
Abstract
Human CD4+ T regulatory cells (Tregs) are a population of phenotypically and functionally diverse cells that downregulate inflammatory and autoimmune responses. As Th17 cells play an important role in the pathogenesis of autoimmune diseases, it is critical to elucidate the mechanisms regulating these cells. In this study, we examined the molecular basis underlying the phenotypic and functional diversity of human Tregs expressing the ectonucleotidase CD39. CD4+CD25hiCD39+ Tregs inhibit the proliferative response and the secretion of IL-17 and IFN-γ of autologous CD4+ T effector cells, while CD4+CD25hiCD39− Tregs only suppress IFN-γ production. We demonstrate that activated human CD4+CD25hiCD39+ Tregs express the Th17-associated surface markers CCR6 and IL-23R, and phosphorylate the transcription factor Stat3. Moreover, suppression of IL-17 by CD4+CD25hiCD39+ Tregs occurs via a Stat3-dependent mechanism as inhibition of Stat3 activation in the CD39+ Tregs reverses their ability to suppress IL-17. CD4+CD25hiCD39− Tregs are not endowed with the ability to inhibit IL-17 as they do not upregulate CCR6 or the IL-23R, and furthermore, they secrete IL-17. Our findings provide the first evidence that human Treg functional diversity is matched to the type of immune response being regulated and reveal a new role for Stat3 in controlling Treg function.
Introduction
H
CD39 and CD73, ectonucleotidases responsible for hydrolysis of ATP to adenosine, are coexpressed on nearly all murine Tregs (Borsellino and others 2007; Deaglio and others 2007). In humans, by contrast, CD39 expression is highly variable and restricted to a subset of CD4+CD25hiFoxp3+ Tregs (Borsellino and others 2007; Fletcher and others 2009; Mandapathil and others 2009). CD39 and CD73 coexpression is observed in a low percentage of human Tregs (Dwyer and others 2010; Mandapathil and others 2010). The difference in expression of these ectonucleotidases between murine and human Treg cells underscores that extrapolation from murine to human Treg cells may not always be applicable. Furthermore, human, but not murine, Th17 cells were described as being resistant to Treg suppression (Annunziato and others 2002; Evans and others 2007). Interestingly, CD4+CD25hiCD39+ Tregs are endowed with the ability to suppress IL-17 production by CD4+ Teffs (Fletcher and others 2009; Ye and others 2011).
The Th17-associated chemokine receptor, CCR6, is expressed on human and mouse Tregs (Kleinewietfeld and others 2005; Acosta-Rodriguez and others 2007; Kluger and others 2014). CCR6+ Tregs express Foxp3, exhibit Treg functional properties, migrate in response to the CCR6 ligand CCL20, express memory cell markers, and produce IL-10. CCR6 was expressed on human Tregs that secrete IL-17 directly ex vivo (Ayyoub and others 2009) or on activation (Voo and others 2009).
The suppressive mechanisms used by Tregs to inhibit immune responses are complex and not completely understood. The suppressive mechanisms are cell–cell contact dependent and include the release of inhibitory cytokines (IL-10, TGF-β, or IL-35) (Nakamura and others 2001; Levings and others 2002; Li and others 2006, 2007; Collison and others 2007, 2009; Niedbala and others 2007; Chaturvedi and others 2011; Chaudhry and others 2011; Huber and others 2011), modulation of antigen-presenting cell function (CTLA-4) (Read and others 2000; Takahashi and others 2000; Oderup and others 2006; Onish and others 2008), cytolysis (granzymes and perforin) (Grossman and others 2004), generation of suppressive metabolites (adenosine) (Huang and others 1997; Fredholm and others 2003; Naganuma and others 2006), and metabolic disruption (consumption of IL-2) (de la Rosa and others 2004; Busse and others 2010). Tregs were also reported to inhibit Th17 cells via a latency-associated peptide (LAP)-dependent mechanism (Ye and others 2011). It is possible that Tregs use other mechanisms to inhibit specific T effector cell responses.
Several groups demonstrated that murine Tregs expressed canonical CD4+ T effector cell-associated transcription factors to control polarized Th1-, Th2-, and Th17-driven immune responses (Chaudhry and others 2009; Koch and others 2009; Zheng and others 2009). Murine Tregs upregulate the expression of the transcription factors T-bet, IRF4, and GATA-3, and Stat3 to efficiently suppress Th1 (Koch and others 2009), Th2 (Zheng and others 2009), and Th17 responses (Chaudhry and others 2009), respectively. Similar lineage-specific Tregs are yet to be identified in humans.
In the present study, we describe a subset of human CD39-expressing CD4+CD25hiFoxp3+ Tregs that upregulate expression of the Th17-associated chemokine receptor CCR6, upregulate the IL-23 cytokine receptor, and phosphorylate Stat3 on activation. These Tregs inhibit the proliferation, as well as IFN-γ and IL-17 production, of autologous CD4+ Teffs. We demonstrate that suppression of IL-17 production by these cells occurs via a Stat3-dependent mechanism. This study suggests that CD4+CD25hiCD39+CCR6+IL-23R+Foxp3+ Tregs represent human Th17-specific regulatory T cells (Treg17).
Materials and Methods
Isolation of peripheral blood mononuclear cells
Human subjects age 23–55 years old with no known autoimmune conditions were recruited from the campus of the University of Medicine and Dentistry of New Jersey-New Jersey Medical School (now Rutgers-NJMS) in Newark, NJ. Research protocols were approved by the Institutional Review Board of NJMS and performed in accordance with the regulatory guidelines. Written informed consent was obtained from each subject. Peripheral blood mononuclear cells (PBMC) were isolated by Ficoll-Histopaque gradient centrifugation of heparinized venous blood as described in Liu and Rohowsky-Kochan (2008).
Antibodies
The following fluorochrome-conjugated mouse monoclonal antibodies specific for human markers were used in this study. APC-Cy7-anti-CD4 (clone RPA-T4), Pacific Blue-anti-Foxp3 (clone 259D), APC-anti-CD39 (clone TU66), and Brilliant Violet 421 anti-CD73 (clone AD2) were obtained from BioLegend (San Diego, CA). PE-anti-CD25 (clone M-A251), Alexa Fluor700-anti-CD25 (clone M-A251), and PE-Cy5-anti-CD152 (CTLA-4, clone BNI3) were obtained from BD Pharmingen (San Jose, CA). APC-anti-CCR6 (clone #53103) and FITC-anti-IL-23R (clone #218213) were obtained from R&D Systems (Minneapolis, MN). LEAF™ purified anti-human CD3 (clone HIT3a) Abs from BioLegend were used for activation.
Surface and intracellular phenotyping of Tregs
Resting, nonactivated, or “ex vivo isolated” PBMC were examined for expression of surface CD4, CD25, CD39, CD73, CCR6, IL-23R, and CTLA-4 using fluorochrome-conjugated Abs. For intracellular staining, cells were fixed, permeabilized using the Foxp3/Transcription Factor Staining Buffer Set (eBioscience; San Diego, CA), and stained for expression of Foxp3 and/or CTLA-4. For all staining protocols, the buffer was 0.1% bovine serum albumin (BSA)/phosphate-buffered saline (PBS) buffer, and heat-inactivated normal human serum was used for blocking. Immunofluorescence flow cytometry data were acquired using the 4-laser LSRII flow cytometer (BD Biosciences, San Jose, CA) in the NJMS Flow Cytometry Core Facility. Data were analyzed using FlowJo software (Version 7.6.4; TreeStar, Stanford, CA). All gates for individual markers were set based on fluorescence minus 1 (FMO) control technique, in which cells are stained with all of the markers except 1 and gates are drawn based on fluorescence of the missing fluorochrome within its individual FMO tube.
Isolation of T cell subsets by fluorescence-activated cell sorting
For Treg cell functional experiments, PBMC were stained with fluorochrome-conjugated CD4, CD25, and CD39 antibodies and separated into Treg and Teff populations by fluorescence-activated cell sorting (FACS) using the 4-laser FACSAria II cell sorter (BD Biosciences) in the NJMS Flow Cytometry Core Facility. Stringent gates were set using FMO, as described above, and CD4+ CD25hi Tregs were <1.5% of CD4+ T cells. The gating strategy for CD25 and sorting experiments is shown in Fig. 1. All cells were collected into 5-mL polypropylene tubes containing 40% fetal bovine serum in RPMI-1640 (Life Technologies, Grand Island, NY) complete medium supplemented with 1% penicillin–streptomycin–glutamine. The purity of the sorted cells was greater than 96%.

Distinct populations of CD4+CD25hi Tregs based on CD39 expression. PBMC were stained for surface expression of CD4, CD25, and CD39.
Suppression assay
CD4+CD25− Teffs (4 × 104 cells/well) were stimulated with 0.5 μg plate-bound anti-CD3 plus irradiated allogeneic CD3− antigen-presenting cells (APCs; 1 × 105 cells/well) and cocultured with or without sorted Tregs at a 1:1 ratio in 96-well round-bottomed plates for 6 days at 37°C in a humidified incubator with 5% CO2 (Fletcher and others 2009). To ensure HLA MHC class II mismatch, CD3− cells were collected from the PBMC of 3 donors, by depletion of CD3+ T cells by the Human CD3 MicroBeads kit (Miltenyi Biotec, Auburn, CA). The CD3− APCs from all 3 donors were mixed and irradiated at 20 Gy before culture. Proliferation was assessed after addition of thymidine (3H-TdR; 1 μCi/well) for 18 h and measured using the RackBeta Liquid Scintillation counter (LKB Wallac, Turku, Finland). For experiments in which proliferation was not measured, cells were harvested on day 6 and stained for expression of various markers, including CD4, CD25, CD39, CD73, CTLA4, CCR6, IL-23R, and Foxp3.
Cytokine quantitation by enzyme-linked immunosorbent assay
IL-17 was measured by enzyme-linked immunosorbent assay (ELISA) using an anti-human IL-17 antibody (Clone No. 41809, 2,000 ng/mL; R&D Systems) as the capture antibody and a biotinylated polyclonal anti-human antibody (75 ng/mL; R&D Systems) as the detection antibody with quantification by reference to an rIL-17 standard (R&D Systems). IFN-γ was measured using a commercially available OptEIA kit (BD Pharmingen), according to the manufacturer's instructions. For all the ELISAs, 1% BSA in PBS was used as the blocking and dilution buffer. All samples were measured in duplicate wells, OD values were measured at an absorbance of 450 nm, and means of the duplicates were compared to a standard curve of known cytokine concentrations. The sensitivity of the ELISA was 7.8 pg/mL for IL-17 and 4.7 pg/mL for IFN-γ.
Intracellular staining for Stat phosphorylation
Phosphorylation of Stat3 and Stat1 was analyzed using intracellular staining following 6 days of activation of FACS-sorted T cells as described (Purvis and others 2014). Cells were stained for surface expression of CD4 and CD25, fixed with 2% paraformaldehyde, permeabilized with 100% ice-cold methanol overnight, blocked with human serum, and stained with pStat antibodies (pY705;clone 4/P-Stat3 or pY701;clone4a/P-Stat1) both from BD Pharmingen for 1 h. Data were acquired using the LSRII flow cytometer and analyzed using FlowJo software, as described above.
Stat3 inhibition
CD4+CD25− Teffs and CD4+CD25hi T cells with or without CD39 expression were isolated by FACS, as described above. FACS-sorted CD4+ T cells and Tregs were then incubated at 37°C for 16 h in the absence or presence of Stattic V, a small molecule inhibitor of Stat3 activation and dimerization (50 ng/mL; Sigma-Aldrich). Following pretreatment with Stattic V, treated and untreated CD4+CD25− Teffs, CD4+CD25hiCD39+ Tregs, and CD4+CD25hiCD39− T cells (4 × 104 cells/well) were cultured in anti-CD3-coated wells in the presence of irradiated allogeneic CD3− APCs (1 × 105/well). Untreated CD4+CD25− Teffs were also cultured with treated and untreated CD4+CD25hiCD39+ Treg cells and CD4+CD25hiCD39− T cells at a 1:1 ratio. After 5 days, cells were harvested and stained for analysis of Stat3 and Stat1 phosphorylation by flow cytometry, and supernatants were collected for analysis of IFN-γ and IL-17 production by ELISA.
Statistical analysis
Data were analyzed by looking at mean differences in fluorescence, cytokine levels and suppression of proliferation between the different cell populations or culture conditions using a repeated measures 1-way ANOVA (F-test) with a 1-tailed alpha level of 0.05 or Student's t-test, as appropriate, using Prism 6.0 software (GraphPad Software, Inc., La Jolla, CA). Tukey's posttest was used to compare pairs of data sets following ANOVA. All experiments were analyzed utilizing appropriate statistical tests for parametric and nonparametric analyses on consultation with the biostatistics core facility at NJMS.
Results
CD39+human Tregs suppress Teff proliferation, IFN-γ, and IL-17 production
To access the functional role of CD39 on Tregs, we examined the ability of FACS-sorted CD4+CD25hiCD39+ and CD4+CD25hiCD39− Tregs to suppress proliferation and cytokine production of CD4+CD25− Teffs. Human CD4+ T cells were sorted into highly purified CD4+CD25-, CD4+CD25hiCD39+, CD4+CD25hiCD39−, and CD4+CD25med/loCD39+ T cell populations, using the gating strategy shown in Fig. 1A. Purity of the sorted populations was >98% (Fig. 1B). Foxp3 expression was higher in the CD4+CD25hiCD39+ compared to the CD4+CD25hiCD39− T cells (Fig. 1C). CD4+CD25hiCD39+ T cells suppressed the proliferation of CD4+CD25− Teffs (mean ± standard error of the mean, 74% ± 12%), but CD4+CD25hiCD39− T cells were not capable of suppressing this response (Fig. 2A). To determine whether expression of CD39 or high CD25 expression was critical for the suppressive ability of these Tregs, we examined the ability of CD4+CD25med/loCD39+ T cells to inhibit the proliferation of CD4+CD25− Teffs. Similar to the CD4+CD25hiCD39− T cells, CD4+CD25med/loCD39+ T cells were unable to suppress proliferation of CD4+CD25− Teffs (Fig. 2B).

CD39+ Tregs suppress proliferation, IFN-γ production, and IL-17 production of CD4+CD25− Teffs. CD4+CD25− Teff cells, CD4+CD25hiCD39+ Treg cells, CD4+CD25hiCD39− Treg cells, and CD4+CD25med/loCD39+ T cells were collected by FACS.
We next examined the ability of the CD4+CD25hiCD39+ and CD4+CD25hiCD39− T cell subsets to inhibit cytokine production, as it has been demonstrated that proliferation and cytokine production are separate functions (Sojka and Fowell 2011). IFN-γ production by CD4+CD25− Teffs was inhibited both by the CD4+CD25hiCD39+ and CD4+CD25hiCD39− T cells (P < 0.001 and P < 0.01, respectively), with slightly stronger inhibition by the CD39+ Treg cells (Fig. 2C). Moreover, the CD4+CD25hiCD39+ T cells significantly inhibited IL-17 production by CD4+CD25− Teffs (P < 0.05), whereas the CD4+CD25hiCD39− T cells did not inhibit IL-17 production (Fig. 2C). Thus, CD39 expression on CD4+CD25hi T cells denotes a subset of Tregs that suppress IL-17 production, while inhibiting T cell proliferation and IFN-γ production, confirming a previous report (Fletcher and others 2009). In contrast, the CD4+CD25hiCD39− T cells have regulatory activity that is restricted to inhibition of IFN-γ production.
To compare the ability of the CD25-expressing CD4+ Tregs to secrete cytokines, we measured cytokine levels produced by CD4+CD25hiCD39+, CD4+CD25med/loCD39+, and CD4+CD25hiCD39− T cells on activation. The CD4+CD25hiCD39+ Tregs produced very low levels of IFN-γ and undetectable amounts of IL-17 (Fig. 2D). The CD4+CD25hiCD39− T cell subset produced higher (not significant) amounts of IFN-γ and significantly higher levels of IL-17 on activation (P < 0.05) compared to the CD4+CD25hiCD39+ Tregs. The nonregulatory CD4+CD25med/loCD39+ T cells produced significantly higher levels of IFN-γ (P < 0.05) and slightly but not significantly higher levels of IL-17 compared to the CD4+CD25hiCD39+ Tregs (Fig. 2D).
Phenotypic analysis of CD39+ human Tregs
The finding that human CD4+CD25hiCD39+ and CD4+CD25hiCD39− Tregs were functionally different prompted us to perform a phenotypic analysis on these regulatory subsets. CD39 expression was significantly higher on the CD4+CD25hi Tregs (44% ± 17%) in comparison to CD4+CD25− (2.8% ± 1%; P < 0.001) and CD4+CD25med/lo (9% ± 4%; P < 0.001) T cells (Fig. 3A, B). CD39 expression on CD4+CD25hi Tregs was highly variable between donors, with expression ranging from 15% to 76% (Fig. 3B). Analysis of the Treg-specific transcription factor Foxp3 revealed that CD4+CD25hiCD39+ Tregs (76% ± 16%) expressed significantly higher levels (P < 0.001) compared to CD4+CD25hiCD39− Tregs (56% ± 17%) (Fig. 3C). CD73 surface coexpression with CD39 was very low in CD4+CD25hi Tregs ranging from below 1% to 10% (Fig. 3D). CD4+CD25+ T cells include both CD4+CD25hi and CD4+CD25med/lo T cells. CD73 was expressed without CD39 on a minor fraction of CD25hi Tregs. We observed very low levels of intracellular CD73 in the different T cell subsets, with almost no coexpression of CD39 and intracellular CD73 (data not shown). The CD4+CD25hiCD39+ Tregs have a significantly higher (P < 0.05) percentage of CD27+ CD127− cells (82.0% ± 4.3%) than the CD4+CD25hiCD39− Tregs (66.1% ± 8.4%). CD45RO was expressed on almost all of the CD4+CD25hiCD39+ Treg cells and CD4+CD25hiCD39− Treg cells (data not shown).
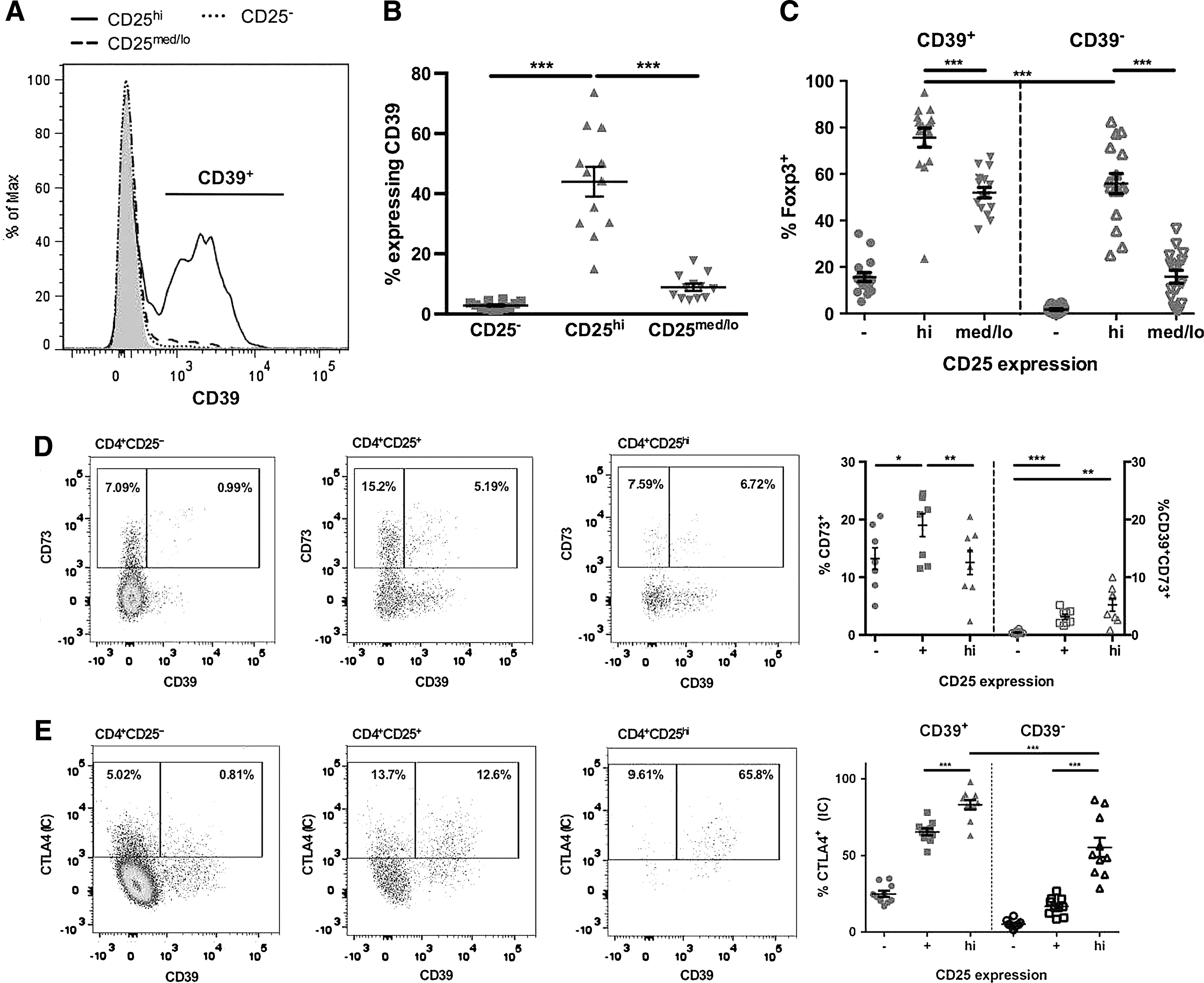
CD39 is expressed on human CD4+ Tregs with high Foxp3 expression. Resting, ex vivo PBMC were analyzed for expression of surface CD25, CD39, CD73, and intracellular CTLA4 and Foxp3.
As CTLA-4 is thought to be important for Treg cell function, we investigated the expression of surface and intracellular CTLA-4 on ex vivo isolated PBMC. CD4+CD25hi Tregs have low (7.3 ± 2.4), although significantly higher, surface CTLA-4 expression than CD4+CD25− (0.03 ± 0.1; P < 0.04) T cells. CD4+CD25hiCD39+ Tregs expressed significantly more intracellular CTLA-4 (83% ± 10%) compared to CD4+CD25hiCD39− Tregs (55% ± 20%; P < 0.001) (Fig. 3E). The phenotyping data indicate that CD39+ Tregs have a higher expression of Foxp3 and intracellular CTLA-4 but similar expression of CD45RO to CD39− Tregs.
CCR6 and IL-23R are expressed on the surface of suppressive CD39+ Treg cells following activation
Evidence from studies on murine Tregs shows that upregulation of chemokine receptors is critical for the ability of Tregs to suppress specific types of immune responses. In addition, 1 study classified human Foxp3+ human Tregs as Th1-like, Th2-like, and Th17-like Treg subsets based on their expression of subset-associated chemokine receptors and transcription factors (Duhen and others 2012). Since CD39+ Tregs suppress in vitro IL-17 secretion, we next investigated the expression of CCR6 and IL-23R on CD4+CD25hiCD39+ and CD4+CD25hiCD39− Tregs. CCR6 expression was very low on resting, ex vivo human CD4+CD25hi Tregs (2.9% ± 1.3%) and CD4+CD25− T cells (1.6% ± 0.5%). CD39+ Tregs markedly upregulated CCR6 expression following activation (P < 0.001) in comparison to the CD39− Treg cells (Fig. 4A, D). Furthermore, CD39+ Tregs strongly upregulated IL-23R expression on activation (P < 0.01) (Fig. 4B, E). CCR6 and IL-23R were coexpressed on significantly more CD39+ Tregs then CD39− Tregs (P < 0.001) (Fig. 4C, F). CD39− Tregs and CD4+ Teffs expressed low levels of either CCR6 or IL-23R or their coexpression following activation (Fig. 4D–F). These findings show that the CD39+ Tregs that suppress IL-17 production express the Th17-associated surface markers CCR6 and IL-23R.
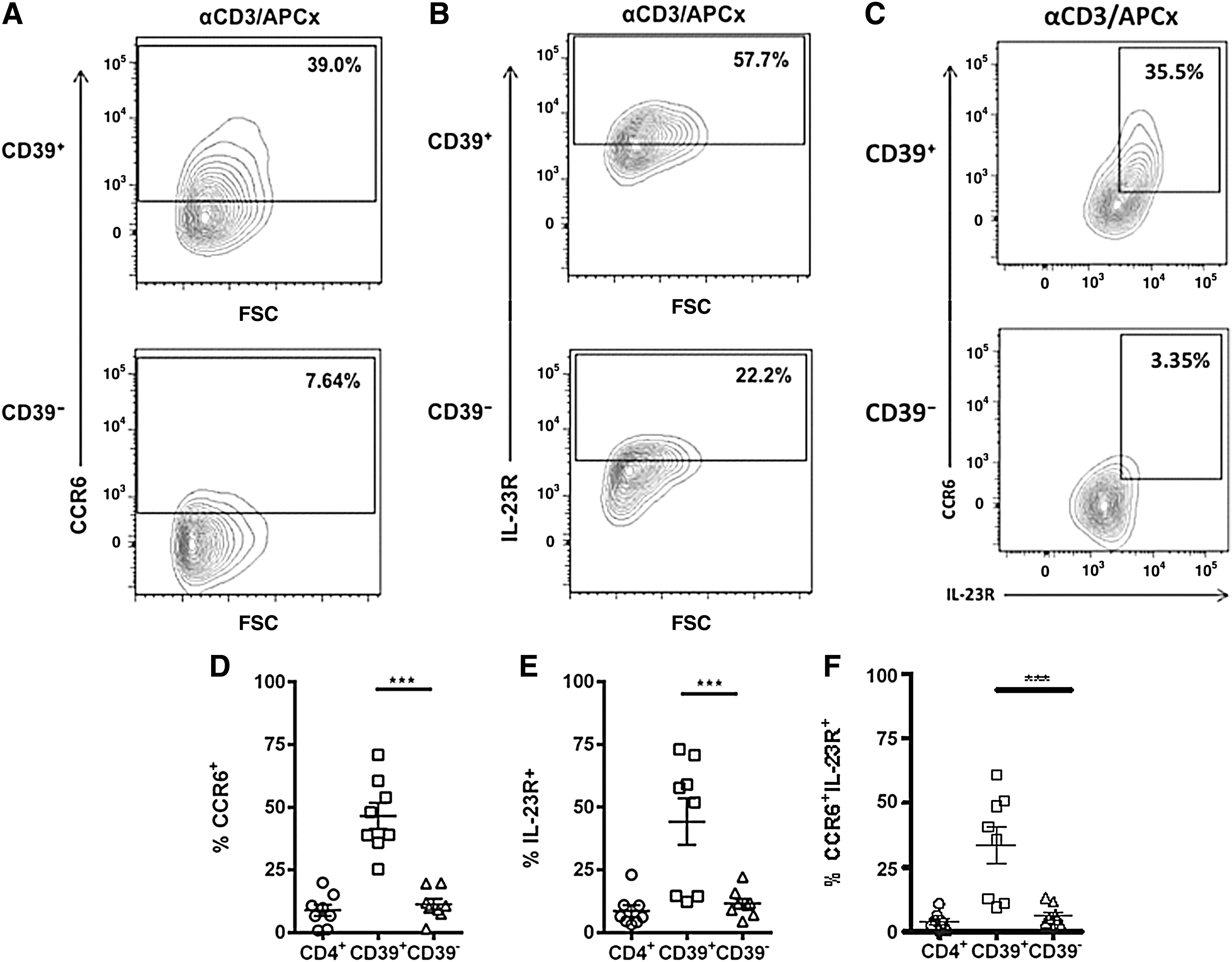
Human CD39+ Tregs express CCR6 and IL-23R following activation. CD4+CD25− Teffs, CD39+ Tregs, and CD39− Tregs were collected by FACS and cultured alone in the presence of plate-bound anti-CD3 and irradiated allogeneic APCs for 6 days. Following activation, cells were stained with fluorochrome-conjugated antibodies to CD4, CD25, CD39, CCR6, and IL-23R, and flow cytometry was used to analyze surface expression of CCR6
Stat3 phosphorylation contributes to CD39+ Treg-mediated suppression of IL-17
As Tregs may be specialized to suppress a specific CD4+ T cell subset by expressing the hallmark transcription factor of that specific Th subset (Chaudhry and others 2009; Koch and others 2009; Zheng and others 2009), we analyzed phosphorylation of the Th17- and Th1-associated transcription factors Stat3 and Stat1, in CD4+ Teffs and CD39+ Tregs activated alone and when cocultured together. CD4+ Teffs expressed similar levels of pStat3 and pStat1 regardless of whether they were activated alone or in coculture with CD39+ Tregs (Fig. 5A, C, and D). CD39+ Tregs activated alone phosphorylated Stat3, and slightly higher levels of pStat3 were seen following coculture with CD4+ Teffs. CD39+ Tregs activated alone phosphorylated Stat1, and significantly lower levels of pStat1 were seen following coculture with CD4+ Teffs (Fig. 5B–D). Analysis of the ratio of pStat3/pStat1 showed a significant (P < 0.05) preference toward Stat3 phosphorylation in the CD39+ Tregs following coculture with CD4+ Teffs, whereas the pStat3/pStat1 ratio did not significantly differ in the Teffs regardless of whether these cells were cultured alone or in coculture with CD39+ Tregs (Fig. 5E). These data suggest that the relative levels of pStat3 and pStat1 may contribute to the functional suppression mediated by CD39+ Tregs.
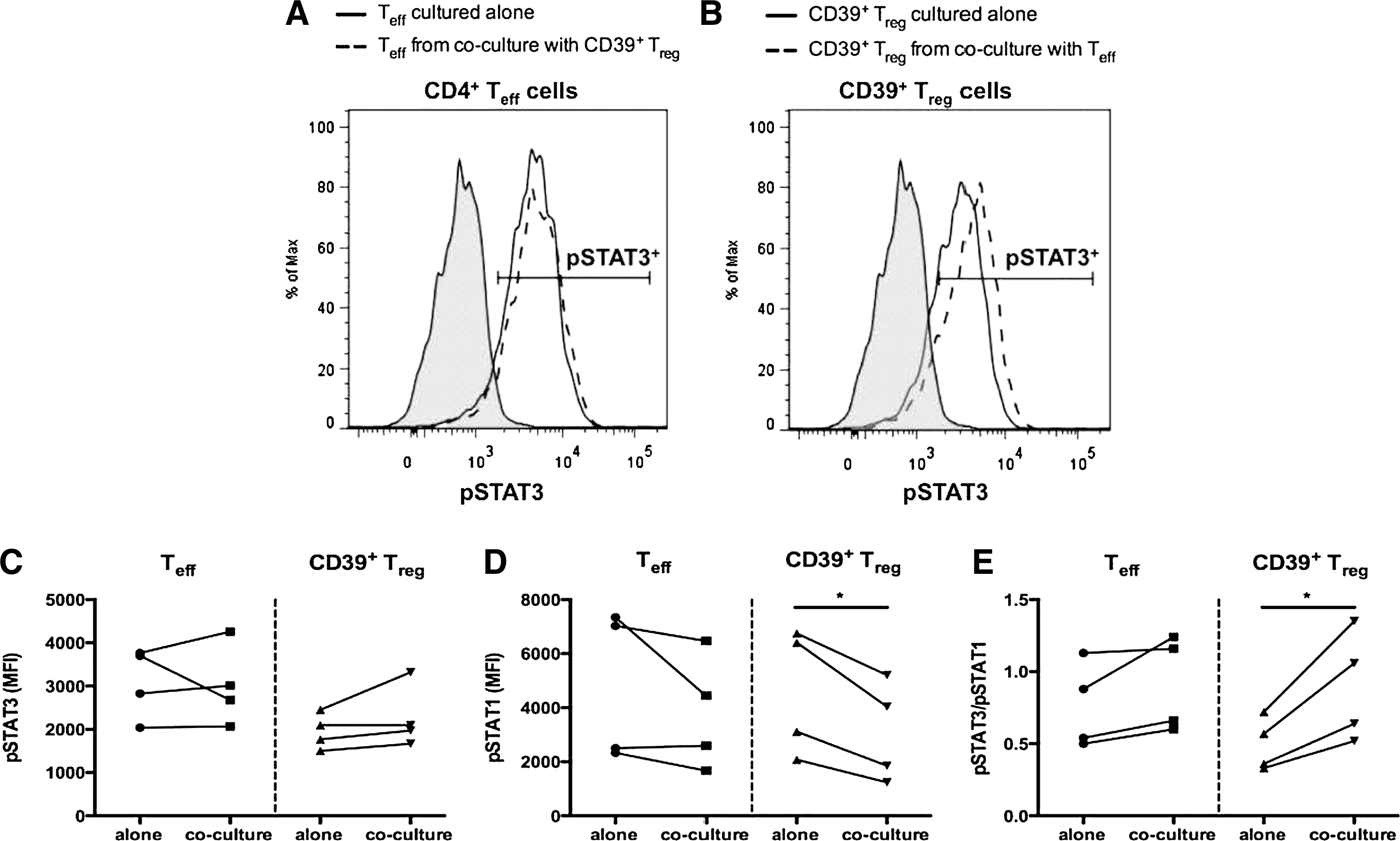
Levels of Stat3 and Stat1 phosphorylation in CD39+ Tregs. CD4+CD25− Teffs and CD4+CD25hiCD39+ Tregs were collected by FACS and cultured alone or in Teff:Treg cocultures at a 1:1 ratio for 6 days in the presence of plate-bound anti-CD3 and irradiated allogeneic APCs.
As Stat3 is associated with IL-17 production, we investigated the role of Stat3 activation in the production and suppression of IL-17. Sorted CD4+ CD25− Teffs, CD39+ Tregs, and CD39− Tregs were pretreated with the Stat3 inhibitor Stattic V for 16 h before activation. Stattic-treated and untreated CD39+ and CD39− Tregs were cultured alone and in 1:1 cocultures with untreated CD4+ CD25− Teffs and activated with plate-bound anti-CD3 and irradiated allogeneic APCs. In the absence of Stattic treatment, CD4+CD25hiCD39+ T cells significantly (P < 0.02) suppressed IL-17 production of CD4+CD25− Teffs, whereas CD4+CD25hiCD39− T cells slightly enhanced although not significantly IL-17 production when cultured with CD4+CD25− Teffs (Fig. 6A). IL-17 production by CD4+ Teffs cultured in the absence of Tregs was significantly inhibited (P < 0.01) by Stattic pretreatment (Fig. 6A). Likewise, production of IL-17 by the CD39− Tregs activated in the absence of CD4+CD25− Teffs was significantly (P < 0.02) inhibited by Stattic treatment (Fig. 6A). In contrast, inhibition of Stat3 activation in the CD39+ Tregs resulted in a significant (P < 0.02) increase in IL-17 in cocultures of CD4+CD25− Teffs and Stattic-pretreated CD39+ Tregs (Fig. 6A, B). We also observed a slight but not significant decrease in IL-17 production in cocultures of CD4+CD25− Teffs and Stattic-pretreated CD39− Tregs (Fig. 6A, B). The magnitude of reversal of IL-17 suppression by Stattic varied between donors, with the increase in IL-17 production ranging from 18% to 150%. These findings suggest that human CD39+ Tregs inhibit IL-17 in a Stat3-dependent mechanism, as activation of Stat3 within these cells is required for suppression of IL-17.

Inhibition of Stat3 activation in the CD39+ Tregs reverses their ability to suppress IL-17. CD4+CD25− Teffs, CD4+CD25hiCD39+ Tregs, and CD4+CD25hiCD39− Treg cells were collected by FACS and incubated with the Stat3 inhibitor Stattic V (50 ng/mL) for 16 h at 37°C. Stattic-treated and untreated CD4+CD25hiCD39+ Tregs and CD4+CD25hiCD39− Tregs were then cultured alone or at a 1:1 ratio with untreated CD4+CD25− Teffs for 6 days in the presence of plate-bound anti-CD3 and irradiated allogeneic APCs. Stattic-treated and untreated CD4+CD25− Teffs were cocultured alone under the same conditions. Supernatants were collected on day 6 and IL-17 production was measured by ELISA
Discussion
Human Tregs are a phenotypically and functionally heterogeneous population of CD4+ T cells. Classifying diverse Treg populations and understanding the mechanisms they use to regulate polarized immune responses are important for the development of disease-specific Treg-based immunotherapy. Murine Tregs express canonical CD4+ T effector cell-associated transcription factors to control polarized Th1-, Th2-, and Th17-driven immune responses (Chaudhry and others 2009; Koch and others 2009; Zheng and others 2009). In this study, we examined the molecular basis underlying the phenotypic and functional diversity of human Tregs. We demonstrate for the first time that human CD4+CD25hiCD39+ Tregs, which suppress IL-17 production by CD4+ Teff cells, express the Th17-associated surface markers CCR6 and IL-23R. Moreover, phosphorylation of Stat3 and Stat1 by CD4+CD25hiCD39+ Tregs contributes to their suppressive effect. Suppression of IL-17 by CD4+CD25hiCD39+ Tregs occurs via a Stat3-dependent mechanism as inhibition of Stat3 activation in the CD39+ Tregs reverses their ability to suppress IL-17. CD4+CD25hiCD39− Tregs are not endowed with the ability to inhibit IL-17, as they do not upregulate CCR6 or IL-23R, and furthermore, they secrete IL-17. Our study suggests that CD4+CD25hiCD39+CCR6+IL-23R+Foxp3+ Tregs represent human Th17-specific regulatory T cells (Treg17).
In searching for “signature” Treg markers, we observed that expression of the ectonucleotidase CD39 was largely limited to the CD25hi subset of CD4+ T cells, with highly variable expression between donors, as previously reported (Fletcher and others 2009; Dwyer and others 2010). In contrast, nearly all CD4+CD25hiFoxp3+ Treg cells were reported positive for CD39 (Mandapathil and others 2010). We found that CD4+CD25hiCD39+ Tregs expressed the highest levels of intracellular Foxp3 and CTLA-4 in comparison to CD4+CD25hiCD39− Tregs. Only a small percentage of CD25hi Tregs coexpressed surface CD39 and CD73, in agreement with other human studies (Borsellino and others 2007; Dwyer and others 2010; Rissiek and others 2015).
To assess the suppressive capacity of these Treg populations, we used FACS-sorted CD4+ Teffs and Tregs activated with plate-bound CD3 and irradiated allogeneic APC (Borsellino and others 2007; Fletcher and others 2009; Miyara and others 2009; Duhen and others 2012). We observed that CD4+CD25hiCD39+ and CD4+CD25hiCD39− T cells have distinct suppressive capabilities. CD4+CD25hiCD39+ Tregs have the ability to suppress the proliferative response, as well as IL-17 and IFN-γ production by CD4+ Teffs, whereas CD4+CD25hiCD39− Tregs only suppress IFN-γ production by CD4+ Teffs. These results suggest that Treg cell subsets use different mechanisms for suppressing Teff proliferation and cytokine production, as has been reported in the murine system (Sojka and Fowell 2011). One study showed that both CD4+CD25hiCD39+ and CD4+CD25hiCD39− Tregs suppressed proliferation and IFN-γ production and that the CD4+CD25hiCD39+ Tregs inhibited IL-17 production (Fletcher and others 2009).
The concept of lineage-specific Tregs is supported by studies demonstrating that mouse Tregs expressing T-bet inhibit Th1 cells, IRF4-expressing Tregs inhibit Th2 cell function, and pStat3-expressing Tregs inhibit Th17-mediated inflammation (Chaudhry and others 2009; Koch and others 2009; Zheng and others 2009). Whether specialization and differentiation of human Tregs occur has not been shown and is important to determine since differences exist between mouse and human Tregs regarding subset specificities and expression of markers. We have demonstrated that expression of CCR6 and IL-23R was specifically upregulated on CD39+ Tregs following activation. These data suggest that CD4+CD25hiCD39+CCR6+IL-23R+ Tregs may be specifically primed to interact with Th17 cells, as they may migrate with Th17 cells to sites of inflammation. Murine models have shown that CCR6-expressing Tregs migrate with Th17 cells to sites of inflammation (Yamazaki and others 2008; Turner and others 2010). Tregs that lacked surface expression of CCR6 resulted in impaired renal trafficking in an animal model of glomerulonephritis (Kluger and others 2014). Furthermore, expression of IL-23R on these Tregs may allow for their expansion at sites of inflammation, as IL-23 is important for Th17 cell expansion and survival (Wilson and others 2007; Liu and Rohowsky-Kochan 2008). Likewise, expression of the IL-23R on the Tregs may deprive Th17 cells of essential survival factors. To our knowledge, this is the first report demonstrating that coexpression of CCR6 and IL-23R is upregulated on human Tregs.
In addition to specific expression of Th17 surface markers on the CD39+ Tregs, our studies suggest that the relative levels of pStat3 and pStat1 may be important in inhibiting Th17 function. The increase in pStat3, in conjunction with the significant decrease in pStat1 in CD39+ Tregs cocultured with CD4+ Teffs, results in a significant increase in the pStat3 to pStat1 ratio. These data are in agreement with studies of mouse Tregs that report increased pStat3 in IL-17-suppressing Tregs (Koch and others 2009; Kluger and others 2014). Further studies using small interfering RNA silencing of pStat3 and pStat1 are warranted to confirm the role of these factors in CD39+ Treg cell-mediated suppression. Moreover, it remains to be determined which Stat-inducing cytokines are involved. No IL-6 or IL-23 was detected in the activated CD4+ Teff and CD39+ Treg cocultures and slight increases in IL-10 were seen (data not shown).
The functional role of pStat3 in mediating the suppressive function of CD39+ Tregs was demonstrated by inhibiting Stat3 activation in CD39+ Tregs. A salient finding of our study is the demonstration that suppression of IL-17 by CD4+CD25hiCD39+ Tregs occurs via a Stat3-dependent mechanism, as inhibition of Stat3 activation in the CD39+ Tregs reverses their ability to suppress IL-17. Treatment with a Stat3 inhibitor resulted in decreased IL-17 production by CD4+ Teffs and by CD4+CD25hiCD39− T cells, whereas suppression of IL-17 production by CD39+ Tregs was significantly reversed on treatment of Tregs with the Stat3 inhibitor. Hence, Stat3 activation is involved in IL-17 production as well as in suppressing IL-17 by Tregs. Our findings are in agreement with murine data describing that lineage-specific Treg17 cells expressed pStat3 for Treg cell suppression of IL-17 production (Chaudhry and others 2011; Kluger and others 2014, 2016). CCR6 expression on human Tregs also appears dependent on Stat3, as patients with dominant-negative Stat3 mutations have greatly reduced CCR6 levels (Kluger and others 2014). Further studies are needed to characterize the mechanism by which activation of Stat3 in CD39+ Tregs endows them with the ability to suppress Th17 responses. Unpublished data from our laboratory found that the immunosuppressive cytokine IL-35 does not appear to be involved.
Our studies differ from those of Goodman and others (2011), who reported that IL-6 inhibits Treg cell function and that the IL-6-mediated loss of Treg suppression requires phosphorylation of Stat3. They showed that the ability of Tregs to suppress proliferation of CD4+ Teffs in response to CD3 and CD28 crosslinking is restored when Stat3 is inhibited. Only Treg-mediated suppression of the proliferative response was examined. We specifically studied CD39+ Tregs and their ability to inhibit IL-17 production by Teff cells stimulated with anti-CD3 and APCs. No exogenous IL-6 was added nor was any IL-6 detected in our culture system.
Distinct subsets of human Foxp3+ human Tregs that phenotypically mirror effector Th cells were reported. Th1-like, Th2-like, and Th17-like Treg subsets were identified based on their expression of subset-associated chemokine receptors and mRNA expression of the transcription factors T-bet, GATA-3, and RORγt, respectively (Duhen and others 2012). No differences in mRNA expression of IRF4 and Stat3 were seen among the different Treg subsets. Although this study indicated a role for Treg expression of a Th17-associated transcription factor, the specificity and function of these cells and mechanism of suppression remain to be elucidated.
As Th17 cells are implicated in various autoimmune diseases, Th17-specific CD39+ Tregs may be utilized as treatment for these conditions. Due to the heterogeneity of this population, Treg cell transplantation has not yet been effective. We have shown that while CD4+CD25hiCD39+ Tregs suppressed Teff proliferation and production of IL-17, the CD4+CD25hi T cell subset that lacks CD39 expression proliferated and produced IL-17. If CD4+CD25hi Tregs were used to treat a Th17-skewed autoimmune disease such as multiple sclerosis (MS), production of IL-17 by CD4+CD25hiCD39− Tregs may counteract the beneficial effects of the CD39+ Tregs. Thus, since Tregs display a degree of plasticity, it is crucial to understand the complex interplay of the transcription factors in controlling Treg suppressive versus pathogenic function. It will be important to further investigate whether the impaired suppressive activity of CD4+CD25hiCD39+ Tregs in MS patients is associated with a dysfunction in Stat3 (Fletcher and others 2009). In the future, targeting of T helper cell subset-specific Tregs may be utilized to reduce specific types of inflammation in humans.
Footnotes
Acknowledgments
We thank the members of the NJMS Flow Cytometry Core Facility specifically, Tammy Galenkamp and Hong Liu for performing the cell sorting. We acknowledge the assistance of Jianfeng Wang in the preparation of all the figures. This work was supported by the National Institutes of Health, National Institute of Neurological Disorders, and Stroke T32 grant 5T32NS051157 and by a fellowship from Rutgers University, Graduate School for Biomedical Sciences (to J.R.M.-B.) as well as by a grant from the Foundation of UMDNJ and Dean's Biomedical Research Support Grant (C.M.R.-K.).
Author Disclosure Statement
No competing financial interests exist.