
Abstract
Hepatitis C virus (HCV) is the leading cause of liver fibrosis and hepatocellular carcinoma (HCC). At present, there is no predictive biomarker for the patients at high risk of developing HCC. In this study, we examined the association between single-nucleotide polymorphisms (SNPs) in 3 innate immunity genes [2′-5′oligoadenylate synthetase 1 (OAS1) rs10774671, interleukin 28B (IL28B) rs12979860, and low molecular mass polypeptide 7 (LMP-7) at codon 49] besides cytomegalovirus (CMV) coinfection and susceptibility to HCC in genotype 4 (GT4) chronically infected Egyptian patients. SNPs were determined using restriction fragment length polymorphism analysis in DNA from HCC patients (n = 34) and compared with either controls (n = 70) or patients with early grades of liver fibrosis (n = 49). Our results demonstrated that patients bearing the genetic combination consisting of LMP-7 CA/AA [OR 4.75, 95% confidence interval (CI) 1.443–15.631, P = 0.007] and IL28B rs12979860 CT/TT (OR 6.00, 95% CI 1.603–22.455, P = 0.004) and positive for CMV viremia (OR 3.11, 95% CI 1.151–8.412, P = 0.02) were more likely to have HCC. However, OAS1 rs10774671 does not seem to contribute to the development of HCC. Binary regression analysis indicated that HCC risk significantly increases with the presence of each unfavorable genotype (LMP-7 CA/AA, IL28B rs12979860 CT/TT), when accompanied by the existence of CMV coinfection (probability of HCC risk is 0.8 for combined factors versus 0.14, 0.07, and 0.07 for individual factor IL28B, LMP-7, and CMV; respectively). These data suggest that the 2 SNPs and the coinfection in concert have potential in predicting the risk of HCC development in patients infected with HCV GT4.
Introduction
H
Several etiologies, including viral, environmental, and genetic factors, interplay to determine the rate of disease progression in HCV infection (Tolmane and others 2012), of those, host genetic factors represent the major determinants for the risk of HCC development. Single-nucleotide polymorphisms (SNPs) are the most common host genetic factors that are highly linked to the clinical outcome of HCV chronic infection (Ma and others 2015). Interferons (IFNs) play a crucial role in combating infectious disease such as HCV, thereby inhibiting the viral proliferation (Koziel 1999). Cumulative data refer to the key role of IFNs in the viral clearance either spontaneous or drug induced (Rauch and others 2010b), as well as in the disease pathogenesis (McCaughan and George 2004; Sebastiani and Alberti 2006; Ishida and others 2011).
Recently, we have shown a strong association between SNPs in 3 genes involved in early immune response against HCV and the risk of progressive liver disease in genotype 4 (GT4) infection; low molecular mass polypeptide 7 (LMP-7) (Omran and others 2013), interleukin 28B (IL28B) (El-Awady and others 2012), and 2′-5′oligoadenylate synthetase 1 (OAS1) (Bader El Din and others 2015). IL28B gene is located on human chromosome 19q and triggers its antiviral effect through driving the JAK-STAT pathway (Par and others 2014). LMP-7 gene resides on chromosome 6 inside the major histocompatibility complex (MHC) class II region and encodes a catalytic subunit of proteosome complex (Trowsdale and others 1990) that helps in the presentation of intracellular peptides by MHC class I (York and Rock 1996). OAS1 gene is an IFN-stimulated gene that, by RNA splicing, encodes various isoforms of OAS enzyme (with varying enzymatic activities). Its main function is the activation of latent ribonuclease leading to viral RNA degradation. We have found that IL28B SNP rs12979860 (located at 3 kb upstream of IL28B), OAS1 SNP rs10774671 (located at exon 7 splice acceptor site 1), and LMP-7 SNP at codon 49 highly influence the clinical course of HCV infection, response to IFN therapy, and the progression of hepatic fibrosis in GT4-infected patients (Ge and others 2009; Tanaka and others 2009; Thomas and others 2009; Rauch and others 2010a; El Awady and others 2011; El-Awady and others 2012; Omran and others 2013; Bader El Din and others 2015).
In synergy with genetic factors, coinfection with other pathogens strongly influences the clinical outcome of HCV infection. Coinfection with HIV or HBV leads to more progressive liver diseases and less response to IFN-based therapy (Park and others 2005). We have shown in our previous studies that the incidence of cytomegalovirus (CMV) coinfection is more frequent among HCV GT4-infected patients showing extremely poor response to IFN therapy (Bader el-Din and others 2011), and treatment-naive patients with higher grades of hepatic fibrosis (unpublished data), presumably due to the inhibition of type I IFN signaling pathway, ie, JAK-STAT cascade.
Identifying noninvasive early biomarkers is very important to predict patients with higher risk of disease progression who are candidates of early therapeutic intervention for the sake of halting progression into liver insufficiency and HCC. In light of our earlier findings, herein, we investigated a possible predictive value of combined LMP-7-, OAS1-, and IL28B-pretested SNPs, besides CMV coinfection for HCC development from early stages of HCV-induced liver fibrosis, ie, F1-F2.
Materials and Methods
Ethical statement
The institution ethics review board (medical research ethics committee at National Research Center, Cairo, Egypt) approved all experiments according to Helsinki Declaration 1975 revised in 2008. Written informed consent was obtained from each subject before blood collection.
Patients and healthy subjects
This study was executed on 153 Egyptian subjects [70 controls and 83 HCV-chronically infected patients with early hepatic fibrosis (F1-F2, n = 49) and with HCC (n = 34)]. All the patients were recruited from the Department of Tropical Medicine, Kasr El Ainy Hospital, Cairo University and Cairo transportation authority hospital in the period between August 2013 and June 2014. None of the selected patients was alcoholic, diabetic, drug addicted, or suffered from other causes leading to chronic liver diseases. All patients had positive HCV antigen and Ab, negative hepatitis B surface antigen (HBVsAg) and Ab, negative antischistosomal Ab, and negative autoimmune markers. The degree of fibrosis was assessed histologically in liver biopsy and calculated with Knodell and Metavir scoring system. Patients with HCC were diagnosed by 2 imaging modalities. Healthy controls did fulfill the criteria of having negative hepatitis markers (HCV Ab, HBVsAg, or HDV Ab) or hepatotropic infections, eg, schistosomiases, autoimmune diseases, and long-term drug-induced hepatitis. All controls had normal liver function parameters.
Genomic DNA extraction
DNA was extracted from peripheral blood using the Qiagen DNA extraction kit (Qiagen), following the manufacturer's instructions. DNA samples were kept at −20°C until ready to use.
Analysis of IL28B rs12979860 polymorphism
IL28B rs12979860 polymorphism was genotyped by polymerase chain reaction-based restriction fragment length polymorphism (PCR-RFLP) as mentioned previously (El-Awady and others 2012). Following the PCR, the amplified products (139 bp) were digested with 1 U of the BstUI restriction endonuclease (New England Biolabs) at 37°C for 4–6 h or overnight. The digested products were electrophoresed on 4% agarose gel. Uncut fragment of 139 bp represents the homozygous TT genotype, whereas the homozygous CC genotype gave rise to a fragment of 109 bp and the heterozygous CT genotype gave rise to 2 fragments of 139 and 109 bp.
Analysis of LMP-7 genotypes
The genotyping of LMP-7 polymorphism was carried out by PCR-RFLP as described by Omran and others (2013). The 304 bp amplified PCR product was digested overnight with 10 U of BsmI restriction endonuclease (Amersham Pharmacia-Biotech) at 37°C. The digested products were electrophoresed in a 2% agarose gel. The homozygous genotype CC gave rise to 2 fragments of 174 and 130 bp, the heterozygous genotype CA gave rise to 3 fragments of 174, 130, and 304 bp, whereas the AA genotype resulted in an uncut fragment of 304 bp.
Analysis of OAS1 genotypes
OAS1 exon 7 SNP rs10774671 was analyzed by PCR-RFLP as shown by El Awady and others (2011). The 203 bp PCR-amplified fragment was digested with 5 U of the restriction enzyme AluI (Promega) at 37°C for 4 h. The digested products were electrophoresed on a 2% agarose gel. The uncut PCR fragment (203 bp) represented the homozygous GG genotype, while the fragments of 150 and 53 bp represented the homozygous AA genotype, and the 3 fragments of 203, 150, and 53 bp represented the heterozygous AG.
CMV DNA detection
Serum CMV viremia was detected by 2 rounds of PCR amplification using sequence-specific primers targeting the gB region of CMV as described previously (Fox and others 1995; Jones and others 2000). The thermal cycling protocol was the same for both first and second PCR rounds (initial denaturation for 5 min at 94°C followed by 35 cycles of 1 min at 94°C, 1 min at 55°C, and 1 min at 72°C; and final extension for 10 min at 72°C). The nested PCR products (100 bp) were electrophoresed on 3% agarose stained with ethidium bromide.
Statistical analysis
The clinical parameters were compared between different groups using the parametric unpaired t-test and were presented as the mean and standard deviation. The distribution of different genotypes and CMV coinfection within groups was analyzed by χ 2 test and variables were described as odds ratio (OR) with 95% confidence interval (CI). The probability of HCC development was measured by binary logistic regression analysis and calculated by the following equation “logit (P) = 2.629269*IL28B + 1.872872*LMP-7 − 1.85132*CMV − 5.23798”; where logit is the natural logarithm of the odds and the coefficient beside each variable of the logistic regression equation is the variable constant. The variable constant reflects the contribution for each variable in calculating the probability of HCC occurrence. The wild type of IL28B rs12979860 and LMP-7 and positive CMV viremia were represented by the digit 1, while the mutant types of IL28B and LMP-7 as well as negative CMV viremia were represented by the digit 2. Difference between groups was considered statistically significant if P ≤ 0.05.
Results
Patients' clinical data
Comparison between early stages of fibrosis (F1-F2) and HCC patients showed significant differences in age, body mass index (BMI) and total bilirubin levels (Table 1, P < 0.0001 for all parameters). However, gender, albumin, hemoglobin (HB), asparate aminotransferase (AST), and alanine aminotransferase (ALT) did not reveal any significant variation between the 2 groups of patients (Table 1). Both F1-F2 and HCC had lower HB and higher AST and ALT values (Table 1, P < 0.0001) when compared with controls with no significant difference in gender distribution. Mean age and BMI were similar in controls and F1-F2, but higher in HCC cases (Table 1, P < 0.0001).
n refers to the sample size. Data are expressed as mean and standard deviation. Bilirubin and albumin were not measured for healthy controls.
P value is calculated based on the comparison between F1-F2 and HCC groups.
ALT, alanine aminotransferase; AST, asparate aminotransferase; BMI, body mass index; HB, hemoglobin; HCC, hepatocellular carcinoma; HCV, hepatitis C virus.
Distribution of LMP-7, OAS1, and IL28B genotypes and CMV coinfection in healthy controls and chronic HCV patients with early hepatic fibrosis and HCC
Our data revealed significant differences in LMP-7 and IL28B variants in F1-F2 and HCC patients (Table 2 and Fig. 1A, B). LMP-7 CC genotype was more frequent in patients with early fibrosis, while AA and CA genotypes were more frequent in HCC patients (Fig. 1A and Table 2, OR 4.75, 95% CI 1.443–15.631, P = 0.007). For IL28B rs12979860, the frequency of CC genotype was significantly higher in early fibrotic patients, whereas the frequency of CT and TT genotypes was significantly higher in HCC patients (Fig. 1B and Table 2, OR 6.00, 95% CI 1.603–22.455, P = 0.004). No significant difference was noticed in the distribution of OAS1 rs10774671 genetic variants (GG, GA, AA) (Fig. 1C and Table 2, P = 0.3). The incidence of CMV coinfection was more in HCC patients than in F1-F2 counterparts (Fig. 1D and Table 2, OR 3.11, 95% CI 1.151–8.412, P = 0.02). When the comparison was set between controls and F1-F2 patients, OAS1 rs10774671 genotypes showed statistically significant differences in distribution between healthy subjects and early fibrotic patients with GG more prevalent in controls and GA/AA more prevalent in F1-F2 patients (Fig. 1C and Table 2, OR 4.706, 95% CI 1.988–11.14, P = 0.0001). However, LMP-7 and IL28B rs12979860 genotypes showed similar distribution between the 2 groups of subjects (Fig. 1A, B and Table 2, P = 0.1 and 0.4 for LMP-7 and IL28B, respectively). Current data showed varying distribution of LMP-7, OAS1 rs10774671, and IL28B rs12979860 genotypes between healthy subjects and HCC patients, where LMP-7 CC, OAS1 rs10774671 GG, and IL28B rs12979860 CC genotypes were more frequent in controls, while LMP-7 CA/AA, OAS1 rs10774671 GA/AA, and IL28B rs12979860 CT/TT genotypes were more frequent in HCC patients (Fig. 1A, C and Table 2, OR 8.906, 95% CI 2.836–27.97, P = 0.0001 for LMP-7; OR 8.214, 95% CI 2.294–29.409, P = 0.0001 for IL28B rs12979860; and OR 6.141, 95% CI 2.131–17.7, P = 0.0001 for OAS1 rs10774671).
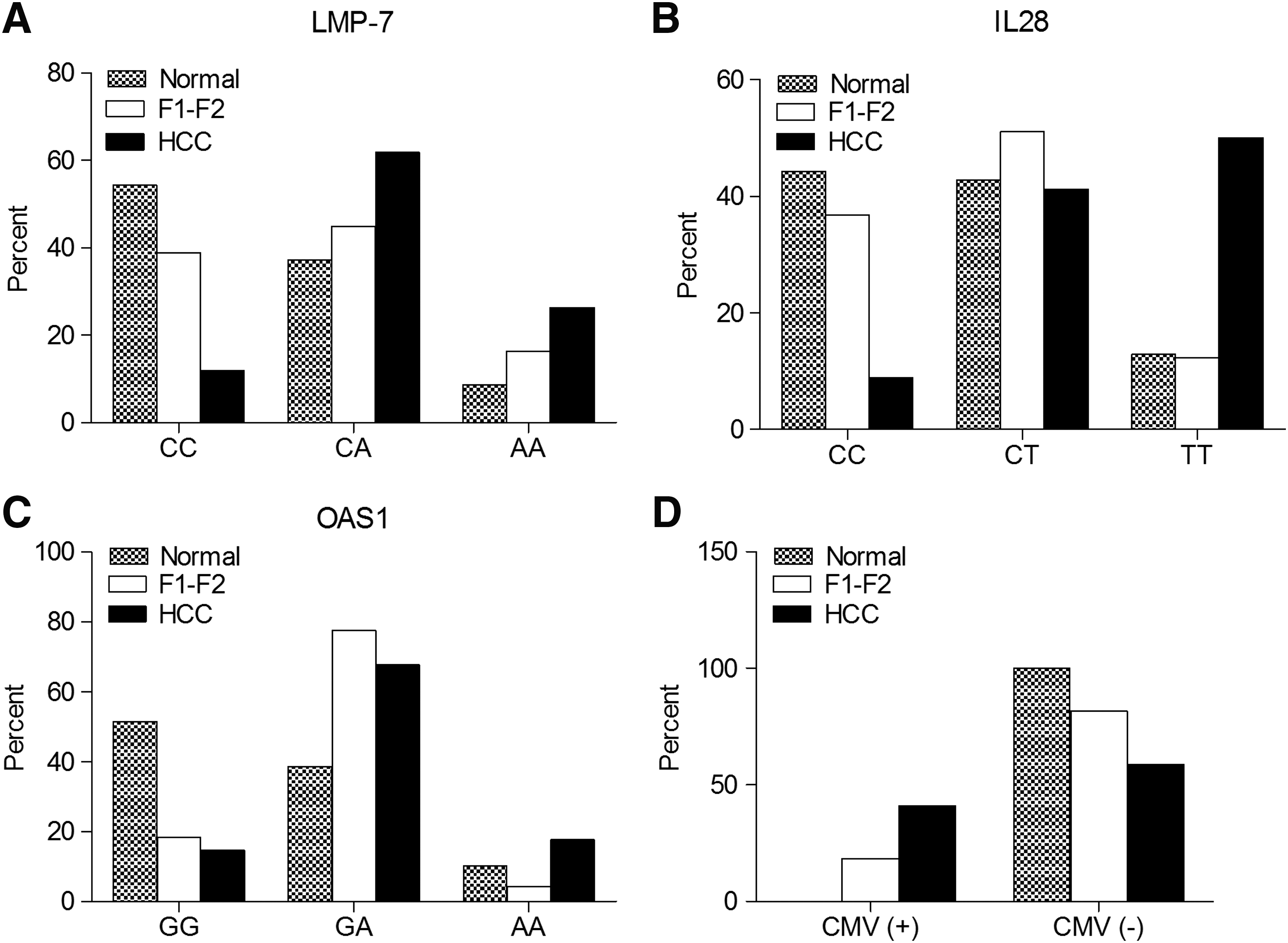
The distribution of LMP-7, IL28B, and OAS1 genotypes and CMV viremia in healthy subjects and HCV patients with early fibrosis (F1-F2) and HCC. The percent of different genotypes of LMP-7 SNP at codon 49
n refers to the sample size.
Genotype chosen as reference category.
Positive CMV chosen as reference category.
CI, confidence interval; CMV, cytomegalovirus; IL28B, interleukin 28B; LMP-7, low molecular mass polypeptide 7; OAS1, 2′-5′oligoadenylate synthetase 1; OR, odds ratio.
Binary logistic regression analysis
Binary logistic regression analysis was used to calculate the probability of HCC development using the logit (P) equation, including all the differentially distributed variables (IL28B, LMP-7, and CMV) (Table 3A). Binary regression analysis indicated that HCC risk significantly increases with the presence of each unfavorable genotype (LMP-7 CA/AA and IL28B rs12979860 CT/TT) when accompanied by the existence of CMV coinfection (probability of HCC risk is 0.8 for combined factors versus 0.14, 0.07, 0.07 for individual factor IL28B, LMP-7, and CMV, respectively, Table 3B). Moreover, our data revealed that 12%, 56%, and 32% of HCC cases had probability less than 0.25, between 0.25-0.5, and above 0.5, respectively (versus 49%, 51%, and 0% of F1-F2 cases, respectively) (Fig. 2).
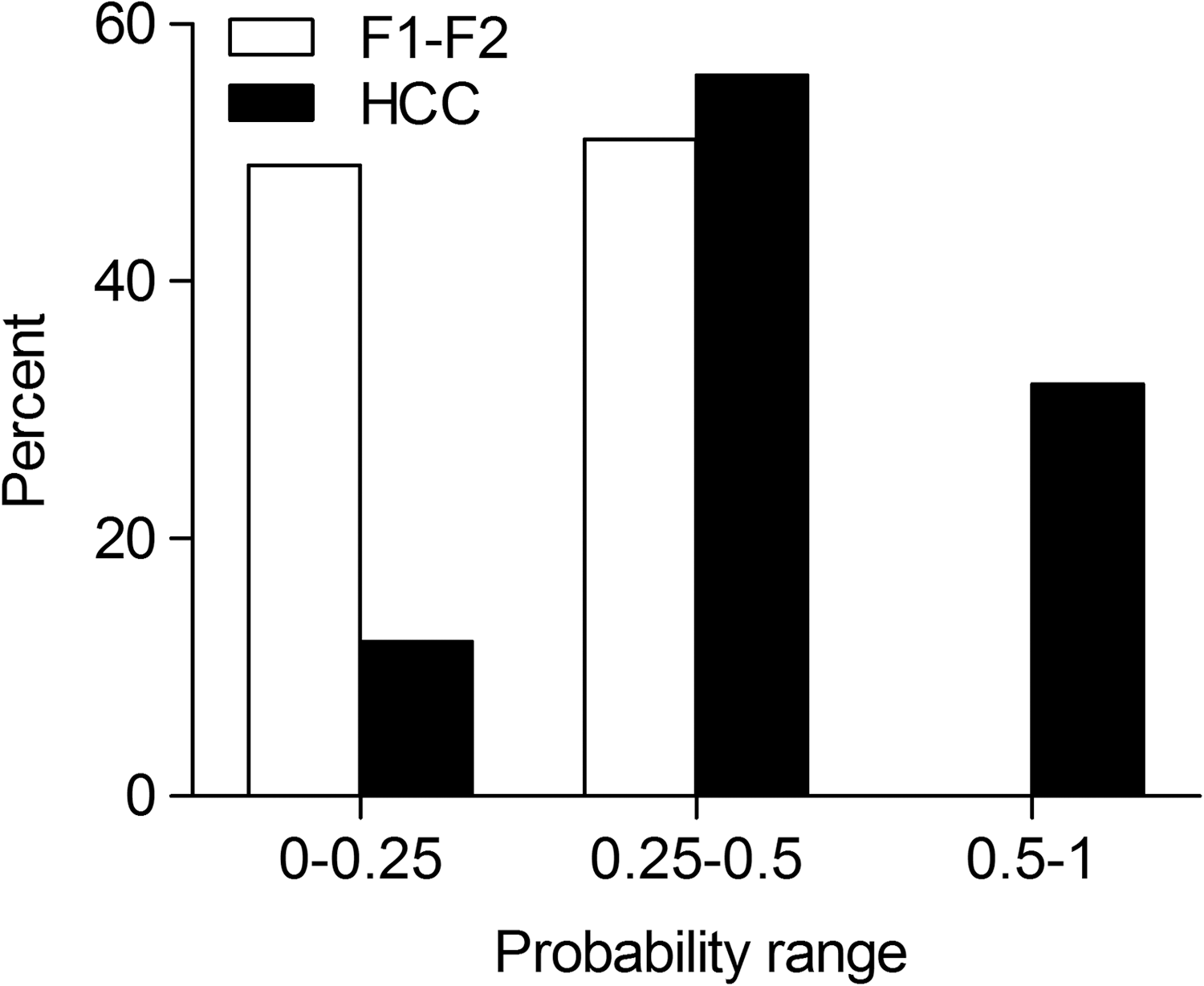
The probability of HCC development in HCV patients with early fibrosis (F1-F2) and HCC. Binary logistic regression analysis was performed to calculate the probability of HCC development, and the percent of patients at different probability ranges (0–0.25, 0.25–0.5, and 0.5–1) was compared between HCV-infected patients with early fibrosis (F1-F2, n = 49) and HCC (n = 34).
logit (P) = 2.629269* IL28B + 1.872872* LMP-7 − 1.85132*CMV −5.23798.
B, the variable constant; Logit, the natural logarithm of the odds.
n refers to the sample size.
F1-F2, early fibrosis.
Discussion
Identifying the immunogenetic factors that increase the susceptibility to HCC in HCV patients improves our understanding of the interplay between HCV and the host immune response leading to increased HCC risk and helps in the discovery of new biomarkers for disease progression. Previously, we showed association between 3 SNPs in key innate immunity genes (LMP-7 at codon 49, OAS1 rs10774671, and IL28B rs12979860) and progressive HCV-related liver fibrosis. Furthermore, the CMV viremia was reported by our laboratory as a promoter in the process of HCV disease progression. However, the association of the combined 4 parameters with liver disease progression has not been explored so far. In this study, we showed that SNPs in LMP-7 and IL28B rs12979860, as well as CMV coinfection, are individually associated with the development of HCC in HCV patients with liver fibrosis. Also, the risk of HCC increases significantly with the coexistence of unfavorable genotypes of LMP-7 and IL28B accompanied by CMV viremia. These data suggest that the latter 2 SNPs plus CMV coinfection act in concert to predict the risk of HCC development.
Although the majority of studies focus on investigating the biology of cirrhosis to develop new diagnostic tools and therapies for HCC, in this study, we focused on studying the genetics of the early stages of hepatic fibrosis and their association with the progression to HCC. Development of HCC might start at the early stages (Gobel and others 2006), but the majority of patients do not have overt symptoms at those stages and start to have apparent symptoms at the incurably cirrhotic stage, and thereby it is of great significance to define genetic biomarkers to predict patients at higher risk of HCC development, to benefit from early intensive therapies, and to minimize HCC incidence.
LMP-7 is a subunit of immunoproteasome that is involved in antigen sampling step of human MHC class I pathway (Vargas-Alarcon and others 2004). Studies in animals have linked the severity of liver diseases to the low proteasome activity (Liu and others 2008) and reduced levels of LMP-7 mRNA and protein (French and others 2011). We and others have reported association between LMP-7 CA or AA genotypes, and high risk of HCV persistence and progression of hepatic fibrosis (Cui and others 2010; Omran and others 2013). Consistent with the aforementioned studies, we noticed higher frequency of CA/AA genotypes in HCC patients than in healthy and early fibrotic subjects; suggesting a protective role for CC genotype against HCC development. In support of this finding, numerous studies showed the implication of LMP-7 genetic variants in the susceptibility to various kinds of human cancers (Cao and others 2005; Fellerhoff and others 2011; Song and others 2014), which is thought to be associated with CA/AA genotypes (Fellerhoff and others 2011).
IL28B modulates the antiviral immune response through its effect on the stimulation of IFN-stimulated genes (de Veer and others 2001; Stark 2007). IL28B rs12979860 is the most prominent SNP located at 3-kb upstream of the IL28B gene that exerts a causal association with altered response to IFN treatment (Ge and others 2009), spontaneous clearance (Thomas and others 2009), and disease progression in HCV-infected subjects (El-Awady and others 2012). However, the relationship between IL28B rs12979860 and the incidence of HCC did not reach consensus. The current data suggest that IL28B SNP rs12979860 seems to augment the overall risk of HCC development owing to CT/TT genotypes, which were more predominant in HCC patients than in healthy and early fibrotic groups. This finding is consistent with the studies of El-Awady and others (2012) and Chang and others (2015) who proposed a protective role of CC genotype against the progress of liver fibrosis to HCC. However, our findings disagreed with those reported by Akkiz and others (2014) and Zekri and others (2014) who concluded that IL28B rs12979860 genetic variation has no impact on the development of HBV or HCV-related HCC.
Several studies have shown that coinfection of HBV or HIV greatly augments the clinical course of HCV disease and accelerates the progression of liver disease to cirrhosis and HCC (Park and others 2005). However, the coexistence of CMV infection with other viral hepatitis and, in particular, its impact on the severity of liver injury are poorly studied. Among the few existing reports, 1 study showed that HBV and HCV patients having CMV infection have more severe necroinflammation and liver fibrosis than those with negative CMV infection (Bayram and others 2009). The incidence of active CMV infection is very high among HCV-infected patients undergoing liver transplantation, on correlation with higher hepatitis activity index and fibrosis stage (Rosen and others 1997; Razonable and others 2002; Chopra and others 2003). Studies from our laboratory showed association between CMV coinfection and both poor response to IFN therapy (Bader el-Din and others 2011) and increased severity of hepatic fibrosis (unpublished data). In this study, we found direct relationship between CMV coinfection and HCC development in GT4-infected patients, and to our knowledge, this is the first report that links CMV-HCV coinfection to HCC development.
We and others have found the association between SNP at OAS1 rs10774671 and disease progression to end-stage fibrosis (Bader El Din and others 2015) and response to IFN therapy (El Awady and others 2011; Imran and others 2014). In light of this finding, we postulated that genetic variants at OAS1 rs10774671 might play a similar role in HCC development. Reciprocally, these data did not reveal any significant difference in OAS1 rs10774671 genotype distribution in HCC patients compared to those with early fibrosis. On the other hand, the protective GG genotype was found significantly more frequent in controls than HCC patients. It is worthy to mention that studies on the genetic association between OAS1 rs10774671 polymorphism and human cancer are scarce or even absent; so we cannot set an ethnic correlation of such phenomenon.
The combined analysis indicates that the risk of HCC significantly increases with the presence of each unfavorable genotype (LMP-7 CA/AA genotypes, IL28B rs12979860 CT/TT genotypes) together with CMV viremia (probability of HCC risk for combined factors is 0.9 versus 0.14, 0.07, and 0.07 for IL28B, LMP-7, or CMV, respectively), with IL28B rs12979860 having the most influence on the progress to HCC. These data suggest that the 2 SNPs and the coinfection in concert have a potential in predicting the risk of HCC development.
This study has a number of limitations: (1) the small sample size, which may limit our power to uncover a correlation between OAS1 SNPs and HCC development, (2) lack of data on the rate of disease progression before HCC onset (in HCC patients) or posterior to early stages of fibrosis (in F1-F2 patients). Such longitudinal study is necessary to test the sensitivity of our mathematical model, through measuring the percent of patients with high calculated probability of developing HCC who will later develop HCC, and (3) we examined HCC development among HCV-infected individuals with no information if our findings would be valid for HCC patients due to different etiologies.
In summary, our data support a role for IL28B and LMP-7 genotyping, together with CMV coinfection, in identifying HCV patients with hepatic fibrosis, who are at high risk for HCC development and present a mathematical model to calculate risk score for HCC incidence.
Footnotes
Acknowledgment
This work was supported by the Science and Technology Development Fund in Egypt (STDF) project No. 3365.
Author Disclosure Statement
No competing financial interests exist.