but the total recoverable protein was only 60% of that of the untreated rhIFNβ-1b. The missing 40% was likely composed of large aggregated forms removed from the sample during the filtration step before the SE-UPLC analysis (Fig. 6; Table 1). The presence of 0.1% (w/v) DDM in the mobile phase may have caused dissociation of noncovalent aggregates and therefore only covalent dimers (potentially linked by disulfide bonds) were detected. For the copper-oxidized protein, no recoverable protein was detected, presumably due to complete loss of the aggregated forms during the filtration step that preceded the SE-UPLC analysis (Fig. 6; Table 1).
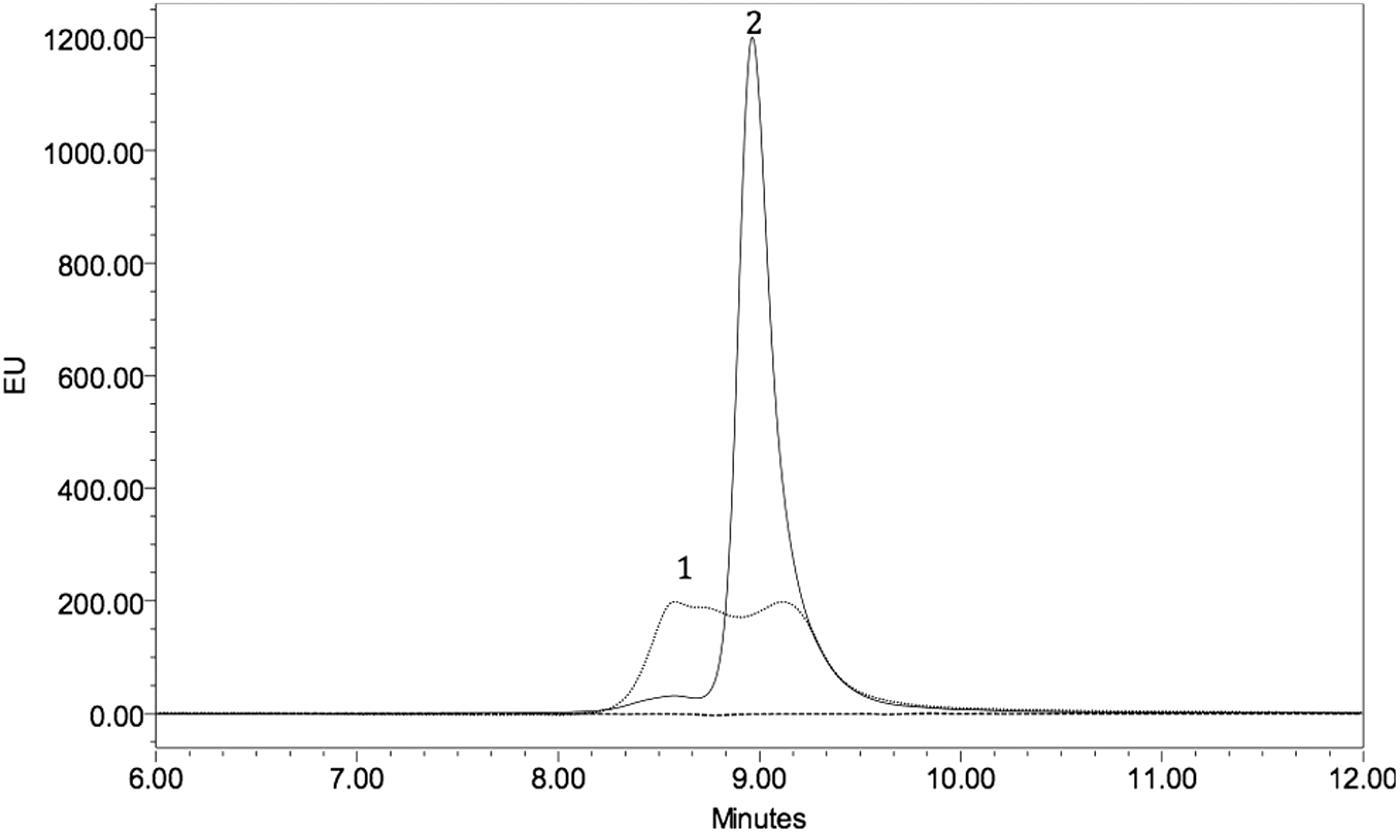
Size-exclusion ultra-performance liquid chromatography chromatograms of untreated (solid line), thermally stressed (dotted line), and copper-oxidized (dashed line) rhIFNβ-1b. The peaks were identified using fluorescence detection. Peak 1 corresponds to a dimeric form, while peak 2 corresponds to the monomeric form. Both dimeric and monomeric peaks were observed for untreated and thermally stressed rhIFNβ-1b, whereas no protein was recovered in the sample of copper-oxidized rhIFNβ-1b. A partial loss of protein (40%) and relatively 49% increase in dimer and 49% decrease in monomer peak were observed in thermally stressed rhIFNβ-1b compared to that for the untreated protein.
Reversed-phase high-performance liquid chromatography
Reversed-phase high-performance liquid chromatography (RP-HPLC) was used to detect potentially oxidized form(s) of rhIFNβ-1b [methionine sulfoxide derivative(s)] that should elute before nonoxidized protein due to increased hydrophilicity (Brown 2011; van Beers and others 2011). Using this method, untreated rhIFNβ-1b was found to contain no detectable oxidized form(s) (Fig. 7). RhIFNβ-1b treated with copper eluted at a position corresponding to oxidized rhIFNβ-1b and showed a total protein recovery of 64% compared to that of untreated rhIFNβ-1b. Thermally stressed rhIFNβ-1b eluted between the peaks of untreated and copper-oxidized protein (Fig. 7), potentially due to partial oxidation and/or Trp22, which is exposed to solvent in the native structure, becoming buried during aggregate formation and thereby lowering overall hydrophobicity (Fan and others 2005). As for the copper-oxidized rhIFNβ-1b, recovery of the thermally stressed protein was <100%.

Reversed-phase high-performance liquid chromatography chromatograms of untreated (solid line), thermally stressed (dotted line), and copper-oxidized (dashed line) rhIFNβ-1b. Peaks are O, copper oxidized; T, thermally-stressed; and N, untreated rhIFNβ-1b.
Sodium dodecyl sulfate–polyacrylamide gel electrophoresis
SDS-PAGE under reducing and nonreducing conditions (Walker 2002) was carried out to investigate the presence of covalent complexes of rhIFNβ-1b (Fig. 8). Untreated rhIFNβ-1b showed a single band at ∼18.5 kDa under nonreducing (Fig. 8E) and reducing (Fig. 8G) conditions, corresponding to the monomeric form of the protein. Under nonreducing conditions, copper-oxidized and thermally stressed rhIFNβ-1b together showed a number of bands, including monomer, but also higher molecular weight species likely representing dimeric, trimeric, and larger aggregates (Fig. 8A). Under reducing conditions, only monomeric and dimeric forms were detected for copper-oxidized and thermally stressed rhIFNβ-1b (Fig. 8C), indicating that the aggregates larger than dimers were likely disulphide bonded.
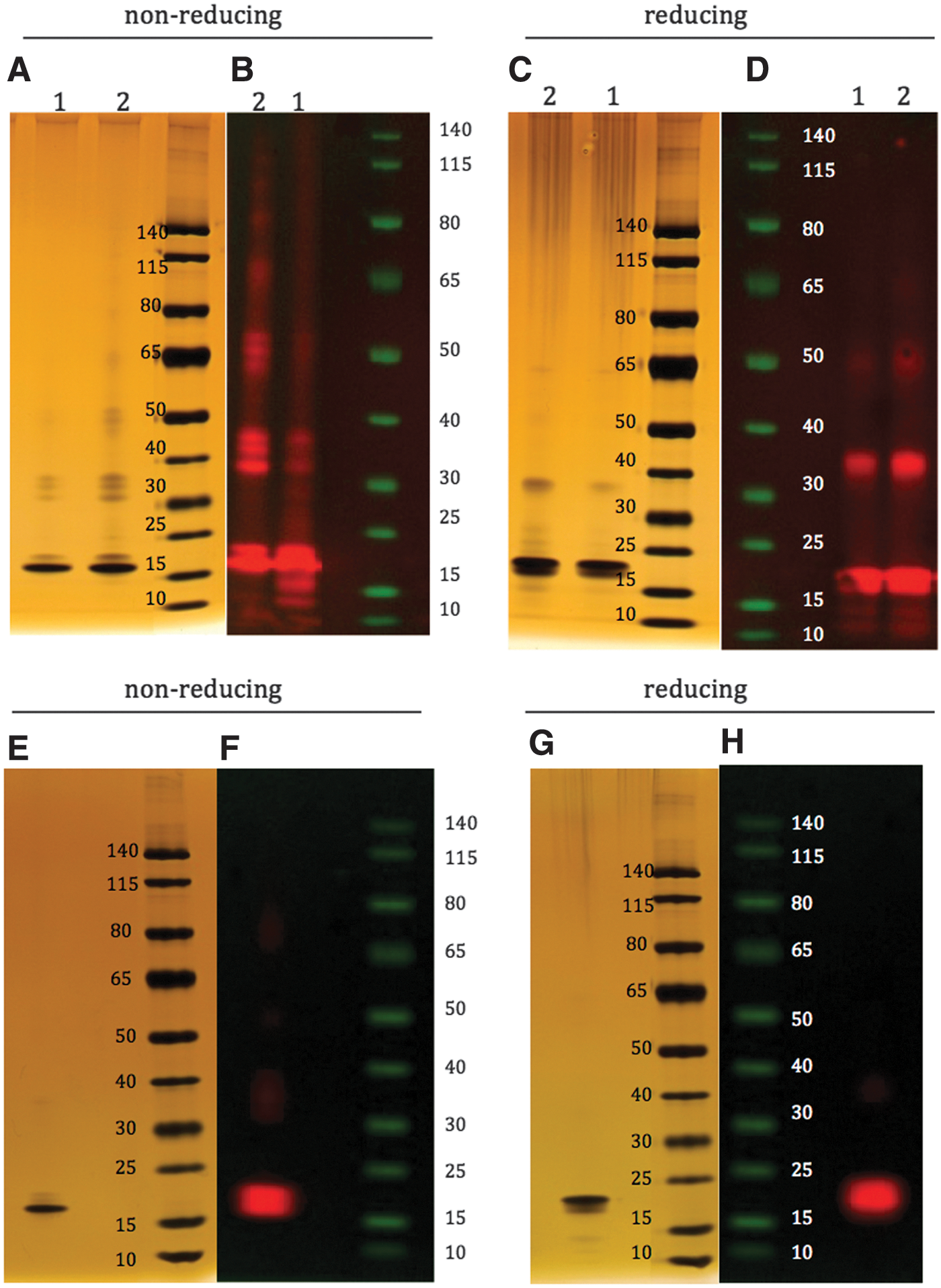
Sodium dodecyl sulfate–polyacrylamide gel electrophoresis under nonreducing and reducing conditions for thermally stressed [
Western blotting
Anti-rhIFNβ-1b antibodies were used to probe untreated, thermally stressed, and copper-oxidized rhIFNβ-1b that had been separated using SDS-PAGE under both nonreducing and reducing conditions. The monomeric form of the protein as well as the multimeric species were clearly observed in thermally stressed and copper-oxidized rhIFNβ-1b. Consistent with the nonreducing and reducing SDS-PAGE data, higher molecular weight forms were detected under nonreducing conditions than under reducing conditions (compare Fig. 8B and D). By contrast, the monomeric form was the predominant form detected in untreated rhIFNβ-1b under both reducing and nonreducing conditions, with very low levels of higher aggregated forms detected (Fig. 8F, H).
Immunogenicity
Antibody responses against untreated, thermally stressed, and copper-oxidized rhIFNβ-1b, as well as against Avonex drug substance and Betaferon drug product, after 3 weeks of treatment (5 times per week) in both transgenic and nontransgenic FVB/N mice were tested as described (Haji Abdolvahab and others 2014a) (Fig. 9). All test articles induced anti-IFNβ antibodies in nontransgenic mice as the human protein is foreign to the murine immune system in this setting (Haji Abdolvahab and others 2014a). All nontransgenic mice exhibited significantly higher immunogenicity compared to transgenic animals administered with the same test article (P < 0.0001). In transgenic mice, 80% of the Avonex drug substance-treated mice did not develop anti-IFNβ antibodies, likely due to the recombinant protein being similar to the human IFNβ expressed by the mice. By contrast, 100% of the transgenic mice administered Betaferon drug product developed anti-IFNβ antibodies, likely due to the test article being different from that of the human IFNβ expressed by the mice, and thereby breaking immune tolerance. These results are consistent with the relative levels of neutralizing anti-IFNβ antibodies formed in multiple sclerosis patients administered Avonex or Betaferon drug products; the mammalian cell-expressed and glycosylated IFNβ-1a of Avonex is significantly less immunogenic than the bacterial cell expressed and nonglycosylated IFNβ-1b of Betaferon when administered as per their respective approved regimens. Of the untreated, thermally stressed, and copper-oxidized rhIFNβ-1b tests articles, the immunogenicity in transgenic mice in terms of rank order from highest to lowest was thermally stressed > untreated > copper oxidized, and in terms of all 5 test articles, the rank order was Betaferon drug product > thermally stressed rhIFNβ-1b > untreated rhIFNβ-1b > copper-oxidized rhIFNβ-1b > Avonex drug substance (Fig. 9), with all but the Avonex drug substance-treated transgenic mice forming anti-IFNβ antibodies in 100% of the animals after 3 weeks. No antibodies were detected in Avonex drug substance or copper-oxidized rhIFNβ-1b-treated transgenic mice at day 7 (after 5 injections), compared to 10%, 30%, and 30% of the transgenic mice administered with untreated rhIFNβ-1b, thermally stressed rhIFNβ-1b, and Betaferon drug product, respectively. Previous studies of Avonex and Betaferon also showed that antibody formation in IFNβ-transgenic mice occurred more rapidly following administration of Betaferon (Hermeling and others 2005b, 2006; van Beers and others 2010c).
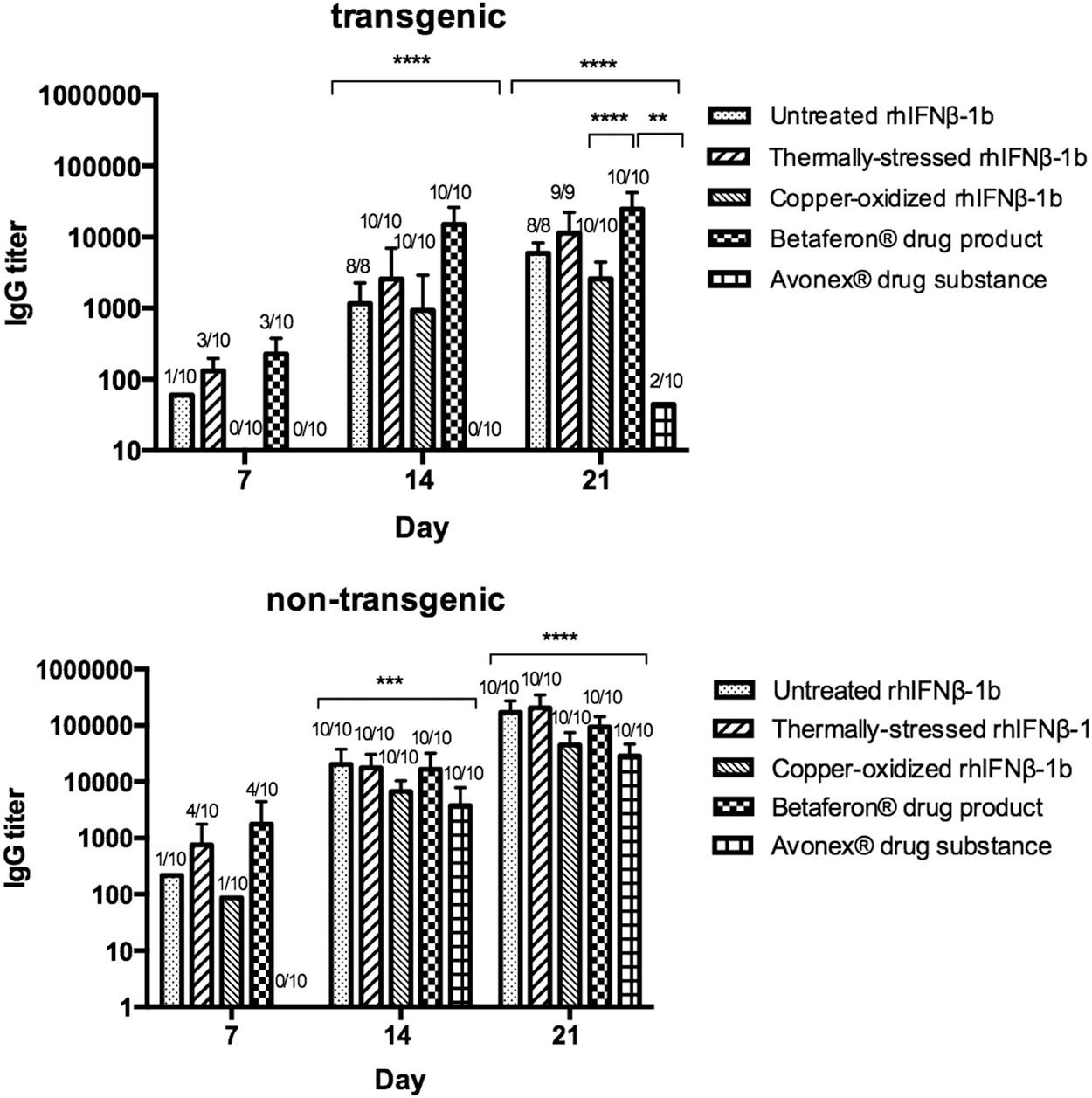
Anti-IFNβ antibody formation in transgenic and nontransgenic FVB/N mice treated with 5 μg of untreated, thermally stressed, and copper-oxidized rhIFNβ-1b, Avonex® drug substance, and Betaferon® drug product (5 times/week) for 3 weeks. Bars represent the average titer of immunoglobulin G (IgG)-positive mice with the corresponding standard error of the means. Statistical analyses (nonparametric Kruskal–Wallis one-way analysis of variance) were performed and showed significant difference in antibody titers among groups in both transgenic and nontransgenic mice on days 14 and 21. In all treatment groups, IgG titers were higher in nontransgenic than transgenic. Furthermore, Dunn's multiple comparisons test showed transgenic mice treated with Betaferon drug product for 21 days significantly enhanced in IgG titers compared to the copper-oxidized rhIFNβ-1b (****P < 0.0001) and Avonex drug substance (**P < 0.01). (*P < 0.05; **P < 0.01; ***P < 0.001; ****P < 0.0001).
Discussion
Biophysical characteristics such as aggregates in protein pharmaceuticals are known to be a significant factor in unwanted immunogenicity (Schernthaner 1993; Braun and others 1997; Hochuli 1997). To discover the relationship between biophysical characteristics and immunogenicity, the immunogenicity of 5 rhIFNβ test articles were investigated in transgenic human interferon beta immune-tolerant mice.
Among 5 rhIFNβ test articles, Avonex drug substance barely broke immune tolerance, likely due to its biophysical properties such as glycosylation that protected the protein from aggregation and enhanced its stability (Kagawa and others 1988; Runkel and others 1998; Sola and Griebenow 2009). The majority of untreated rhIFNβ-1b contained monomers recovered by UV spectroscopy, SDS-PAGE, and SE-UPLC. However, low amounts of larger noncovalent polydisperse particles were present, as measured by DLS, RMM, and MFI. RhIFNβ-1b monomers present in the untreated sample were in native form as determined by RP-HLPC. We hypothesize that the higher immunogenicity of untreated rhIFNβ-1b compared to the copper-oxidized rhIFNβ-1b is because of more epitopes exposed to the immune system, which was also discussed in previous studies on interferon alpha2b (Hermeling and others 2005a). Untreated rhIFNβ-1b contained mostly native monomeric and slightly large noncovalent native-like aggregates, which were enough to break the tolerance of the transgenic mice.
The copper-oxidized rhIFNβ-1b consisted of oxidized, heterogeneous, visible, and subvisible particles, according to RMM and MFI, with and without covalent bounds observed in SDS-PAGE and Western blot. Large aggregates with a relatively high degree of covalent bonds were detected by DLS in the presence of 0.1% SDS. The UV spectrum of copper-oxidized sample showed an intense light scattering owing to the presence of large number of aggregates in these samples, which could not enter the SE-UPLC column.
The poor immune response of transgenic mice for copper-oxidized sample was explained by the size of aggregates. It might be due to lower exposure of the number of epitopes with the increasing size of aggregates (Hermeling and others 2005a). Furthermore, large aggregates may slow down the clearance from the peritoneal cavity or impair the uptake by phagocytic cells (Hermeling and others 2005a).
Thermally stressed rhIFNβ-1b exhibited smaller and more polydisperse aggregates in the presence and absence of SDS than those in copper oxidized, according to DLS. The UV spectroscopy confirmed the presence of less aggregates in this product compared to the copper-oxidized sample. The dimers were observed by SE-UPLC and larger particles were cleared during filtration step or with precolumn before elution into the column. The heated sample exhibited alterations in the protein conformations, which can be explained by the introduction of new epitopes and changed interactions of protein with RP-HPLC column materials. Fluorescence spectra of both aggregated samples showed a change in tertiary structure of the protein, which was intact in untreated rhIFNβ-1b.
In presence of SDS, the UV spectra of both aggregated rhIFNβ-1b presented significant differences, suggesting not only that high proportions of noncovalent aggregates dissociated with SDS but also a high degree of covalent aggregates present in these samples. Protein oxidation was detected only in copper-oxidized rhIFNβ-1b. Hypothetically, it is possible that the unrecovered fraction (40%) of thermally stressed sample in RP-HPLC included oxidized rhIFNβ-1b. Alteration in tertiary structure of the protein detected for thermally stressed rhIFNβ-1b may lead to the introduction of new epitopes to the immune system and presentation of these may break the immune tolerance (Hermeling and others 2005b).
Betaferon drug product contained dimers and apparently multimeric aggregates (van Beers and others 2011; Barnard and others 2012) similar to thermally stressed rhIFNβ-1b. However, the lower antibody response observed in thermally stressed rhIFNβ-1b compared to Betaferon drug product was probably explained by the change of protein structure and native epitopes. The more native epitopes present, the higher antibody response is observed.
Processes such as oxidation, aggregation, covalent versus noncovalent bonds, degree of conformational change, and introduction of new epitopes are involved in unwanted immune response. The role of these biophysical properties in immunogenicity was confirmed before (van Beers and others 2010a, 2011). Thus, it is difficult to determine the contribution of each to the observed immunogenicity. We believe that a combination of all factors mentioned above could be responsible for the immunogenicity against rhIFNβ-1b.
Together, our data indicate that the dogma of “aggregation leads to immunogenicity” should be taken with caution. Two different aggregated rhIFNβs obtained by copper oxidation and heating, both differ in protein conformation and size distribution of aggregates, and in addition vary substantially in immunogenicity. Whereas, aggregates in thermally stressed rhIFNβ and Betaferon drug product are able to induce high immune responses, those obtained by copper oxidation failed to do so. Taking into account that this study is limited to the transgenic mice, which were genotypically positive for hIFNβ gene and were not confirmed to be phenotypically transgenic, we could not extrapolate these results to humans, and complementary studies in patients are needed.
Overall, among different biophysical characteristics, aggregation plays a significant role in immunogenicity of rhIFNβ-tested articles. Aggregates composed of native epitopes are more likely to elicit immune system responses than aggregates consisting of denatured proteins (Hermeling and others 2005a; van Beers and others 2011). By preventing formation of native-like aggregates, nonimmunogenic therapeutic protein formulations might be achieved. More studies are needed to elucidate the relationship between biophysical characteristics and immunogenicity of rhIFNβ-1b products, to assess the minimum amount of altered protein evoking antibody formation in patients.
Conclusions
This work shows that not only the size of the aggregates but also the shape and structure of the constituent proteins determine immunogenicity. Different rhIFNβ-1b test articles with varied biophysical characteristics showed unlike immune responses in FVB/N-tolerant mice. The copper-oxidized rhIFNβ-1b, in spite of being highly aggregated, showed a low immune response due to the lower number of epitopes exposed to the immune system. However, thermally stressed rhIFNβ-1b, which may lead to the formation of aggregates with distinct characteristics, is capable of inducing a high level of antibodies compared to untreated rhIFNβ-1b as demonstrated in transgenic FVB/N immune-tolerant mice. Betaferon drug product was determined to be the most immunogenic article and Avonex drug substance as the lowest immunogenic as reported previously (van Beers and others 2011; Barnard and others 2012).
Footnotes
Acknowledgments
This research was supported financially by the Ministry of Science, Research and Technology of Iran. Biogen Idec, Inc. and Zistdaru Danesh Co. Ltd. are acknowledged for kindly providing Avonex® drug substance and rhIFNβ-1b drug substance, respectively. We thank Reza Nejadnik (Faculty of Science, Leiden Academic Centre for Drug Research, Leiden University) for his assistance in performing the MFI. Professor Wim Jiskoot from Leiden University is acknowledged for his support and Darren Baker from Biogen Idec for critically reviewing this article. We would like to express our gratitude to John Carpenter for allowing us to perform the Archimedes test in his laboratory.
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.