
Abstract
This study aimed to evaluate the effects of Peste des petits ruminants (PPR) vaccine on cytokine and antibody levels in sheep when administered alone or in combination with Corynebacterium cutis lysate (CCL). The PPR vaccine group received a single subcutaneous axillary injection of the PPR vaccine (1 mL containing tissue culture infectious dose (TCID)50 attenuated live PPRV, n = 6) and the combination treatment (1 mL CCL and 1 mL PPR vaccine, n = 6) groups received a single subcutaneous axillary injection of both CCL and PPR vaccine. Blood samples were collected from sheep before the treatment and at different points after treatment (1, 3, 7, 14, 21, and 28 days). Plasma and serum samples were evaluated for antibody percentage, levels of cytokines IL-6, IL-10, IFN-γ, IL-4, IL-12, and IL-18, oxidative stress marker Thiobarbituric acid reactive substances, and hematological and biochemical parameters. Maximum protective antibody levels reach 3–4 weeks after vaccine administration. The combination treatment resulted in significant changes in IFN-γ, IL-4, IL-12, and IL-18 cytokine levels. These changes were not evident when only the PPR vaccine was administered and antibody percentage against PPRV was short term in PPR vaccine group. In conclusion, combined usage of the PPR vaccine with CCL resulted in a heightened cytokine response, leading to improved antibody level against PPR virus. Repeated CCL treatments can lead to earlier vaccine potency, provide protective efficacy for a longer time, and increase passive immunity.
Introduction
P
Several vaccines are known to generate an immune response through TLR signaling, leading to activation of dendritic cells and secretion of pro-inflammatory cytokines (van Duin and others 2006). PPRV contains hemagglutinin and fusion protein that lead to formation of monoclonal antibodies in the cell after vaccinations (Diallo and others 2007).
PPR vaccine is usually administered once a year and results in generation of immunity 3–4 weeks after vaccination (Altug and others 2013). Development of an effective vaccine with a high immunologic potential and prolonged duration of action requires extensive studies and drug trials (Mohamed and others 2013). Since the PPR vaccine is thermolabile, it is particularly important to develop suitable vaccination strategies to be used in subtropical countries such as Ethiopia (Silva and others 2014). Attenuated morbillivirus vaccines have been shown to induce cell-mediated immunity. However, it is unclear whether this protection is effected by systemic neutralizing antibodies, cytotoxic T lymphocytes, or mucosal immunity (Kumar and others 2014).
Corynebacterium cutis lysate (CCL) is a potent immunostimulant and has been reported to potentiate the immune response to a vaccine when administered in combination (Mohamed and others 2013; Anonymous 2015). Immunostimulants, such as CCL, are captured and digested by macrophages and stimulate the synthesis of cytokines that in turn activate lymphocytes, leading to an inflammatory response and production of antibodies (Yılmaz and Kasıkcı 2013).
Cytokines, glycoproteins released by immune cells, play a major role in the regulation of immunity, inflammation, and cell development and differentiation (Sasi and others 2014). In addition, cytokines play a central role in immunity as Th-1/type-1 cytokines cause predominantly cell-mediated immunity, whereas Th-2/type-2 cytokines cause predominantly humoral immunity (Patel and others 2012). Cell-mediated immunity is leading for intracellular pathogen (O'Garra 1998). IL-12 and IFN-γ induce T helper 1 (Th1) response and IL-4 and IL-10 induce Th2 response. While IL-4 and IL-10 cytokines inhibit macrophage and Th1 cells, IFN plays an important role in primary immune response (O'Garra 1998; Dittmer and others 2001) and Th1 cells enable the eradication of infectious pathogen. After vaccination, level of cytokines switches according to induced cell types and immunity (Patel and others 2012).
Increased expression of IFN-β, IFN-γ, IL-4, IL-1β, IL-8, IL-10, IL-6, and IL-12 has been shown in response to PPR infection (Baron and others 2014). T cells and other cells of the immune system are responsible for secretion of various cytokines and interferons. Th1 cells secrete IFN-γ and IL-2, while Th2 cells secrete IL-4, IL-5, and IL-6 cytokines. IL-12 is released by macrophages and dendritic cells (Choi and Reiser 1998). IFNs released by T and natural killer (NK) cells play a role in antiviral immune response using Janus kinase and signal transducer and activator of transcription (STAT) pathway (Kumar and others 2014). The initial phase of protective effects against PPRV is characterized by a variable expression of IFN-γ and IL-4, while a Th2 response is elicited in the later stage of infection (Patel and others 2012). In addition, increased expression of IL-12 further contributes to high IFN-γ levels (Baron and others 2014), and this increase has been reported to induce differentiation of Th1 cells (Trinchieri 1994). IL-4 plays an important role in the humoral and cell-mediated immune response by inducing B-cell proliferation and T-cell differentiation and survival. IL-6 is a pro-inflammatory cytokine that supports maturation and differentiation of B cells, activation of macrophages, and production of acute phase proteins and antibody.
IL-12 stimulates cytotoxic T lymphocytes and plays a specific role in defense against viruses. In addition, it exhibits synergistic effects with IFN, IL-4, and IL-6 in early cellular immune response (Trinchieri 1994). IL-18, in synergism with IL-12, promotes the release of pro-inflammatory cytokines and production of IFN-γ by NK and T cells (Gracie and others 2003; Bastos and others 2007; Darwich and others 2009). Increased IL-12 and IL-18 production leads to cytotoxic activity and proliferative effects on NK and T cells resulting in protection against infections (Sahoo and others 2011). IL-10 regulates the proliferation and differentiation of Th1, Th2, and antigen-presenting cells. IL-10 levels have been shown to increase in chronic viral infections and play a role in viral clearance (Filippi and von Herrath 2008; Baron and others 2014).
Since CCL has been shown to act as a potent immunostimulant resulting in improved immune responses when administered with vaccines, we speculate whether it increases vaccine potency when administered in combination with PPR vaccine. The aim of this study was to determine the effect of PPR vaccine alone and in combination with CCL on antibody and cytokine levels in sheep.
Materials and Methods
Animals and experimental design
Twelve Akkaraman ewes (unvaccinated for PPR, aged ≥6 months, and weighing 40–50 kg) were used in this study. Ewes received yearling fattening feed, barley, vetch hay, sugar beet pulp, and water ad libitum. The study protocol was approved by the Ethics Committee (Veterinary Faculty, Selcuk University, Konya, Turkey). The ewes were divided into 2 groups of 6 sheep each. First group (PPR vaccine group) was given a single subcutaneous axillary injection of 1 mL of vaccine consisting of 103 tissue culture infectious dose (TCID)50 attenuated live PPRV (Pestdoll-S®, lyophilized vaccine strain PPR75/1; Dollvet). Second group (combination treatment group) received 20 mg/mL of the commercial product containing CCL (Ultra-corn® Inj. Susp. Virbac Animal Health) along with a single subcutaneous dose of PPR vaccine similar to the PPR vaccine group.
Analysis of parameters
Blood was drawn before administration (0 h, control) and after vaccination (1, 3, 7, 14, 21, and 28 days) by catheter from vena jugularis into serum gel and anticoagulant (EDTA) plastic tubes. Hematological parameters (including red blood cell, white blood cell, and platelet counts, hematocrit and hemoglobin) were determined by hemocell counter (BC-2800 Auto Hematology Analyzer; Mindray Bio-Medical Electronics).
The serum biochemical values (creatine kinase-MB, alkaline phosphatase, alanine aminotransferase, aspartate aminotransferase, gamma glutamyl transferase, total protein, albumin, blood urea nitrogen, and creatinine) were measured by autoanalyzer (Cobas Integra® 400 plus; Roche Diagnostics).
All ELISA Kits were prepared according to the prescribing in ELISA prospectus. Serum thiobarbituric acid reactive substance (TBARS Assay Kit, Item no: 10009055; Cayman Chemical Company) level for oxidative stress, PPR antibody levels (ID Screen® PPR Competition ELISA Kit, Catalogue No. PPRC-4P; ID-Vet) for immune response, and IFN-γ (Sheep IFN-γ ELISA Kit, Catalogue No. 201-07-0063; Shanghai Sunred Biological Technology Co., Ltd.), IL-6 (Sheep IL-6 ELISA Kit, Catalogue No. 201-07-0009; Shanghai Sunred Biological Technology Co., Ltd.), IL-10 (IL-10 ELISA Kit Sheep, Catalogue No. 201-07-0068; Shanghai Sunred Biological Technology Co., Ltd.), IL-12 (IL-12 ELISA Kit Sheep, Catalogue No. 201-07-0081; Sunred Shanghai Biological Technology Co., Ltd.), IL-18 (Sheep IL-18 ELISA Kit, Catalogue No. 201-07-0061; Shanghai Sunred Biological Technology Co., Ltd.), IL-4 (IL-4 ELISA Kit Sheep, Catalogue No. 201-07-1006; Sunred Shanghai Biological Technology Co., Ltd.) cytokine levels were determined by ELISA reader (MWGt Lambda Scan 200; Bio-Tek Instruments).
Antibody percentage for PPRV is interpreted and above 65% values are positive according to the manufacturer's protocol.
Statistical analysis
All values are depicted as mean ± standard error of mean. The results were analyzed using analysis of variance and Duncan test as a post hoc test (SPSS 19.0). In all cases, P < 0.05 was the criterion for statistical significance.
Results
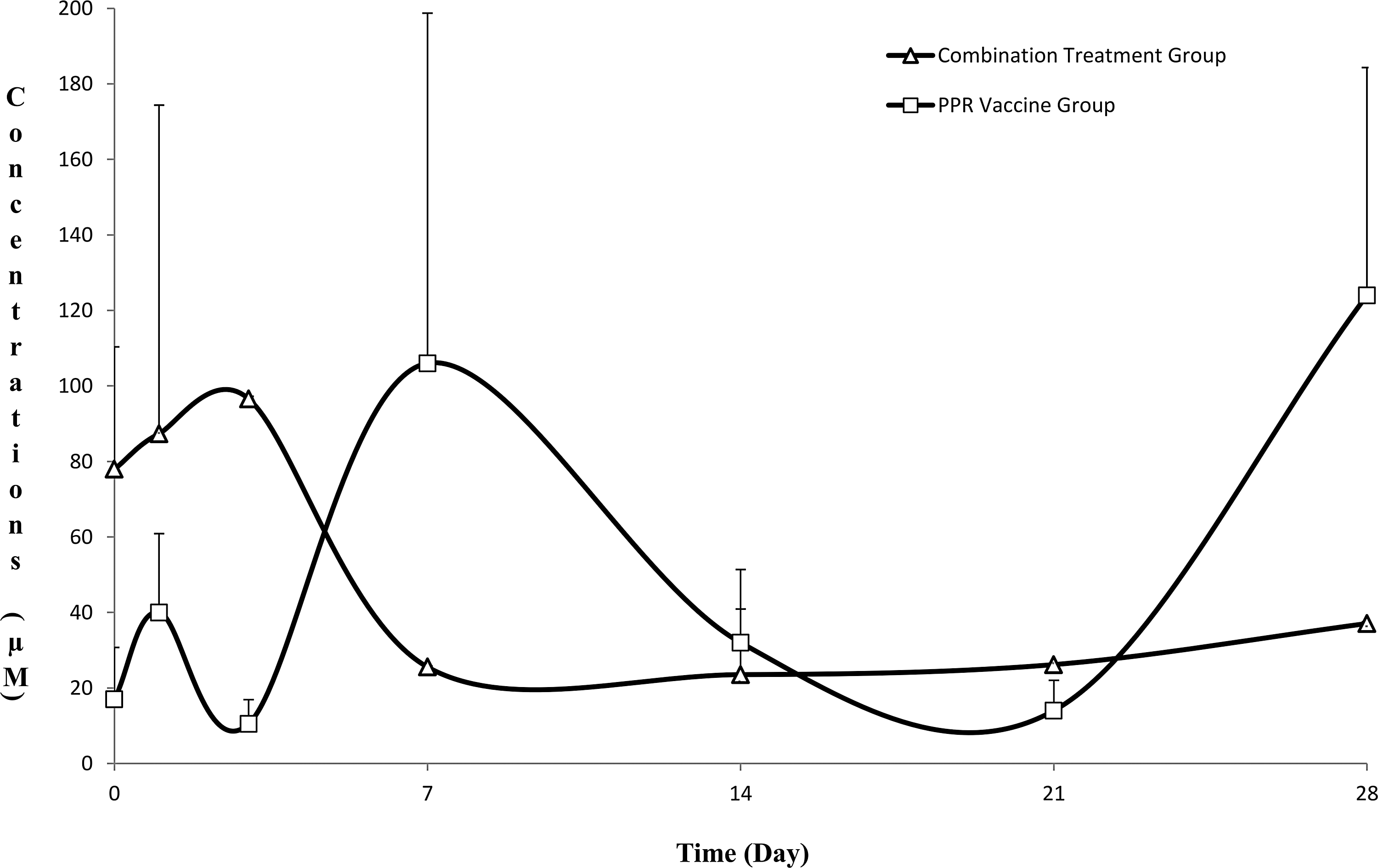
The antibody percentage in both groups (PPR vaccine group and combination treatment group) is depicted in Fig. 1. PPR-specific antibody percentage test valid according to the manufacturer protocol and different antibody levels were revealed among the days. Antibody percentage against PPRV had more high level and longer time in the combination group compared to the PPR vaccine group.

The antibody percentages in Peste des petits ruminants (PPR) vaccine and combination treatment groups [mean ± standard error of mean (SEM)]. Dashed Line: If the percentage of inhibition was ≥65, the group was said to have specific antibody against PPR virus.
The changes in serum IFN-γ, IL-4, IL-6, IL-10, IL-12, and IL-18 levels in PPR vaccine and combination treatment groups are presented in Tables 1 and 2. Day 21 in the PPR vaccine and day 7 in the combination group had maximum IFN-γ level. IL-4 levels were highest on day 28 and day 7 in PPR vaccine group and the combination treatment group, respectively. The IL-6, IL-10, and IL-18 were at high level on day 28 in PPR vaccine group and on day 21in the combination group. Peak levels of IL-12 were on day 28 in the PPR vaccine group and on day 3 in the combination group. The changes in the cytokine levels were significant for effective time and force in the combination groups (Tables 1 and 2).
Values are given in mean ± SEM.
Different letters in the same line (a,b,c) denote statistical significance (P < 0.05).
SEM, standard error of mean; TCID, tissue culture infectious dose.
Values are given in mean ± SEM.
Different letters in the same line (a,b,c) denote statistical significance (P < 0.05).
TBARS levels did not show statistically significant changes in both PPR vaccine and combination treatment groups (Fig. 2).
TBARS concentrations in PPR vaccine and combination treatment groups (mean ± SEM).
Hematological parameters for PPR vaccine and combination treatment groups are shown in Tables 3 and 4, while biochemistry parameters are shown in Tables 5 and 6, respectively. Both hematological and biochemical parameters were found to be statistically different in both groups (P < 0.05). All of these parameters were in reference limits in both groups.
Values are given in mean ± SEM.
Different letters in the same line (a,b,c) denote statistical significance (P < 0.05).
GRAN, granulocyte; HCT, hematocrit; HGB, hemoglobin; LYMPH, lymphocyte; MON, monocyte; PLT, platelet; RBC, red blood cell; WBC, white blood cell.
Values are given in mean ± SEM.
Different letters in the same line (a,b,c) denote statistical significance (P < 0.05).
Values are given in mean ± SEM.
Different letters in the same line (a,b,c) denote statistical significance (P < 0.05).
ALB, albumin; ALP, alkaline phosphatase; ALT, alanine aminotransferase; AST, aspartate aminotransferase; BUN, blood urea nitrogen; CK-MB, creatine kinase MB; CREAT, creatinine; GGT, gamma glutamyl transferase; TPROT, total protein.
Values are given in mean ± SEM.
Different letters in the same line (a,b,c) denote statistical significance (P < 0.05).
Discussion
PPR is a highly infectious disease affecting sheep and goats and resulting in high rates of mortality and morbidity. Considering the great economic losses caused by death of these animals, there is an urgent need to control this disease and develop suitable strategies for its eradication. However, full protection and high antibody level cannot ensure against the disease due to lack of vaccine efficiency, nonthermostable vaccine, poor immunity of the herd, and other reasons (Abubakar and others 2011; Silva and others 2014).
Systemic neutralizing antibody titers are reported to increase from 7 to 21 days after PPR vaccination (Hosamani and others 2006; Patel and others 2012). Studies have shown that single dose of PPR vaccine is able to provide protective immunity 14 days postvaccination and it is maintained throughout the year. However, the antibody inhibition titers were variable (decrease after 30 days, increase after 90 and 150 days) (Singh and others 2004; Rajak and others 2005). In this case, additional medical treatment is needed for maintenance of stable antibody titers. Another study has shown that total leukocyte and lymphocyte levels varied in parallel with the antibody levels in goats postvaccination (Rajak and others 2005). A combination treatment of CCL with some vaccines is reported to increase protective antibody titers and high IgG levels in pregnant sheep (Mohamed and others 2013). In this study, we aimed at investigating the effect of PPR vaccine alone and PPR vaccine in combination with CCL on the potential protective immunity against PPR.
In the current study, the peak antibody level is observed on day 7 postvaccination followed by a decrease after 21 days in the PPR vaccine group. Total leukocyte and lymphocyte numbers underwent statistically significant reduction in both PPR vaccine and combination treatment groups depending on live vaccine. These variations were within the normal reference range (Bulbul 2013). In the combination treatment group, peak antibody percentage was observed after 7 days, while the protective immune response continued throughout the course of the experiment. PPR vaccine with oil-based adjuvant and at a dose higher than 103 TCID50 is reported to generate a more favorable immune response (Anees and others 2014). Immune response to viral diseases, such as PPR, has been shown to stimulate the production of IFN-γ and IL-4 (Patel and others 2012). In the present study, antibody levels were not maintained for a long time in the PPR vaccine group and the vaccine led to generation of short-term antibody titers. The combination treatment could induce Th1 cells in the early phase and lead to long-term stimulation of Th2 cells. Longer time expressions of IFN and other cytokines may be depending on these effects. Therefore, the PPR vaccine in combination with CCL treatment may provide higher antibody percentage in a shorter time and contribute to the vaccination program. CCL may increase due to force of vaccine.
IL-12 and IL-18 induce production of IFN-γ that is important for an antiviral immune response. IFN-γ modulates Th1 and Th2 cells and plays a role in cytotoxic T-cell response (Shtrichman and Samuel 2001). In addition, IFNs are known to exhibit antiviral response through the STAT pathway (Durbin and others 2002). In the current study, we show that a combination treatment with CCL may lead to increased IFN levels, activation of immune system cells, and signaling through the STAT pathway in the early-phase postvaccination. IL-12 and IL-18 cytokines induce Th1 cells and result in increased IFN-γ synthesis using p38 mitogen-activated protein kinase (MAPK) pathway leading to a more long-lived T cell and inflammatory response (Yu and others 2003). In the present study, IL-12 and IL-18 levels did not change significantly. However, increase in IFN-γ levels was more apparent in the combination treatment group compared to the PPR vaccine group. IL-12 and IL-18 cytokines have been reported to synthesize in correlation and provide macrophage activation (Bastos and others 2007). Similar results were obtained for these cytokines in the present study and IFN-γ level earlier increased in combination treatment group compared to PPR vaccine group. This increase may be because of the stimulatory effect of CCL such as IL-12 and IL-18 on macrophages (Yılmaz and Kasıkcı 2013).
IL-10 increased extremely in the PPR vaccine group, but it was variable in combination treatment group and decreased after 21 days. IL-10 is expressed mainly on monocytes and T and B lymphocytes, and its increased levels are associated with a decrease in viral clearance (Brockman and others 2009). Experimental PPR infections in goats have shown an increase in IL-10 levels after day 4 postinfection. An increase in IL-10 and IL-12 results in increased IFN secretion by NK cells (Baron and others 2014). The enhancement of IL-10 level may be from derived positive feedback mechanism in PPR vaccine and combination groups. In addition, an early increase in IL-10 levels can be in response to live virus and CCL in the combination group. These can explain an increase in numbers of immune cells, such as monocytes, depending on CCL treatment. In addition, CCL may be increased by IFN secretion linked to IL-10 level.
IL-6 and IL-10 levels have been reported to increase some viral infections (Ergonul and others 2006; De Pablo-Bernal and others 2016). IL-6 induces IL-10 expression and an increased expression of these cytokines is observed as long as the virus remains alive (Yu and others 2011). Attenuated virus can lead to increase in IL-10 levels as positive feedback because of chronic stimulation of immune system cells, rising inflammation, and pro-inflammatory cytokines in the present study.
IL-4 level fluctuated in combination treatment group although it increased duration of the experiment in the PPR vaccine group. IL-4 and IL-10 cytokines are known to shift the immune response to the Th2 type. Reduced expressions of these cytokines are important for the healing process. As a result, the release of these cytokines by Th2 cells should be regulated (Connors and others 1994; Johnson and Graham 1999). In the present study, we observed a significant increase in IL-10 levels, whereas IL-4 levels remained unchanged in the PPR vaccine group. In the combination treatment group, IL-4 and IL-10 levels varied during the course of experiment, probably because of modulation by CCL. These findings suggest that delayed type immune response can be prevented by CCL. In addition, combination therapy generated the desired immune response with CCL regulating the antibody levels and production of IFNs and other cytokines from Th1 cells. IL-4 is reported to inhibit the activity of memory T cells in mice (Jackson and others 2001). Therefore, suppression of IL-4 secretion is desired in case of viral infections and after vaccination (Dittmer and others 2001). CCL cannot block memory T cells because IL-4 level causes overall fluctuations. So, it can enhance immunity because of rise of antibody level markedly.
Oxidative stress plays an important role in viral diseases, with an increased oxidative stress resulting in cell damage (Hahn and others 2008). In vitro studies have shown that a decrease in oxidative stress and cell damage caused to reduce viral replication after some treatment (Chen and others 2015). In a previous study, CCL has been shown to have no effect on the levels of TBARS in healthy sheep (Er and others 2014). Its levels are expected to increase with time (between 7 and 28 days) depending on the live attenuated virus vaccine in the PPR vaccine group. However, oxidative stress and cell damage are expected to be regulated in the combination treatment group.
Oxidative stress increases and the antioxidant capacity (SOD, GPX, and CAT levels) decreases depending on virus replication (Yavru and others 2015). Live PPR vaccine resulted in increased oxidative stress and inflammation in response to virus replication; however, TBARS levels did not show a significant change in the present study. In addition, oxidation and DNA damage can be reduced as this increase is minimized by CCL.
Some bacterial, parasitic, and viral infections can result with changes in biochemical and hematological parameters (Kerr 2008). We observed a decrease in ALP levels after day 14 postvaccination and an increase in total protein level at all sampling points compared to control in the PPR vaccine group. However, all biochemical parameters were within the reference range in both the groups (Turgut 2000; Kerr 2008) and no significant changes were observed in sheep (Avcı and others 2013). Thus, both PPR vaccine alone and combination treatment with CCL did not result in organ or metabolic damage in the current study.
Hematological parameters were observed to fluctuate within the reference range in both groups (Turgut 2000; Kerr 2008). Increased RBC and platelet numbers may be associated with the increase in IL-6 in PPR vaccine group as heightened IL-6 level is reported to increase RBC and thrombocyte cells (Tanaka and others 2014). Although the observed changes in other hematological parameters were reference range, they need to be further investigated through detailed studies in future.
CCL administration in the combination treatment group did not cause any side effects or changes in hematological and biochemical parameters that have been observed on administration of some drugs or adjuvants.
Thus, we show that CCL administration with PPR vaccine can induce significant alterations in cytokines and it may improve effects of the PPR vaccine. Combination treatment with CCL can lead to an early and heightened cytokine response leading to better vaccine efficacy. In addition, higher doses of CCL with vaccine treatment can ensure earlier time and higher antibody level against diseases. Administration of CCL along with PPR vaccine may be a feasible approach to control PPRV infection and reduce the economic loss caused by PPR. In addition, the combination treatment can significantly reduce the cost of vaccination and reduce stress to the animal. Further studies are required to ascertain the efficacy of combination treatment with CCL in goats that are also targeted by PPRV and further challenge studies are needed to determine the protection.
Footnotes
Acknowledgments
This study is supported by SUPABK (15401153). A part of the abstract will be presented at the 6th European Congress of Virology (ECV), Hamburg, Germany and the other part of the abstract was presented at the 18th International Conference on Laboratory Medicine and Clinical Research, Amsterdam, the Netherlands.
Author Disclosure Statement
No competing financial interests exist.