
Abstract
Previously, we demonstrated that Lactobacillus salivarius was more abundant in patients with rheumatoid arthritis (RA), an inflammatory autoimmune disease wherein the gut microbiota is altered, than in healthy individuals. However, the effect of L. salivarius in RA is unclear. Hence, we investigated the effect of L. salivarius isolated from patients with RA on collagen-induced arthritis (CIA) in mice. L. salivarius UCC118 or L. plantarum WCFS1 isolated from patients with RA was administered orally for 5 weeks, starting from 2 weeks before the induction of arthritis in DBA/1 mice. Clinical score progression, histological changes, serum cytokine concentrations, and the proportion of interleukin (IL)-17-producing T cells [T helper 17 (Th17)] and regulatory T cells (Tregs) in the spleen were evaluated. Bone erosion was evaluated by micro-computed tomography. CIA mice treated with either L. salivarius or L. plantarum showed lower arthritis scores, milder synovial infiltration, and less bone erosion when compared with phosphate-buffered, saline-treated CIA mice. Administration of L. salivarius and L. plantarum reduced the Th17 cell fraction and increased the Treg fraction. L. salivarius-treated CIA mice displayed a significant increase in serum anti-inflammatory IL-10 levels. Thus, pretreatment with L. salivarius could significantly improve CIA in mice and may help alleviate RA in a clinical setting.
Introduction
R
Several studies have shown dysbiosis of fecal microbiota in RA (Frizen 1994; Vaahtovuo and others 2008; Scher and others 2013). In a previous study, we demonstrated that patients with early RA exhibited elevation and population changes of fecal lactobacilli, where Lactobacillus salivarius, L. iners, and L. ruminis were found to be the predominant bacterial species (Liu and others 2013). Similarly, fecal 16s RNA sequencing showed that patients with new onset RA carried distinct gut microbiota, characterized by abundant L. salivarius, which was correlated with disease activity (Zhang and others 2015). However, it is unclear whether the abundance of L. salivarius affects the initiation or progression of autoimmunity.
Two contradictory roles of Lactobacillus have been proposed. The first hypothesis is that Lactobacillus spp. can confer a range of beneficial effects on the host. For example, L. casei and L. delbrueckii have been shown to attenuate swelling and inflammation of the joints in arthritis models (Kano and others 2002; Amdekar and others 2011). Furthermore, oral administration of lactobacilli can affect RA disease and inhibit its progression (Alipour and others 2014). However, several species of lactobacilli have been shown to potentially exacerbate disease onset; in particular, arthritis was successfully induced by L. casei cell walls in mice (Simelyte and others 2003). A spontaneous inflammatory arthritis model showed absence of arthritis development in germ-free conditions. However, disease exacerbation was observed when mice were exposed to L. bifidus (Abdollahi-Roodsaz and others 2008).
The aim of this study was to investigate the effects of L. salivarius on RA. Collagen-induced arthritis (CIA) is the most utilized experimental arthritis model. As in RA, CIA features imbalance in an T helper 17 (Th17) cell and regulatory T cell (Treg) populations (Rogier and others 2015). Proinflammatory cytokines, such as tumor necrosis factor-α (TNF-α) and interleukin (IL)-17, are abundantly expressed in the serum of mice with CIA. Blockage of these molecules has been shown to reduce disease severity (Zhu and Qian 2012; Schmidt and others 2013).
In this study, L. salivarius UCC118 was isolated from patients with RA and transferred to mice by oral gavages. Arthritis in mice was then induced with collagen II to observe the prophylactic or pathogenic effect of L. salivarius.
Methods
Isolation of Lactobacillus salivarius UCC118 from feces of patients with RA
Fecal samples were collected from 15 patients (3 men and 12 women) with early RA at the Southwest Hospital. None of the patients had received antibiotics within 2 months before stool collection, and none had history of gastroenteritis or particular dietary habits. Freshly collected fecal samples (5 g) were suspended in phosphate-buffered saline (PBS) and serially diluted by plating on de Man, Rogosa, and Sharpe (MRS) agar (Merck). Plates were incubated at 37°C under anaerobic conditions for 48 h (Smelt and others 2012). Dominant slimy colonies were observed following incubation, propagated in MRS broth (Merck), and incubated under the same conditions. Gram staining was performed for the culture and cell morphology was assessed.
Potential L. salivarius colonies were selected and investigated further for genotypic identification. The sequences were verified against GenBank database entries to identify their classification and genera. Stock powder of L. salivarius UCC118 was prepared. L. plantarum WCFS1 was also prepared and propagated using the same methods (Dhewa and others 2014).
Animals
Twenty-four DBA/1 mice aged 8 weeks and weighing 18–20 g were obtained from the Shanghai SLAC Laboratory Animal Corporation (Production License No. 2007000573633). The mice were housed under specific pathogen-free conditions at the Department of Laboratory Animal Science, Third Military Medical University, with 44%–65% humidity, temperature of 22–26°C, and 12 h light/dark cycle. The mice were bred in-house for at least 3 generations to ensure adaptation to housing conditions. Animal protocols were approved by the Animal Care and Use Committee of the Southwest Hospital, Third Military Medical University.
Induction and assessment of arthritis
Arthritis in DBA/1 mice was induced with a subcutaneous injection of 2 mg/mL bovine type II collagen (CII; Chondrex) emulsified in complete Freund's adjuvant (Chondrex) at day 0. On day 21, the mice received a subcutaneous booster dose of 100 μL of CII without complete Freund's adjuvant.
Induction of arthritis and disease severity were assessed by 2 investigators blinded to the treatment groups, using a previously established CIA scoring system for subjective evaluation of arthritis severity (Brand and others 2007), as follows: 0, no evidence of swelling or erythema; 1, erythema and slight swelling confined to the tarsals or ankle joint; 2, erythema and mild swelling extending from the ankle to the tarsals; 3, erythema and moderate swelling extending from the ankle to the metatarsal joints; 4, erythema and severe swelling encompassing the ankle, foot, and digits. The maximum total score for the 4 limbs of each animal was 16.
Experimental design
Lactobacilli were propagated on MRS agar (Merck). Incubations were performed anaerobically at 37°C for 48 h. A probiotics powder containing 1 × 1011 cfu/g of L. salivarius or L. plantarum was prepared. Feeding with the probiotics was initiated 2 weeks before the primary CII injection and continued for 5 weeks. The mice were fed 3 times per week. The amount of L. salivarius or L. plantarum given was 1 × 1010 cfu/dose (in 1 mL PBS) per mouse in all feeding protocols. The mice were divided into 4 groups according to their feeding regime (N = 6): (1) L. salivarius-treated CIA; (2) L. plantarum-treated CIA; (3) PBS-treated CIA; and (4) normal control mice without CIA.
Histological analysis
At the conclusion of the study period, the mice were euthanized with diethyl ether. Histopathological analysis was performed on the right hind knee joint of the mice. Briefly, 5 μm thick sections were stained with hematoxylin and eosin and the slides were analyzed under 100 × magnification.
Micro-computed tomography imaging
The front limbs of the mice were excised and fixed in 4% formalin for 2 days and subsequently scanned and reconstructed into a 3-dimensional (3D) structure using the Quantum FX MicroCT Imaging System (PerkinElmer) with a voxel size of 20 μm, X-ray tube voltage of 90 kV, current of 160 μA, and scan time of 3 min. X-ray projections were obtained at 0.5° intervals with a scanning angular rotation of 180°. 3D reconstructions were performed using Analyze 12.0 Software (Analyze Direct) to generate a visual representation of the results.
Enzyme-linked immunosorbent assays for cytokines
Blood samples were collected to isolate serum for cytokine analysis (Amdekar and others 2011). A standard sandwich cytokine enzyme-linked immunosorbent assay was performed to assess TNF-α, IL-17 (a proinflammatory cytokine), and IL-10 (an anti-inflammatory cytokine) levels in the serum according to the manufacturer's instructions (Ebioscience). Absorbance values were recorded at 450 nm.
Flow cytometry analysis
Single-cell suspensions were prepared from the spleen. Cells were stained with fluorochrome-conjugated antibodies and analyzed using an Accuri C6 flow cytometer (BD Biosciences). The antibodies used in the analysis were as follows: fluorescein isothiocyanate-conjugated anti-CD4, and Alexa 647-conjugated anti-IL-17 (Biolegend), and the Mouse Regulatory T Cell Staining Kit (eBioscience).
Statistical analysis
Results are expressed as the mean ± standard error of mean from 6 mice per group. The incidence of arthritis was compared between groups using Fisher's exact test. Clinical scores were comparisons between groups were performed using the Wilcoxon signed-rank test. Serum cytokines and flow cytometry data were compared using one-way analysis of variance. Data were analyzed using SPSS 17 software (IBM) and P < 0.05 was considered statistically significant.
Results
In this study, both L. salivarius and L. plantarum showed protective effects against chronic inflammations associated with CIA in mice.
Lactobacillus-treated mice had reduced arthritis scores
The incidence of arthritis was lower in mice that received oral administration of L. salivarius (P = 0.016) or L. plantarum (P = 0.017) than in PBS-treated CIA mice (Fig. 1a). Furthermore, we found that the L. salivarius group displayed lower arthritis incidence (P = 0.01) than the L. plantarum group. Arthritis symptoms were first visible at day 27 post-CII injection in all groups, except in normal mice, which were not injected with CII. All mice in the PBS group had arthritis at day 35. However, 4 of the L. salivarius-treated mice did not display any arthritis symptoms.
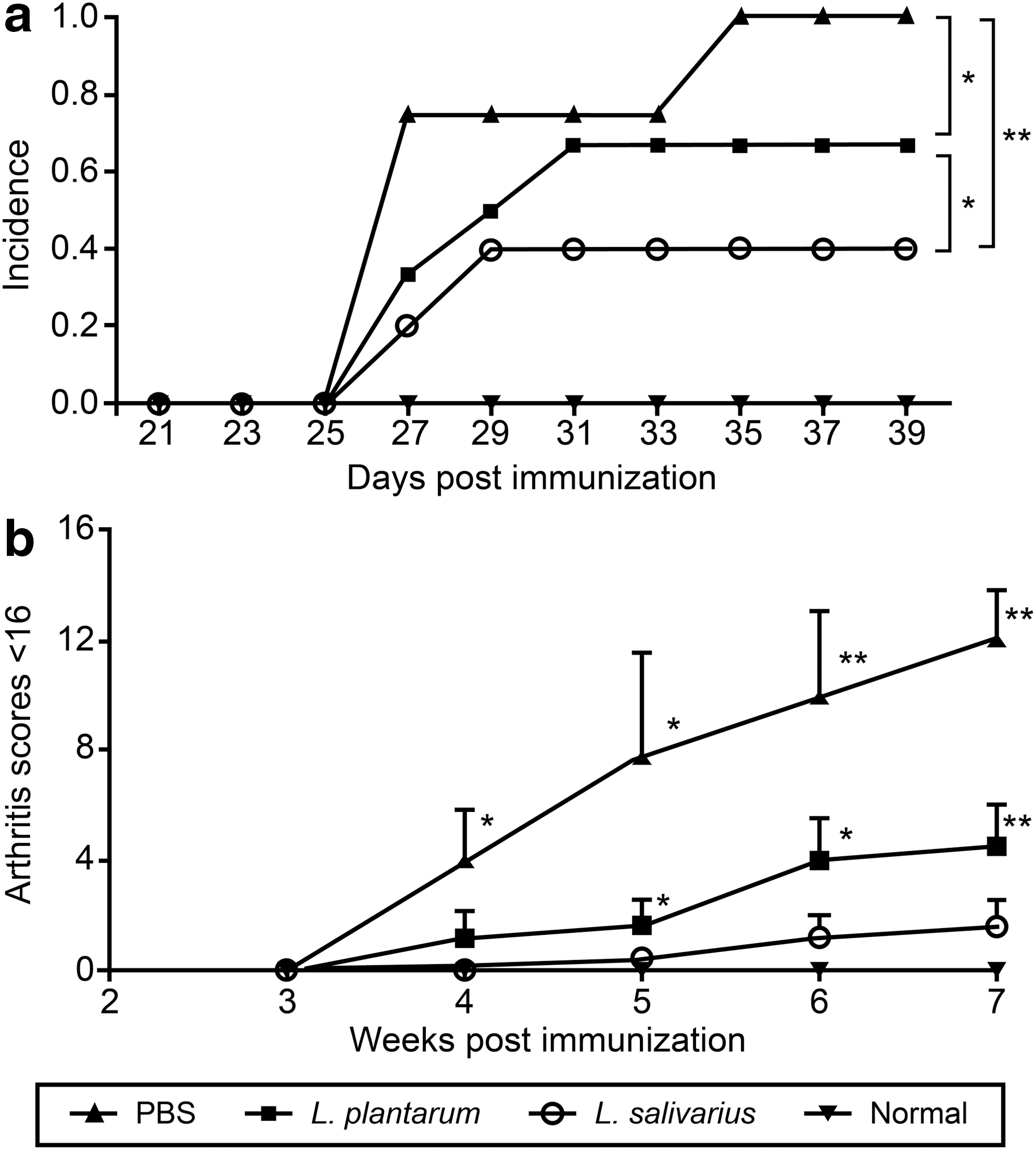
Arthritis score of collagen-induced arthritic DBA/1 mice.
Swelling and redness were prominent in PBS-treated CIA mice. Scores indicative of arthritis were observed in CIA mice treated with L. salivarius, but they were less severe than the scores observed in L. plantarum-treated (P = 0.038) or PBS-treated (P = 0.019) mice. At week 7, the arthritis score was significantly lower in the L. salivarius group (1.6 ± 0.9) than in the PBS group (12.0 ± 1.8; P < 0.0005) (Fig. 1b).
Lactobacillus-treated mice displayed reduced inflammation by histopathology
Inflammation during CIA was associated with cellular infiltration, edema, granuloma formation, and bone destruction on day 49. Mice in the PBS group showed significant elevation of inflammatory infiltrates when compared to Lactobacillus-treated mice. Histopathological analysis revealed necrotic and degenerative changes in the PBS group, whereas the Lactobacillus-treated groups showed moderate destruction and less degeneration (Fig. 2).
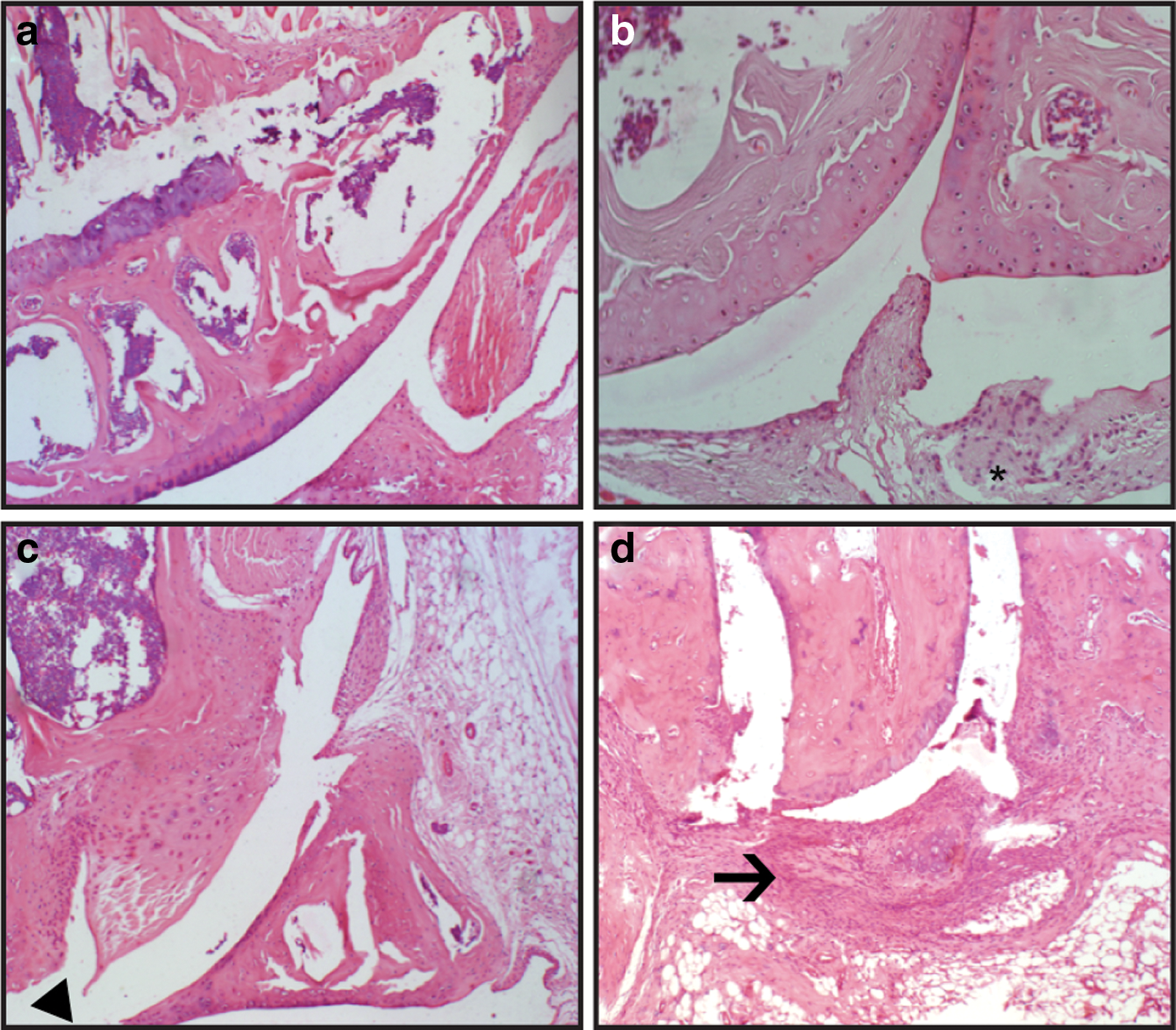
Histological analysis of the knee joints of CIA mice. Histopathological evaluation revealed no inflammation in the hind knee joint of normal mice
Micro-computed tomography scan demonstrated efficacy of Lactobacillus treatment in CIA mice
3D reconstructions of the paw bones of mice are shown in Fig. 3. Severe bone erosion was observed in PBS-treated CIA mice. L. salivarius-treated CIA mice showed moderate joint destruction when compared to PBS and L. plantarum-treated mice.
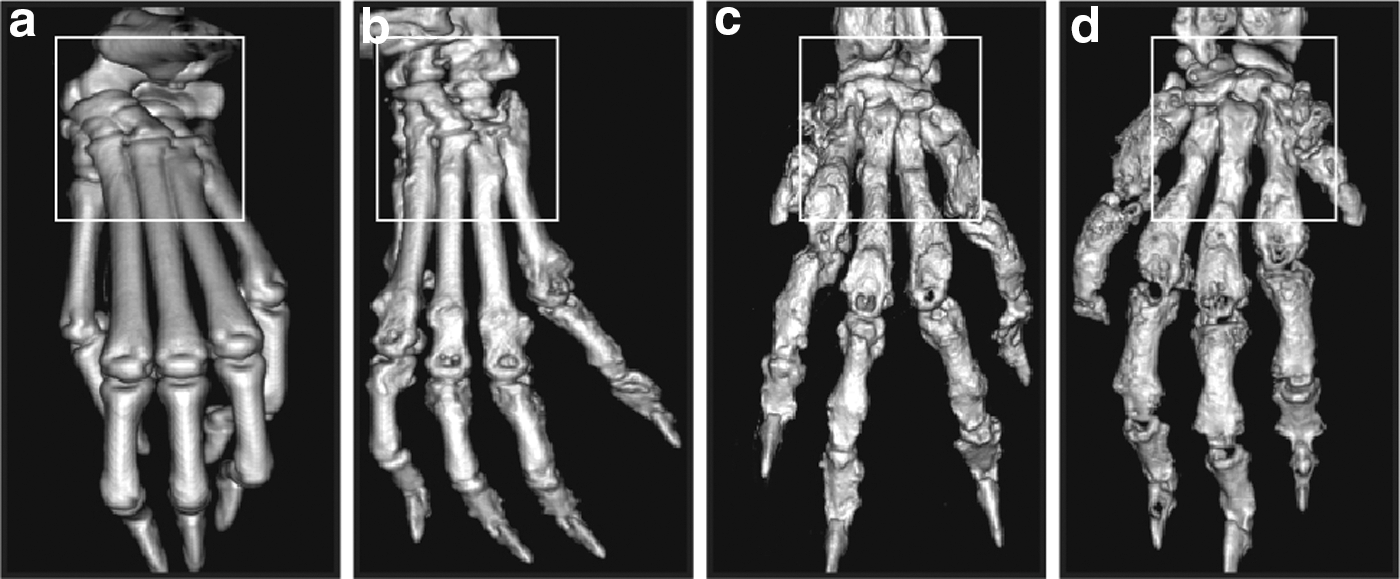
3D longitudinal visualization of bone changes. Representative 3D reconstructions of micro-CT images of joint tissues of the front limbs of normal control
Lactobacillus treatment modulated anti- and proinflammatory cytokine levels
The level of the anti-inflammatory cytokine IL-10 was significantly increased by Lactobacillus treatment. The IL-10 concentrations for the L. salivarius and L. plantarum groups were 263.03 ± 20.55 and 251.06 ± 46.19 pg/mL, respectively, which were higher than that observed in the normal group (97.25 ± 10.90; P < 0.05). The IL-10 concentration was significantly higher in L. salivarius-treated mice than in PBS-treated mice (185.03 ± 10.91; P = 0.48) (Fig. 4a). Among all treatment groups, mice in the PBS group displayed the highest concentration of the pro-inflammatory cytokine IL-17. L. salivarius and L. plantarum treatments decreased IL-17 and TNF-α cytokine levels, but these reductions were not statistically significant (Fig. 4b, c).

Lactobacillus treatment modulates anti- and proinflammatory cytokine levels. Serum levels of the anti-inflammatory cytokine IL-10
Lactobacillus treatment reduced Th17 and increased Treg cell proportions
L. salivarius and L. plantarum reduced Th17 and increase Treg proportions in the spleen. Both strains significantly decreased Th17 (CD4+IL17+) (Fig. 5a) and increased Treg (CD4+CD25+Foxp3+) (Fig. 5b) cell subsets, but these changes were more apparent in the L. salivarius group. The proportion of CD4+IL17+ cells in the spleen was higher in the PBS group (2.11 ± 0.26) than in the L. salivarius (0.55 ± 0.05; P < 0.0005) and L. plantarum (0.76 ± 0.08; P < 0.0005) groups. Mice in the L. plantarum group displayed significantly higher CD4+IL17+ cell proportion than those in the normal group (P = 0.012). However, no significant difference in CD4+IL17+ cell proportion was observed between the L. salivarius and normal groups. The proportion of CD4+CD25+Foxp3+ cells was lower in the PBS group (7.00 ± 0.67) than in the L. salivarius (11.18 ± 1.24; P = 0.013) and L. plantarum (11.11 ± 1.02; P = 0.017) groups.

L. salivarius and L. plantarum affect splenic Th17 and Treg cell proportions in CIA. Proportions of Th17
Discussion
In this study, we investigated the effect of L. salivarius isolated from patients with RA on CIA in mice. Our results demonstrated that systemic administration of L. salivarius affected the development of CIA in mice. Previously, we found elevated presence of L. salivarius in patients with RA (Liu and others 2013). Recent studies demonstrated over-representation of L. salivarius in individuals with RA and that L. salivarius was present in increased numbers in very active cases of RA (Zhang and others 2015). However, the exact role of L. salivarius in RA remained clear.
Gut microbiota are a major environmental factor that can elicit autoimmune diseases in genetically susceptible individuals (Pasztoi and others 2011). Several species of lactobacilli have been reported to cause arthritis (Simelyte and others 2003). In a previous study, L. bifidus was found to induce joint swelling in germ-free mice (Abdollahi-Roodsaz and others 2008). However, probiotic supplementation was shown to improve inflammation and alleviate symptoms in patients with RA (Vaghef-Mehrabany and others 2013; Alipour and others 2014).
Although L. salivarius is believed to prevent disease in healthy individuals, immune homeostasis may be significantly perturbed by L. salivarius, contributing to the pathology of RA. Because studies on the role of L. salivarius in the development of RA are lacking, we utilized an arthritis mouse model to demonstrate the efficacy of L. salivarius treatment. Among available arthritis models, CIA is a promising model to evaluate the effectiveness of antiarthritic drugs and various agents for therapeutic strategies.
In this study, we demonstrated for the first time the ability of L. salivarius from patients with RA to attenuate the clinical symptoms of CIA. Oral administration of L. salivarius significantly decreased arthritis score. Swelling and reddening were mild in mice treated with L. salivarius and L. plantarum. Histopathological analysis corroborated these findings and showed less neutrophil infiltration in the knee joint of mice that received oral administration of L. salivarius or L. plantarum. Using 3D reconstruction of micro-computed tomography images of joint tissues, significant reduction of bone erosion was also observed in L. salivarius-treated mice when compared to arthritic control mice.
Thus, administration of L. salivarius isolated from patients with RA was found to improve CIA. We believe that L. salivarius regulated immune balance rather than perturbing immune homeostasis in CIA mice. Furthermore, the elevated number of lactobacilli in patients with RA may be a consequence of RA progression. Increased reactivity and number of fecal lactobacilli, and increased intestinal colonization in CIA mice may enhance their immunoregulatory properties. Different mechanisms have been proposed to explain the Lactobacillus-dependent amelioration of arthritis (So and others 2011; Vaghef-Mehrabany and others 2013; Wachi and others 2014).
In autoimmune diseases, an imbalance between pro- and anti-inflammatory cytokines favors the induction of autoimmunity, chronic inflammation, and joint damage. In our study, a protective effect of L. salivarius isolated from patients with RA was observed in CIA mice. This effect may not be associated with a reduction of TNF-α and IL-17 levels in the serum. Mice that received L. salivarius showed the lowest proinflammatory cytokine levels, but the differences were not statistically significant. However, an increase in IL-10 level was also observed in L. salivarius-treated CIA mice. IL-10 is a cytokine with potent anti-inflammatory activity and a potent macrophage deactivator(Feldman and others 2000). So cytokine IL-10 and Treg may be more important in the protection.
IL-17-producing Th17 cells appear to play a critical role in arthritis (Nakae and others 2003) and are thought to be a key driver of joint, cartilage, and bone damage (Joosten and others 2008). The balance between Th17 and IL-10-producing regulatory T cells (Tregs) is also critical in CIA (Gol-Ara and others 2012). Commensal bacteria have been found to influence T helper cell differentiation, immune responsiveness, and homeostasis (Kelly and Mulder 2012). Previous studies demonstrated that segmented filamentous bacteria (Lee and others 2011; Goto and others 2014) and bacteria in the Lachnospiraceae family (Wu and others 2016) can promote Th17 cell polarization, whereas Bacteroides fragilis (Wang and others 2014), Clostridium spp. (Atarashi and others 2011), and Bifidobacterium infantis (O'Mahony and others 2008) can induce Tregs accumulation.
Based on these findings, we analyzed changes in the Th17 and Treg balance in the spleen. Our study demonstrated that both species of lactobacilli were able to increase the proportion of Tregs and reduce the amount of Th17 cells in the spleen when compared to PBS-only treatment. Indeed, other studies have shown a close link between the anti-inflammatory capacities of lactobacilli and the generation of Tregs or IL-10 production (O'Hara and others 2006; Kwon and others 2010). Additionally, the effect of L. salivarius was only found to be modest and not as pronounced as that observed following L. plantarum treatment (Smelt and others 2012). L. plantarum WCFS1 directly affected the systemic immune system by skewing it toward the Treg phenotype.
Nevertheless, we found that L. salivarius from patients with RA had strong systemic immunomodulatory effects in CIA mice than L. plantarum WCFS1. What is more, disease incidence and arthritis score were increased in the L. plantarum-treated group compared with the L. salivarius-treated group, possibly because the changes were more apparent in the L. salivarius group compared with the L. plantarum group. Discrepancies in these findings may be also due to different source of strains used or differences between healthy and arthritic individuals. We plan to further explore this and other possible reasons for the observed difference between species in future studies.
In summary, our results suggest that L. salivarius from patients with RA can exhibit antiarthritic and anti-inflammatory effects in a CIA model. To our knowledge, this is the first report of systemic immune changes following treatment of an animal model of arthritis with bacteria isolated from patients with RA. We explored the basal immunomodulatory properties of L. salivarius in the arthritis model and demonstrated that L. salivarius treatment induced profound changes in the cellular adaptive immune response. However, the exact mechanism by which L. salivarius influences RA remains unclear. Further clinical investigations should confirm the efficacy of L. salivarius in RA to enable it to be utilized as a promising therapeutic strategy against arthritis.
Ethical Approval
The study involving human participants was in accordance with the ethical standards of the institutional research committee and with the 1964 Helsinki declaration and its later amendments or comparable ethical standards. Informed oral consent was obtained from all study participants. All applicable international, national, and/or institutional guidelines for the care and use of animals were followed.
Footnotes
Acknowledgment
This work was funded by Project 81072921, 81302582 of the National Nature Science Foundation of China (NSFC).
Authorship Contributions
Y.F. and L.W. conceived and designed the experiments. X.L. and H.W., performed part of the experiments. F.X.M., Q.Z., B.Z., and J.Z. supervised the experiments and gave advice. X.L., J.Z., and F.X.M. contributed reagents, materials, and/or equipment. X.L. wrote the main article. All authors reviewed the article.
Author Disclosure Statement
No competing financial interests exist.