
Abstract
Major depressive disorder (MDD) is not a single disease, but a number of various ailments that form one entity. Psychomotor retardation, anhedonia, sleep disorders, an increased suicide risk, and anxiety are the main symptoms that often define the clinical diagnosis of depression. Interleukin-6 (IL-6), as one of the proinflammatory cytokines, seems to be overexpressed during certain mental disorders, including MDD. Overexpression of IL-6 in depression is thought to be a factor associated with bad prognosis and worse disease course. IL-6 may directly affect brain functioning and production of neurotransmitters; moreover, its concentration is correlated with certain clinical symptoms within the wide range of depressive symptomatology. Furthermore, there is a strong correlation between IL-6 synthesis and psychosomatic functioning of the patient. This article discusses potential sources and significance of IL-6 in the pathogenesis of depression.
Introduction
M
According to the immune-cytokine model of depression, there is an imbalance in immune response of type 1 and 2 helper T lymphocytes (TH1 and TH2, respectively), which leads to the overexpression of proinflammatory cytokines (Myint and others 2005; Gabbay and others 2009).
Interleukin-6 (IL-6) is secreted by both TH1 and TH2 cells (Szelényi 2001). After binding with the receptor, it affects cells in a variety of ways (März and others 1997; D'Arcangelo and others 2000). When it comes to the central nervous system, it has primarily a tendency to cause inflammation (Rohleder and others 2012); however, it needs to be emphasized that IL-6 can have both a proinflammatory and immunosuppressive effect, depending on biological conditions (Wang and others 2000, 2016; Henderson and others 2012; Jin and others 2013; Ohno and others 2016; Yeh and others 2016) (Fig. 1).
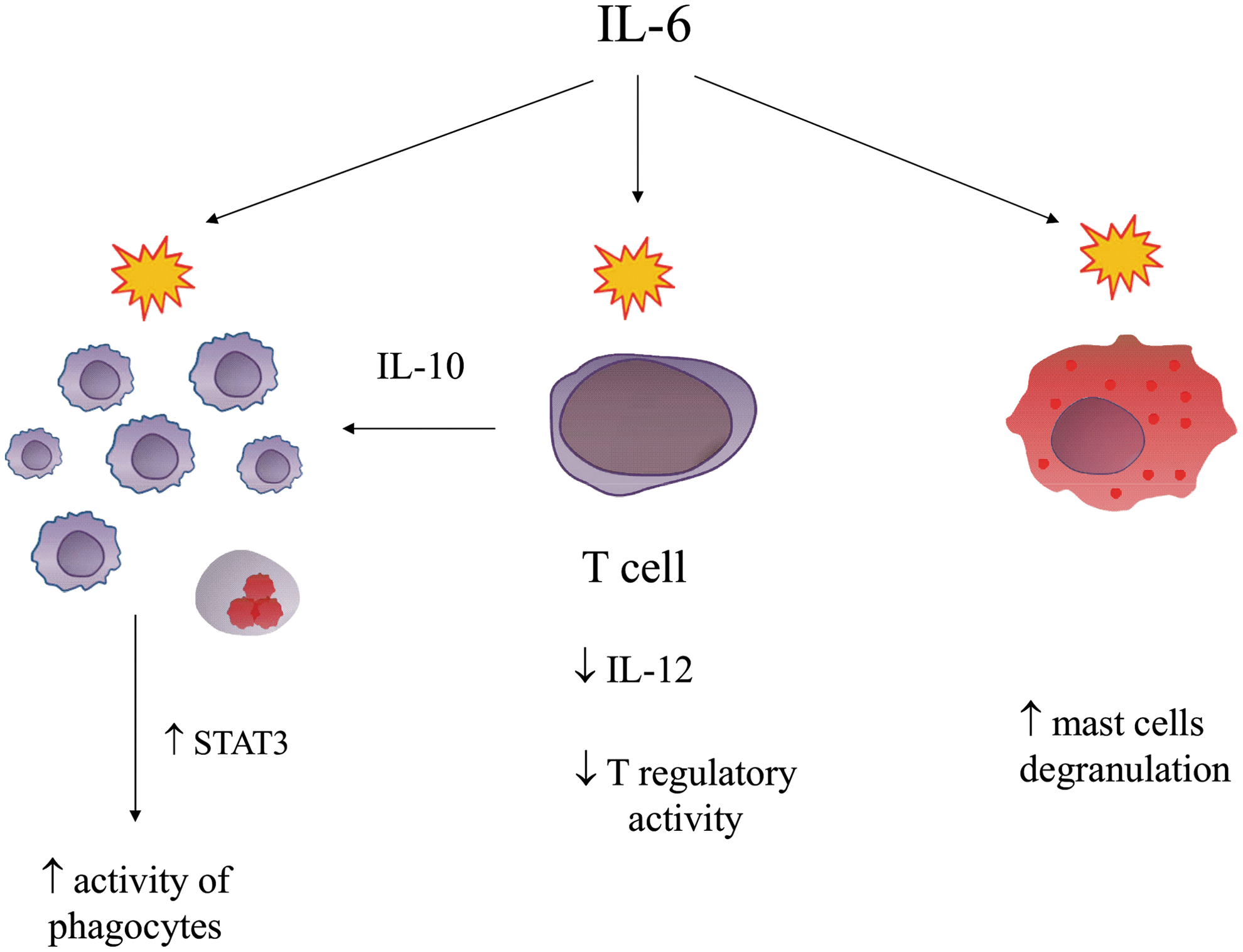
Biological properties of interleukin-6 in the regulation of immune response.
Numerous studies identify IL-6 to be a key proinflammatory factor in the pathogenesis of depression (Sjögren and others 2006; van Dooren and others 2016; Walsh and others 2016) and its elevated serum levels are under consideration as a future diagnostic marker (Chase and others 2016; Walsh and others 2016) and a predictive parameter of response to treatment (Manoharan and others 2016).
However, due to the inconclusive results of studies that seek to prove the concept of TH1-skewed immune response in the immune-cytokine model of depression (Huang and Lee 2007) and confirm elevated IL-6 levels (Stübner and others 1999; Carpenter and others 2004), this model cannot be now fully accepted. MDD involves a number of symptoms and not just 1 ailment (American Psychiatric Association 2013), and therefore a different impact of IL-6 in the pathogenesis of MDD may be not only due to the direct influence of the cytokine but also due to differences in the psychomotor state of patients (Goldsmith and others 2016).
Secretion of IL-6 in the Central Nervous System
Regulation of synthesis by monoamines
Norepinephrine (NE) is one of the basic neurotransmitters responsible for the pathogenesis of depression when it comes to certain elements of emotional regulation, such as the inability to cope with stressors, the feeling of inner aggression, and emotional tension (Gu and others 2016). According to recent data, NE induces inflammation by stimulating the synthesis of proinflammatory cytokines and metalloproteinases (Yang and others 2006; Slota and others 2015). In relation to IL-6, NE binds to beta adrenergic receptors and thus enhances intracellular reactive oxygen species (ROS) synthesis by activating the β-ADR-NAD(P)H oxidase system —NF-κB signal pathway. Similarly, binding of NE to beta-2 adrenergic receptors located on T CD8+ cells resulted in an increased IL-6 mRNA expression and elevated serum levels of certain proinflammatory cytokines (Li and others 2015).
Cholinergic stimulation influences the production of IL-6
After stimulation of the alpha7 subunit of nicotinic acetylcholine receptors located on the surface of astrocytes, the p38 kinase pathway is activated, which in turn promotes IL-6 production (Kalashnyk and others 2014). On an animal model, increased IL-6 concentrations were found to be correlated with decreased acetylcholine levels in the cerebrospinal fluid, which was associated with deteriorated cognitive functions (Zhou and others 2015). A completely different influence on memory was a result of the upregulation of metabotropic M1 receptors by IL-6 (Kim and others 2013).
Neuroglia—the active compartment altering the profile of secreted cytokines
Expression of IL-6 by astrocytes is induced through NFkB transcription factor (mediated by p38) and negatively controlled by the PI3K-mTOR pathway. Therefore, inhibition of PI3K-mTOR is necessary for IL-6 secretion (Codeluppi and others 2014). It seems to be dependent on the elevation of cytoplasmic concentrations of calcium ions Ca2+, which is a result of increased permeability of ryanodine receptors within the smooth endoplasmic reticulum, which enhance the secretion of IL-6 by exocytosis (Parpura and others 2010). Retinoic acid receptor-related orphan receptor-alpha is a neuroprotective factor and one of the nuclear transcriptional regulators, which can directly control astrocytic IL-6 synthesis (Journiac and others 2009).
The stimulation of Toll-like receptors on the cell membranes of astrocytes results in IL-6 overexpression, possibly due to irreversible inactivation of the Rho protein through glycosylation (Borysiewicz and others 2009). Increased synthesis of IL-6 and other proinflammatory cytokines may also be caused by altered mitochondrial function; adenosine 5′-triphosphate (ATP), among other functions, regulates the activity of microglia by stimulating the P2X7 receptors, which in turn increase microglial IL-6 synthesis (Shieh and others 2014). A study using magnetic resonance spectroscopy among depressed elderly patients confirmed the correlation between white matter concentrations of ATP and cognitive functions (Harper and others 2016).
During depressive episodes, the concentrations of stromal cell-derived factor 1 (SDF-1) and C-X-C chemokine receptor type 4 (CXCR4) were markedly elevated and were correlated with higher IL-6 levels and a more severe disease course (Ogłodek and others 2014). Microglia secrete IL-6 by interactions of SDF-1with CXCR4, which in turn leads to enhanced IL-6 mRNA expression and increased protein synthesis by activating the MAPK/ERK signal pathway and phosphatidylinositol-3 kinase (Lu and others 2009).
Importance of IL-6 for Functioning of the Central Nervous System
Cognitive functions
Deterioration of cognition, most frequently occurring as a slower information-processing rate, weakening of working memory, and impaired concentration, is a significant factor among depressed patients that limits their social and personal activities (He and others 2016).
The influence of immune system on thinking has not yet been fully elucidated. IL-6 affects brain activity already during early prenatal development. According to Brown and others (2014), IL-6 stimulation of astrocytes and neurons leads to significant changes in the kynurenine pathway, thus influencing tryptophan metabolism, and in the synthesis of certain glycerophospholipids. These changes can alter neurotransmitter secretion and biological properties of cell membranes. IL-6 may disrupt synapse formation by affecting their length and the morphology of dendrites, particularly in the CA1 and CA3 regions of hippocampus and the somatosensory cortex, which leads to neurodevelopmental disturbances and subsequent cognitive deficits (Wei and others 2012).
Elevated IL-6 levels regulate the expression of adhesion molecules and metalloproteinases that play a crucial role in ensuring the plasticity of the extracellular matrix, which is important in synapse formation (Suzuki and others 2010; Özkan and others 2014; Symeonidis and others 2014). Synapse formation is crucial for information processing in subsequent stages of morphogenesis—long-term potentiation and associative memory are regulated by intercellular interactions (Nelson and Alkon 2015). Clinically, deteriorated executive functions in vascular dementia were found to be correlated with elevated IL-6 levels and with decreased concentrations of blood Treg CD4+CD25+ cells (Guoping and others 2015).
The serotonergic system
5-Hydroxytryptamine (5-HT) was first isolated in 1948 as a factor released by platelets, and then, when 20 years later it was synthesized in the laboratory, it was named serotonin (Malone and Mann 1993). Currently, the mechanism of action of most antidepressant medications relies on the monoamine theory of depression that preconceives serotonergic system dysfunction (Fakhoury 2016). Children with neurodevelopmental disorders were found to have significantly higher levels of both IL-6 and 5-HT when compared with the control group (Yang and others 2015), and serotonin concentration in the platelets of patients with schizophrenia was correlated with the occurrence of symptoms of depression (Peitl and others 2016). Administration of IL-6 resulted in an increased release of 5-HT and tryptophan (TRP) in the region of hippocampus (Dwarkasing and others 2016).
The induction of depression by IL-6 may be due to its influence on serotonin transporter (SERT) because the levels of its mRNA and protein expression are significantly lower when stimulated by IL-6 (Kong and others 2015). Moreover, a correlation was found between genotypes of polymorphisms in IL-6 and SERT genes and symptoms of depression and anxiety (Udina and others 2013). The secretion of 5-HT in the brain when faced with stressors is affected by CRHR1 and CRHR2 receptors in raphe nuclei, which are activated by nerve impulses from the amygdalae through the stria terminalis after corticotrophin-releasing hormone (CRH) stimulation (Donner and others 2016).
Depressive Potential of IL-6
Appetite regulation
Eating disorders, including both increased and decreased appetite, are one of the additional criteria of MDD diagnosis (American Psychiatric Association 2013). A strong correlation between mood disorders and emotional expression is also connected with eating habits and food quality (Baskin and others 2015). A study conducted using functional magnetic resonance showed that central regulation of appetite and satiety in depression is affected by variable activity of the insular and orbitofrontal cortex, putamen, striatum, and ventral pallidum (Simmons and others 2016). Hormonal appetite control is affected by certain hormones of the gastrointestinal tract, such as ghrelin, and the metabolic activity of adipose tissue, which secretes leptin that induces the feeling of satiety (Yu and Kim 2012).
Ghrelin synthesis is not limited to the gastrointestinal tract. It is also secreted by hypothalamic neurons and its long-term expression increases appetite and weight gain (Qi and others 2015). That mechanism also seems to be affected by the serotonergic system because polymorphisms in the genes of SLC6A14, responsible for the bioavailability of tryptophan, and 5-HTR2C, encoding a G-coupled receptor, are associated with nutritional status of children (Miranda and others 2015). The importance of cytokines for the correct functioning of the appetite and satiety center is not yet fully understood, but IL-6 has a potential role to play there. Its elevated level and decreased concentration of the soluble form of its receptor (sIL-6R) were detected among patients suffering from anorexia nervosa (Solmi and others 2015).
The elevated IL-6 mRNA expression in hypothalamic and cerebellar neurons induced by glucagon-like peptide-1 by influencing calcium homeostasis of nerve cells led to a decreased food intake and stopping weight gain (Anesten and others 2016). Studies conducted among adolescent females suffering from anorexia nervosa reported a decreased level of peripheral blood IL-6 mRNA (Kahl and others 2004), whereas there was no correlation between the serum concentration of IL-6 and changes in clinical parameters assessed before and after the onset of treatment (Brambilla and others 1998).
Psychotic symptoms
A psychotic episode in a depressed patient is a diagnostic difficulty and has a negative influence on disease course of depression (Kim-Romo and others 2016). Despite few publications that concern the interactions between the immune system and psychotic symptoms during a depressive episode, IL-6 overexpression seems to be associated with both the psychotic and emotional components (Noto and others 2015; Harrison and others 2009). During exacerbation in the course of schizophrenia, a decrease in CD3+ T lymphocytes was observed (Maino and others 2007), and dopamine receptor D3 (DRD3) mRNA expression was significantly higher in T cells of these patients (Cui and others 2015).
Stimulation of DRD3 decreases the intracellular cyclic adenosine monophosphate (cAMP) concentration and phosphorylation of the extracellular signal-regulated kinase 2 (ERK2), which modify the activity of CD4+ lymphocytes (Franz and others 2015), whereas in neurons, it can also modify synaptic plasticity of deeper layers of the prefrontal cortex (Xing and others 2016).
An increased expression of S100B protein within the neuroglia was described among patients suffering from paranoid schizophrenia in the dorsolateral prefrontal cortex and pyramidal layer of hippocampus (Steiner and others 2008). S100B protein seems to promote hippocampal neuroregeneration during psychosis by activating the NF-κB transcription factor (Kögel and others 2004). Nevertheless, postmortem brain examinations of patients with affective disorders showed that the S100B protein has a completely different influence on the neuroglia because there it is correlated with a decreased hippocampal astrocyte density, particularly among depressed patients (Gos and others 2013).
When stimulated by the S100B protein, astrocytes secreted proinflammatory cytokines, such as TNF-alpha and inducible nitric oxide synthase, while IL-6 in a particular concentration range initially increased the expression of S100B (Reali and others 2012; de Souza and others 2013). S100B protein expression might also be affected by the serotonergic system by modifying intracellular cAMP levels, particularly through interactions with the 5-HT7a receptor (Stroth and Svenningsson 2015). Investigated on animal models of depression, elevated serum S100B levels were reported with their variable levels in the prefrontal cortex (Luo and others 2010). Interestingly, S100B stored intracellularly in natural killer (NK) and CD8+ lymphocytes can be released when these cells are stimulated (Steiner and others 2011).
NK cells with S100B overexpression were also found among patients with an acute episode of schizophrenia (Steiner and others 2012).
Peripheral blood S100B+ leukocytes can affect the permeability of the blood–brain barrier. The S100B protein is a marker of its dysfunction and IL-6 investigated on an animal model of psychosis was found to further increase its permeability by inducing NOX2 (NADPH oxidase 2 enzyme) overexpression, which indirectly activates certain metalloproteinases that degrade the extracellular matrix (Schiavone and others 2016). IL-6 downregulates the expression of adhesion proteins, such as epithelial cadherin, occludin, and claudin-5, by increasing intracellular ROS levels (Rochfort and others 2014). That regulates the permeability of the blood–brain barrier to a considerable degree and exposes the central nervous system to the uncontrolled inflow of stimulating factors (Fig. 2).
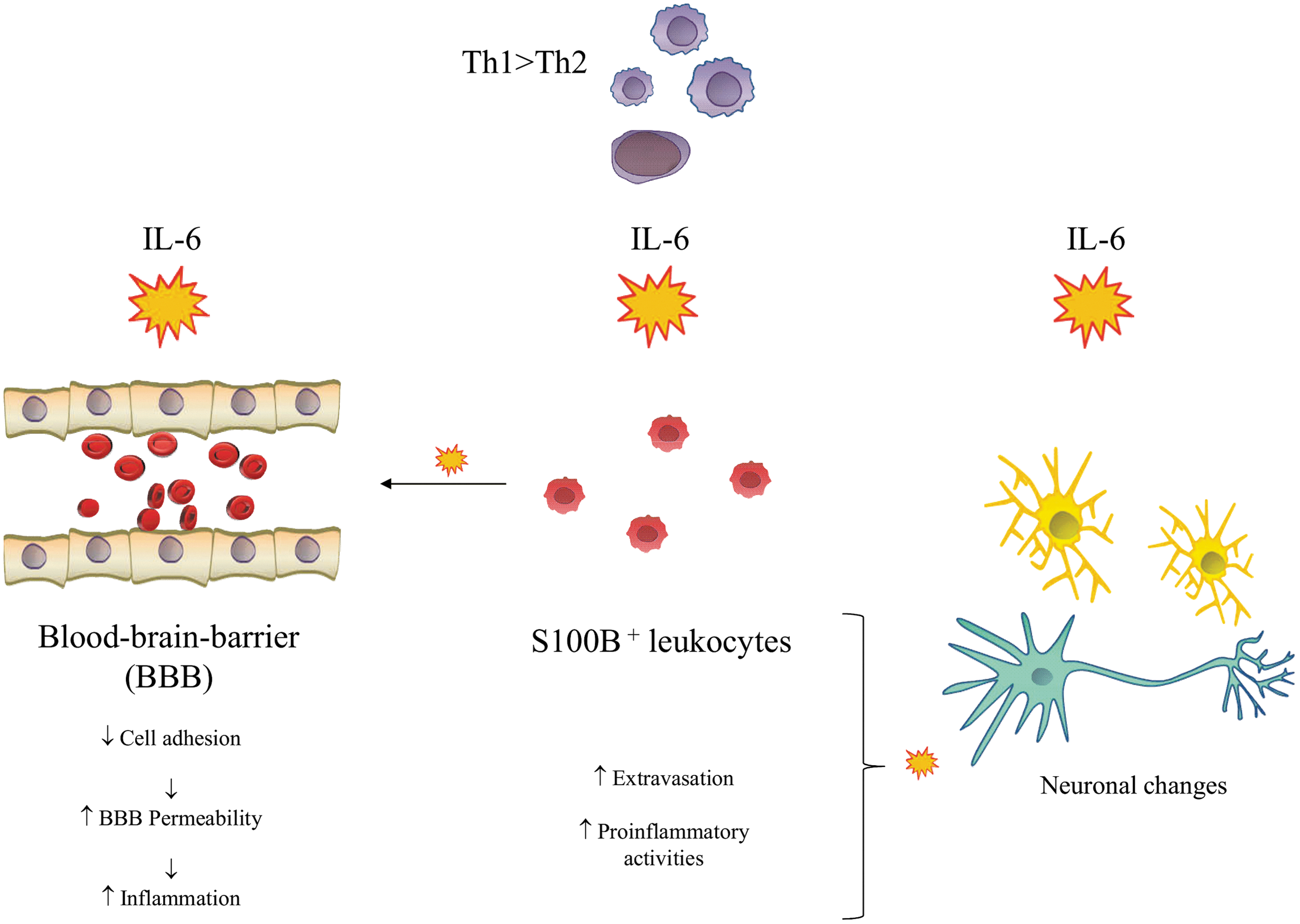
Differential proinflammatory effects of action of interleukin-6.
Plasticity of the emotional component
Anhedonia, one of the typical symptoms of depression, may cause hyperglycemia and increase the risk of diabetes mellitus type 2 (Carter and Swardfager 2016). Chronic inflammation was reported to be correlated with mastocyte activation and increased IL-6 production (Betto and others 2015). Its elevated levels in the course of induced acute inflammation were positively correlated with the intensity of negative emotions (Wright and others 2005). The undesirable influence of proinflammatory cytokines such as IL-6 on emotions probably stems from the increased activity of the anterior cingulate cortex (Carter and Swardfager 2016).
Anxiety occurs frequently among depressed patients and it impairs their functioning (Betto and others 2015). Elevated IL-6 levels were found to be positively correlated with the severity of anxiety disorder, but not with the severity of depressive symptoms (O'Donovan and others 2010). Central secretion of IL-6 might be an explanation for how it exerts such an influence on anxiety (Erta and others 2015). Stress increases IL-6 release from astrocytes located primarily in the periventricular region (Aniszewska and others 2015), which may be the cause of observed elevated IL-6 levels in the cerebrospinal fluid of depressed patients (Sasayama and others 2013; Kern and others 2014).
The threshold of pain
Chronic pain among depressed patients is a frequent ailment that deteriorates their quality of life. It can be located in various body parts, but most frequently headaches and flank or neck pain occur (Maneeton and others 2013). Numerous studies link MDD pathophysiology with concurrent chronic pain, and this mutual interaction exacerbates low mood and behavioral reactions (Zambito and others 2015). In psychiatric wards, up to 64% of patients suffer from depression with concurrent chronic pain (Bair and others 2003). The most frequent type of pain is headache, which is simultaneously an independent risk factor for suicidal thoughts and suicidal attempts (Ilgen and others 2008). Headache intensity is positively correlated with the severity of depression (Scanlon and others 2016).
Due to the axial MDD symptoms, emotional reactivity among depressive patients and the ability to experience pleasure remain impaired (Terry and others 2013). IL-6 overexpression and its elevated levels in case of surgical tooth extraction were found to amplify the experienced pain (Wang and others 2009). IL-6 was also reported to affect hypersensitivity to pain, both its affective and behavioral components (Edwards and others 2008). The rs1800795 polymorphism located in the promoter region of the IL-6 gene was positively correlated with concurrent pain among patients with exacerbation of depressive symptoms (Kovacs and others 2016).
Nutritional habits in depression
In certain phenotypes of depression, there is a greater inclination to frequent consumption of sweets, such as chocolate and coffee (Franco and others 2013), which probably aims to compensate for the low mood of the patient by stimulating the brain reward system and providing short-term satisfaction (Parker and others 2006).
Moreover, unhealthy food, including sweets, is currently considered to be a significant environmental factor that increases the prevalence of depressive symptoms (Mikolajczyk and others 2009; Benko and others 2011).
Tryptophan deficiency, present in depression, is the main argument of supporters of the serotonin theory, which explains the increased need for such types of food through the necessity to provide more substrates for the kynurenine pathway (Strasser and others 2016). It seems to be vital for correct functioning of the central nervous system, including its cognitive and emotional functions (Strasser and others 2016). Experimental studies reported that chocolate intake was correlated with pleasure and satisfaction, but this association depended on the percentage cocoa content: for extreme values, a reverse effect was observed, with a negative effect and personality traits (Macht and Mueller 2007). This result may be explained by the reaction to different cocoa content, with its high content tasting bitter, interpreted as an unpleasant stimulus.
Nevertheless, current research does not support this hypothesis as no relationship was reported between depression severity and stimuli after chocolate consumption (Scinska and others 2004).
On the other hand, low cocoa content is usually complemented by a greater amount of sugar, which even in dark chocolate comprises about 30% of its ingredients (di Renzo and others 2013). Alkaloids present in this food are in their chemically active form and their influence on gastrointestinal tract and nervous system seems to be confirmed (Franco and others 2013; Alañón and others 2016). Interestingly, flavonoids affect immune response through regulation of cytokine synthesis (Netea and others 2013). Dark chocolate intake altered the expression of proinflammatory cytokines under acute stress (Kuebler and others 2016).
Patients with depressive symptoms tend to eat sweet products with high-fat and simple carbohydrate content. This tendency during depressive episodes is considered to be a marker of personality traits due to the fact that it more frequently concerns self-critical nervous people, who thus try to relieve tension and anxiety (Parker and Crawford 2007; Sivanathan and others 2015). Among neurotic people, the concentration of IL-6 was substantially elevated and was positively correlated with lack of conscientiousness (Sutin and others 2010). However, in later studies investigating the association between IL-6 levels and the extent of neuroticism, there was a negative correlation between the 2 values, and additionally also with excessive concern with detail, but not the abovementioned lack of conscientiousness (Turiano and others 2013).
The explanation behind these associations does not need to focus only on character traits. As was noticed by Turiano and others (2013), the elevated levels of IL-6 were dependent on body–mass index (BMI), which suggests a positive correlation between excess weight and inflammation.
Lifestyle-induced chronic inflammation also seems to be closely connected with diet. Evolutionary aspect of eating habits changes the sugar intake, which means that with time, products with higher glycemic index are available, which affect the intestinal microbiota and may induce immune response (Spreadbury 2012). Gut flora seems to have a considerable influence on the central nervous system through cytokines, which may cause functional brain changes (Rogers and others 2016). Long-term exogenous glucose intake results in changes in intestinal microbiota and enhanced mRNA expression of proinflammatory cytokines, such as IL-1β and IL-6 (Mori and others 2015).
In vivo studies reported elevated IL-6 levels in response to induced transient hyperglycemia (Siklova and others 2015). High-fat and glucose content in sweets, coupled with sedentary lifestyle, facilitates uncontrolled growth of adipose tissue. Epidemiological data suggest that overweight people more frequently suffer from mood disorders, and obesity is a risk factor for depression (Roberts and others 2003; Simon and others 2006; Appelhan and others 2012). Excess consumption of sweets by patients with depression may increase IL-6 secretion through a complex cellular mechanism, which not only alters the profile of proinflammatory cytokines but also regulates the functioning of the central nervous system (Tam and Stepień 2011; de Rivero Vaccari and others 2014; Sathanoori and others 2015; Li and others 2016; Shirai and others 2016) (Fig. 3).
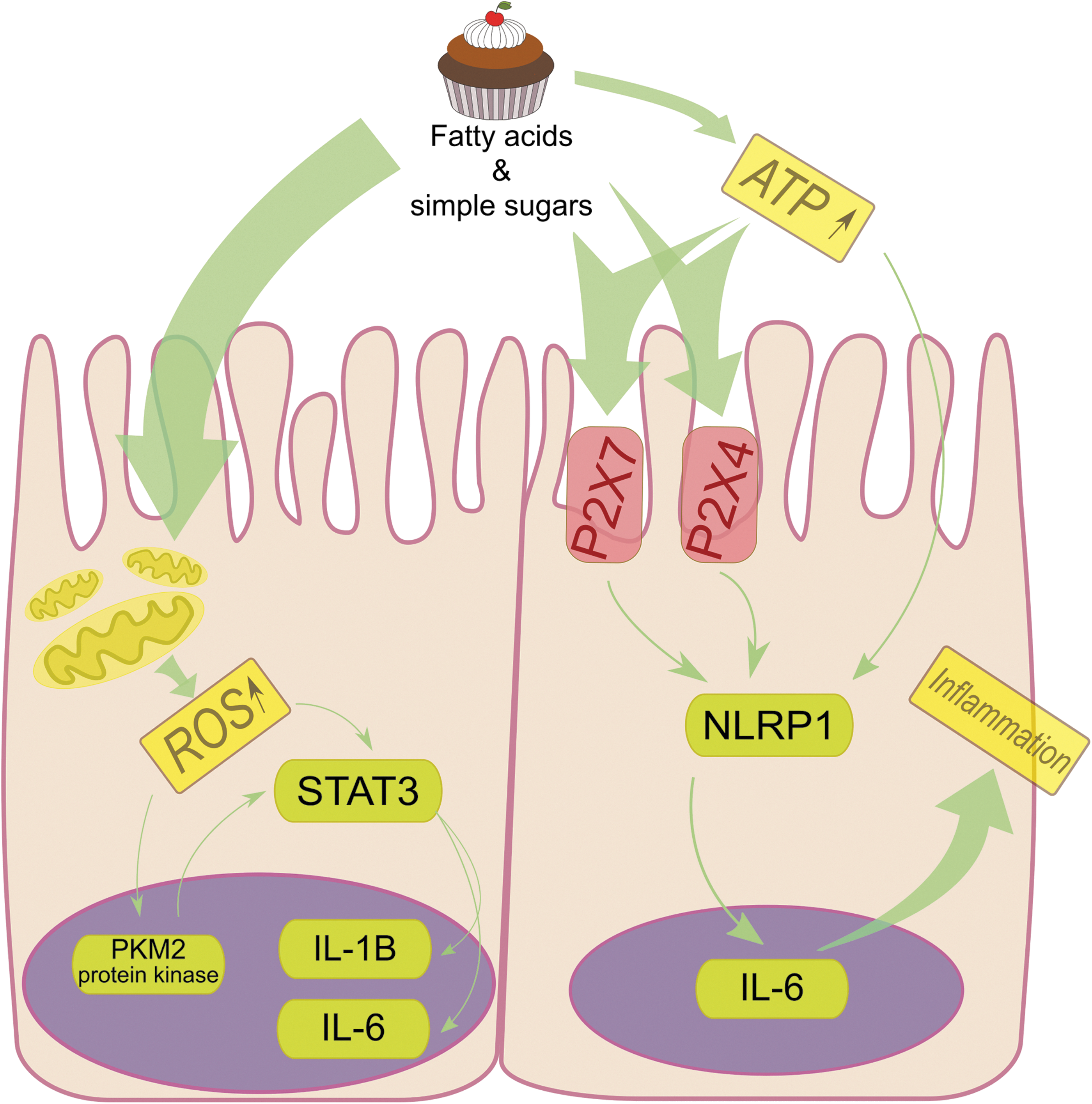
Long-term consumption of considerable amounts of sweets causes regional hyperglycemia. Simple carbohydrates and fatty acids, abundant in that type of food, may induce inflammation. High-glucose concentration leads to overexpression and activation of P2X7 and P2X4 receptors in the endothelium of gastrointestinal tract through increases in extracellular adenosine triphosphate (ATP) levels. The interaction of ATP with these receptors activates the intracellular inflammasome NLRP1 complex, which in turn enhances the expression of proinflammatory cytokines. Excessive glucose affects mitochondria, which increase their glycolytic activity and in turn secrete ROS. ROS leads directly and through activation of nuclear PKM2 protein kinase to the phosphorylation of STAT3 transcription factor, thus enhancing production of IL6 and IL1β. ROS, reactive oxygen species.
Depressive Symptoms Associated with Proinflammation
Sleep disorders
Incorrect sleep habits are clearly associated with mood disorders and increase the risk of depression in later life (Paunio and others 2015). In the course of depression, sleep disturbances are significantly more frequent than among the general population (Castro and others 2013). Depressed patients may encounter difficulties at every stage of sleep, subsequently compensating for it by lying in bed, which result in increased drowsiness during the day.
Nap time is closely associated with immune reaction, and the concentrations of acute-phase proteins, such as IL-6 and C-reactive protein (CRP), are higher during longer sleep (Irwin and others 2016).
Among healthy subjects, elevated IL-6 concentration was negatively correlated with sleep quality and positively correlated with ineffective interrupted sleep (Hong and others 2005). Higher basic levels of IL-6 among patients treated with interferon were associated with an increased prevalence of depression, more severe disease course, and were positively correlated with worse sleep quality (Prather and others 2009). Chronic insomnia changes the circadian rhythm of IL-6 secretion, the elevated concentration of which can exacerbate the disease course, probably by influencing the hypothalamic–pituitary–adrenal axis (Vgontzas and others 2002).
Proinflammation, as a stressor of the immune system, stimulates CRH release by hypothalamus, which activates pituitary adrenocorticotropic hormone secretion that affects adrenal glands (Kim and others 2016). The rs4719714 IL-6 gene polymorphism was reported to be correlated with the occurrence of insomnia during sleep initiation (Gay and others 2015). When investigated on animal models, IL-6 concentration was dependent both on the circadian rhythm and the type of tissue—in this case, higher IL-6 levels were noted during sleep rather than wakefulness (Guan and others 2005). Enhanced IL-6 secretion at night was also reported in primary insomnia, where its high total concentration had a deleterious effect on sleep quality (Burgos and others 2006).
The suicide threat
Almost 50% of patients suffering from depression have suicidal thoughts, of whom more than 16% undertake a suicide attempt (Patten and others 2015). The risk of suicide in depression is clearly associated with the severity of depressive episode and can be a result of exacerbated disease symptoms or an additional psychological burden (Kerr and others 2013; Knorr and others 2016).
In postmortem examinations, elevated IL-6 mRNA levels were detected in the brains of teenagers who had committed suicide (Pandey and others 2012). Furthermore, IL-6 concentrations were increased in the cerebrospinal fluid of people who had undertaken a suicide attempt (Lindqvist and others 2009). Suicidal risk in the course of depression is a factor that indicates its exacerbation, and IL-6 seems to be an independent parameter that is positively correlated with acceleration of depressive symptoms. Previously described insomnia, frequently associated with elevated levels of proinflammatory cytokines, increases suicidal risk and exacerbates the amplitude of depressive symptoms (Bozzay and others 2016).
Interestingly, levels of certain metabolites of the kynurenine pathway were found to be changed in the cerebrospinal fluid of patients who had attempted to commit suicide (Bay-Richter and others 2015). They also had significantly higher concentrations of IL-6 and quinolinic acid (QUIN). Some of the proinflammatory cytokines, including IL-6, are currently under consideration as markers of suicide risk because their levels were markedly elevated among patients who had attempted to commit suicide when compared with depressed patients with no suicidal thoughts (Janelidze and others 2011).
Although not all studies confirm the association between IL-6 and increased suicidal risk, there is nonetheless a clear association between its levels and the severity of depressive symptoms (Kim and others 2008). Molecular influence of IL-6 on behavioral suicidal tendencies may be due to the enhanced NOX2 and ROS expression in the GABAergic system, which leads to increased activity of glutamate receptors (Schiavone and others 2016). These, in turn, further stimulated by higher concentrations of QUIN, result in a more severe disease course and nervous tissue damage (Steiner and others 2011).
Fatigue
Chronic fatigue in the course of MDD, as one of the residual symptoms, decreases the number of remissions, increases the incidence of recurrences, and lengthens their duration (Fava and others 2014). Moreover, fatigue makes depressive symptoms more severe, which in turn deteriorate the psychophysical functioning of the patients (Ferguson and others 2014). However, it needs to be remarked that this symptom occurs in the course of numerous other disorders, it is not specific to depression, and its definition remains under debate (Carney and others 2014; Chung and others 2014).
Among dialysis patients, fatigue was found to be associated with the severity of depression and elevated serum IL-6 levels (Bossola and others 2015). Nevertheless, its higher concentration could be caused by the somatic state of the patient and additional stress associated with the procedure of dialysis (Rohleder and others 2012). A polymorphism in the IL-6 gene was reported to negatively affect the clinical course of fatigue among patients with cancer (Miaskowski and others 2010). Although earlier studies refuted the association between the overexpression of proinflammatory cytokines and fatigue after infection (Vollmer-Conna and others 2007), recently elevated IL-6 concentration was found to be positively correlated with fatigue severity after viral infection (Garcia and others 2014).
Psychomotor retardation
Psychomotor retardation is one of the axial depressive symptoms (American Psychiatric Association 2013). Phenotype of the disease, including chronic fatigue and psychomotor retardation, limits the activity of patients. Physical exercise modifies the expression of cytokines (Wang and others 2014). Regular aerobic exercise silences the expression of proinflammatory cytokines, whereas it activates the immunosuppressive genes (Iyalomhe and others 2015). Lower serum concentrations of IL-6 and acute-phase proteins were also observed after cardiovascular exercises (Kohut and others 2006). Elevated serum levels of IL-6 and the soluble form of its receptor were found during other types of physical activity, such as strength and endurance training (Steensberg and others 2006; Robson-Ansley and others 2009).
However, it needs to be remarked that initially excess exercising leads to burning adipose tissue, which may secrete cytokines in the process. Physical exercise induces changes in the concentrations of zinc and copper ions (Beghelli and others 2011) and alters the activity of certain metalloproteinases (Kim and others 2015). The serum ratio of Cu/Zn ion concentration is considered to be a marker of inflammation due to its association with elevated IL6 levels and CRP (Malavolta and others 2010). Zinc deficiency was found not only to induce proinflammation in leukemic monocytes but also to decrease the methylation of IL6 gene promoter, which might lead to its altered expression (Carmen and others 2015).
What is more, physical exercise was found to have an additive influence to metformin on the secretion of IL10 and leptin by adipose tissue, thus increasing their serum concentrations (Jenkins and others 2012). During physical activity, the overexpression of the IL6 gene leads to an enhanced expression of uncoupling protein 1 (UCP-1), which regulates the metabolism of adipose tissue (Sanchez-Delgado and others 2015). Adipocytes affect the availability of free fatty acids, which among obese individuals, may be responsible for the activation of macrophages through interactions with TLR4 receptors, thus leading to overexpression of proinflammatory cytokines by inducing NFkB (Howe and others 2013; Raajendiran and others 2016).
Lack of physical exercise caused by symptoms of psychiatric disorders may affect the profile of secreted cytokines, skewing it toward proinflammation. Increased IL-6 levels were found in the course of grave neurodegenerative disorders, in the course of which there frequently occurs considerable motor function impairment, that is, bradykinesia (Brodacki and others 2008; Scalzo and others 2010).
Epidemiological data indicate that mood disorders occur more frequently among people with BMI equal to or exceeding 30 kg/m2 (Simon and others 2006). Obesity may be closely linked with depression, significantly increasing the risk of symptom occurrence during 5 years of follow-up (Roberts and others 2003). Adipose tissue, as an organ, is functionally connected with the immune system. Adipocytes, which form the greater part of the adipose organ, synthesize proinflammatory cytokines and are intermediaries in antigen presentation. Among patients with a pathological adipose tissue distribution, this organ has autoimmune properties and induces chronic inflammation (Majdoubi and others 2016).
IL-6 and Depressive Symptoms in Bipolar Disorder—Similarities and Differences
Symptoms of depressive episodes vary greatly and their correct interpretation is vital for stating the appropriate diagnosis and introducing treatment. The classification of mood disorders is complex and requires constant updates of the current state of knowledge (Benazzi 2002). Unipolar depression, in a simplified manner understood as MDD, differs from bipolar depression by a number of features, including disease severity, duration of symptoms, epidemiology, genetic susceptibility, and chronicity of episodes (Benazzi 2002; Goodwin and Jamison 2007).
Depressive symptoms in bipolar disorder (BD), depending on the type of condition, are characterized by more frequent relapses, earlier occurrence of symptoms, stronger irritability, less severe psychomotor retardation and interest loss, more prevalent coexistence of particular anxiety and personality disorders, and greater proneness to overuse of certain psychoactive substances (Hantouche and Akiskal 2005; Akiskal and Benazzi 2006b; Akiskal and others 2006a, 2006c).
Differentiating BD from MDD can be achieved not only through symptomatology but also through their immunophenotype.
Immunological causes of BD may involve both Th1 lymphocytes and macrophages, the proinflammatory activity of which can alter the polarization of microglial cells and lead to their subsequent dysfunction through influencing the reactivity of blood–brain barrier endothelial cells (Ascoli and others 2016). In BD, the immunophenotype of T lymphocytes showed a changeable expression of TLR receptors, the stimulation of which in monocytes changes the profile of synthesized cytokines when compared with the control group (Wieck and others 2016). The expression of proinflammatory cytokine genes was also elevated in peripheral blood lymphocytes among bipolar patients (Pandey and others 2015).
Moreover, IL6 in BD is also considered to be a marker of the emotional component as its decreased level seems to be correlated with euthymia and its higher concentration with the severity of depressive episodes (Goldstein and others 2015). Elevated IL-6 concentration during depressive phase of BD was found among men whose disease started with a manic episode (Becking and others 2013). Nonetheless, this association was not statistically significant (Becking and others 2013).
Changes in cytokine concentrations may be caused by varying activation of Tregs. The immunophenotype of Tregs seems to differ depending on the type of BD. In BD type I, characterized by alternate depressive and manic episodes, a reduction of Tregs was observed with accelerated polarization of immune response typical for Th1 lymphocytes among patients in remission (do Prado and others 2013). A different study reported a decreased number of Tregs with the same IL6 mRNA expression among BD patients and control group (Drexhage and others 2011).
Chronic stress as an indicator of the emotional component of affective disorders may modify the plasticity of hypothalamic–pituitary–adrenal axis through activation of the JAK/STAT3 signaling pathway, which is regulated, among others, by IL-6 (Girotti and others 2013). Elevated IL6 concentration in childhood seems to be a risk factor for occurrence of hypomanic symptoms in the adult life (Hayes and others 2017). Furthermore, during manic episodes in the subtype of BD with rapid cycling, a higher level of this cytokine was noted (Munkholm and others 2015).
When it comes to BD, IL6 is one of the proinflammatory cytokines that regulates the production of kynurenic acid. The influence of IL6 on tryptophan metabolism may induce cognitive deficits, such as impaired memory, attention, and executive functions (Erhardt and others 2017). In fibroblasts of BD type I patients, the activity of the kynurenine pathway was enhanced, and IL6 affected the concentration of 3-hydroxykynurenine, but not that of kynurenic acid (Johansson and others 2013). The reason why there are conflicting results when it comes to the activation of immune system and elevated IL-6 levels, and depressive episodes in MDD and BD, might be caused by their heterogeneous symptoms, which in turn affect the concentration of this cytokine.
The severity of symptoms in BD, assessed using the Hamilton Depression Rating Scale and YMRS (Young Mania Rating Scale), was found to be correlated with IL-6 concentration and CRP and TNF-α levels, respectively (Lee and others 2013). These results suggest that there is an association between the emotional component and immune system (Lee and others 2013).
Investigating the role of cytokines in depression is difficult due to its chronicity, which requires long-term pharmacological treatment not only during exacerbation but also in remission (Tsai and others 2014), which in turn affects its immunophenotype and makes it difficult to assess it objectively. Introducing long-term pharmacological treatment induces iatrogenic changes in cytokine concentrations.
In case of BD pharmacotherapy, treatment of choice includes normothymic agents, which are mood stabilizers. The applications of classical drugs, such as lithium, valproic acid, carbamazepine, and lamotrigine, are limited by age of the patient and clinical phenotype of disease (Goodwin and others 2008; Himmerich and others 2013). Monotherapy with any of these drugs affects the profile of secreted cytokines, including IL-6 (Himmerich and others 2014). Treatment with lithium tends to increase the concentration of IL-6 (Nassar and Azab 2014; Boyko and others 2015).
Antidepressants used, for example, in MDD, directly affect the immune system and immune response. Chronic treatment with tricyclic drugs results in immunosuppression, with a decreased number of T lymphocytes and inhibition of NK cells (Xiao and Eneroth 1996; Komori and others 2002), whereas other groups have different impacts on the immune system (Taler and others 2007; Eller and others 2008; Li and others 2013).
The significance of IL-6 as a parameter of immune response in neuroimaging of depression
Due to numerous reports concerning the role of general inflammation in pathogenesis of psychiatric and neurodegenerative disorders, certain cytokines are currently investigated as potential markers of structural and functional brain changes (Harrison and others 2015). Particular attention is paid to IL-6, the level of which was correlated with a decreased total brain volume (Jefferson and others 2007). Interestingly, its level was also correlated with a decreased volume of hippocampal gray matter, which in turn resulted in a greater prevalence of limited cognitive functions among the elderly (Metti and others 2015).
Aging was found to be an important factor that affected the concentration of IL-6 and subsequent structural and functional brain changes. Cognitive functions and motor skills among healthy elderly seem to be correlated with volumes of particular brain regions, and minor degenerative changes can subsequently cause structural differences, thus explaining the cognitive and motor impairment (Koppelmans and others 2016). Systemic increase of IL-6 concentration and other proinflammatory cytokines may be explained by age-dependent loss of muscle mass and growth of adipose tissue, which alters the profile of immune response (Lutz and Quinn 2012).
Adipocytes as the main component of adipose tissue take part in cytokine synthesis and mediate in antigen presentation, and among people with visceral obesity, they exert autoimmune properties and induce chronic inflammation, including enhanced IL-6 synthesis (Majdoubi and others 2016). With age, the receptors of adipocytes also change, which affect not only their metabolism but also systemic glucose tolerance and the influence of insulin on cellular signaling (Mueller and others 2016).
The association between IL-6 and structural brain changes as a result of an aging immune system does not seem to fully elucidate the significance of this cytokine. Among young women, IL-6 turned out to be a significant factor of response to stress and its elevated levels were positively correlated with functional changes in dorsomedial prefrontal cortex and lower social status in functional magnetic resonance imaging (Muscatell and others 2016). The correlation between IL-6 concentration and the volume of hippocampal gray matter was confirmed by the study of Marsland and others (2015) and it was independent of age and body fat. Impaired cognitive functions, such as worsened short-term memory, spatial reasoning, and verbal communication, are common in depression and are currently considered to be the effect of general inflammation, and their correlation with elevated IL-6 concentrations was dependent on changes in volume of gray and white matter (Marsland and others 2015). The correlation between IL-6 and changes in hippocampal volume was described in MDD, and its joint occurrence with lower mRNA expression of particular genes associated with glucocorticoids suggests a potential influence of this cytokine on modulating the cognitive and emotional components under acute stress (Frodl and others 2012).
Conclusions
Currently, there are no known markers with clinical and diagnostic applications in MDD (Olajossy and others 2014; Buoli and others 2016). The potential influence of the immune system on pathogenesis of psychiatric diseases seems to be promising, but requires further more detailed studies. IL-6 as a potential marker of MDD is a cytokine with multiple properties. Both its synthesis and biological activity depend on a variety of factors. Clinical heterogeneity of depressive episodes can alter cytokine expression, including IL-6. The treatment and somatic state of the patient also play a role in regulating the immune response. Taking the abovementioned factors into consideration in future studies may help to elucidate the influence and significance of cytokines in the pathophysiology of depression.
Footnotes
Author Disclosure Statement
No competing financial interests exist.