
Abstract
Despite new treatments for hepatitis C virus (HCV) infection, IFNα-based regimens still have clinical relevance in special populations of patients and remain the only therapeutic option for many patients. We sought to elucidate the interplay between two relevant factors (IL28B polymorphism and T cell immune responses) involved in the outcome of this therapy in HCV-infected patients. We evaluated 38 patients infected with HCV genotype 1—17 coinfected with HIV—who were undergoing a full course of pegIFNα/RBV therapy. The interdependence and roles of T cell-mediated immune responses and IL28B rs12979860 single-nucleotide polymorphism genotype as predictors of virological response to anti-HCV treatment in patients with chronic hepatitis C were evaluated using nonparametric tests. Factors associated with rapid virological response (RVR) in univariate analysis were presence of CD4 T cell response against NS3 HCV protein, low baseline HCV-RNA, and IL28B CC genotype. Factors associated with sustained virological response (SVR) in univariate analysis were IL28B CC genotype, low baseline HCV-RNA, and presence of CD4 response against NS2. In the multivariate analysis, low baseline HCV-RNA and NS3-specific CD4 response showed a clear trend toward association with RVR (P = 0.09 and P = 0.07, respectively). Regarding SVR, IL28B CC genotype was the strongest predictor (P = 0.02), with presence of NS2-specific CD4 response showing a clear trend (P = 0.09). HCV-specific T cell response influences the outcome of pegIFNα/RBV therapy regardless of IL28B genotype. HCV-specific T cell responses (adaptive immunity) seem to influence viral clearance both in the short and long term during therapy (RVR and SVR), whereas the influence of the IL28B genotype (innate immunity) may be more relevant to the long-lasting therapeutic effect (SVR).
Introduction
N
Treatment with pegylated IFNα and ribavirin (pegIFNα/RBV) has been shown to have limited effectiveness and serious side effects (Fried and others 2002). These side effects and suboptimal effectiveness are exacerbated in patients coinfected with HIV, who make up a difficult-to-treat population and one with higher rates of chronicity and faster liver disease progression (Hernandez and Sherman 2011). Therefore, despite its limitations, there is still much interest in identifying aspects that allow for a better management of patients treated with IFNα-based regimens. In addition, research on genetics and immune mechanisms and the virus–host interaction in the context of this therapy could shed light on correlates of immune protection, which are quite necessary for the development of an effective HCV vaccine. This search, however, has proven to be much more complex than expected (Fauvelle and others 2013).
To date, several host and viral factors have been described as predicting virological response to anti-HCV therapy with IFN-based regimens (Shirakawa and others 2008; Masarone and Persico 2011). Given the immunomodulatory effects of pegIFNα and ribavirin (Brinkmann and others 1993; Thomas and others 2003; Langhans and others 2012) and the role of HCV-specific cellular immune response in viral clearance among patients able to eradicate the virus after acute HCV infection (Brinkmann and others 1993), HCV-specific immune responses have been suggested as a potential factor influencing IFNα-based treatment outcome, although with controversial results (Cramp and others 2000; Thimme and others 2001; Aberle and others 2007; Capa and others 2007; Pilli and others 2007; Rosen and others 2007; Burton and others 2008; Caetano and others 2008; Arends and others 2010; Moller and others 2011; Pembroke and others 2012; Zhang and others 2012).
As a result of this, there is no conclusive evidence for the role of HCV-specific T cell responses in treatment-induced viral eradication, and this may be due to different reasons. Most studies analyzing the potential association between virus-specific T cell responses and virological response to treatment did not control for other important baseline factors predicting response, such as the single-nucleotide polymorphism (SNP) located near the gene coding for IL28B or interferonλ3 (IFNλ3). This SNP is a host factor that is strongly associated with both sustained virological response (SVR) and on-treatment viral kinetics (Ge and others 2009; Tanaka and others 2009; Rallón and others 2010).
IFNλ3 belongs to the family of type III interferons, shares functional characteristics with type I interferons (IFNα and IFNβ), and has antiviral properties against HCV (Kotenko and others 2003; Robek and others 2005). A recent study has shown that plasma levels of IFNλ3 upregulate following administration of exogenous IFNα as part of anti-HCV therapy and that this upregulation is influenced by IL28B genotype, suggesting a tight in vivo relationship between the two interferons (Rallón and others 2012). On the contrary, in response to viral infections, the IFNλ family seems to upregulate the production of IFNγ (Ank and others 2006), a cytokine with an important role in lymphocyte recruitment into the liver tissue (Lalor and others 2002), and with a central role in regulating adaptive T cell responses (Schoenborn and Wilson 2007).
On the basis of this knowledge, it is important to clarify the relative role of HCV-specific T cell responses and of the IL28B genotype (as a factor related to antiviral innate response) in the context of HCV clearance induced by IFNα-based anti-HCV treatment. The aim of this study was to analyze the interdependence and relative roles of T cell-mediated immune responses and IL28B rs12979860 SNP genotype as predictors of virological response to anti-HCV treatment in patients with chronic hepatitis C.
Materials and Methods
Study population
A longitudinal study was conducted in 38 patients chronically infected with HCV genotype 1 (HCV-1), including 21 HCV-monoinfected (HCV group) and 17 HCV/HIV-coinfected (HCV/HIV group) patients. Twenty HCV- and HIV-seronegative healthy volunteers were included as controls to establish reference values for a positive HCV-specific T cell response as previously described (Rallón and others 2011a). All patients had completed a full course of therapy with pegIFNα-RBV, had a validated treatment outcome, and had stored cell samples available for IL28B rs12979860 SNP genotyping as previously described (Rallón and others 2011b) and for HCV-specific T cell response analysis. To participate in the study, written informed consent for specific examinations was obtained from all individuals, and the study protocol was evaluated and approved by the hospital ethics committee.
The following on-treatment virological responses were assessed: rapid virological response (RVR), defined as undetectable plasma HCV-RNA at week 4 after initiation of therapy; early virological response (EVR), defined as a decrease in HCV-RNA level of ≥2 log at week 12; end-of-treatment virological response (EOTVR), defined as undetectable plasma HCV-RNA at the completion of therapy; and SVR, defined as undetectable plasma HCV-RNA 24 weeks after the end of treatment. Patients with poor drug compliance and/or who discontinued therapy due to side effects were excluded from this analysis, as previously described (Rallón and others 2011b).
Viral load and HCV genotyping
Plasma HCV-RNA was measured using a real-time PCR assay (COBAS TaqMan 48; Roche, Barcelona, Spain), which has a lower limit of detection of 10 IU/mL. HCV genotyping was performed on plasma using a commercial RT-PCR hybridization assay (Versant HCV Genotype 2.0 LiPA; Siemens, Barcelona, Spain). Plasma HIV-RNA was measured using Versant HIV-1 RNA v 3.0 (Siemens), which has a lower limit of detection of 50 HIV-RNA copies/mL.
Clinical specimens
All immunological studies were done retrospectively using baseline (before initiation of anti-HCV therapy) stored cryopreserved peripheral blood mononuclear cells (PBMCs) samples. Viability of thawed PBMCs was always >85%.
IL28B genotyping (rs12979860 SNP)
The characterization of IL28B allelic variants was conducted in a blinded manner on DNA specimens collected from each individual. The SNP rs12979860 was assessed with a custom TaqMan assay (Livak 1999) using the 5′ nuclease assay with allele-specific TaqMan probes (ABI TaqMan allelic discrimination kit and the ABI7900HT Sequence Detection System; Applied Biosystems, Carlsbad, CA).
Overlapping HCV peptides
A genotype-matched peptide array of 460 overlapping peptides (Biodefense and Emerging Infections Research Resources Repository, Manassas, VA) spanning all HCV proteins was used to assess HCV-specific CD4+ and CD8+ T cell immune responses as previously described (Rallón and others 2011a). Each peptide was 12 to 19 amino acids (aa) long and overlapped the adjacent peptides by 11 to 12 aa. Overlapping peptides have been previously shown to be suitable for the measurement of both CD4+ and CD8+ T cell responses (Maecker and others 2002). In the stimulation assays, peptides were grouped into eight pools according to HCV protein of origin: core (28 peptides), E1 (29 peptides), E2/p7 (64 peptides), NS2 (32 peptides), NS3 (98 peptides), NS4A and NS4B (together 47 peptides), NS5A (71 peptides), and NS5B (91 peptides) as previously described (Rallón and others 2011a). The dimethylsulfoxide concentration in culture medium was <5% for all peptide pools.
Measurement of HCV-specific T cell responses
HCV-specific CD4+ and CD8+ T cell responses were evaluated by measuring the production of two different cytokines (IFNγ and TNFα) in response to HCV genotype-matched overlapping peptide pools, using multiparameter flow cytometry as previously reported (Rallón and others 2011a). For each sample, a minimum of 50,000 CD4+ and 50,000 CD8+ events were acquired using an FC500 flow cytometer, and data analysis was performed using CXP software (Beckman Coulter). Gating was done on CD8+ bright and CD4+ T cells. The percentage of CD8+ bright or CD4+ T cells expressing any combination of cytokines was assessed using the negative control of each sample to establish quadrant boundaries, as previously described (Rallón and others 2011a). Figure 1 shows a representative example of flow cytometry data with the gating strategy used.
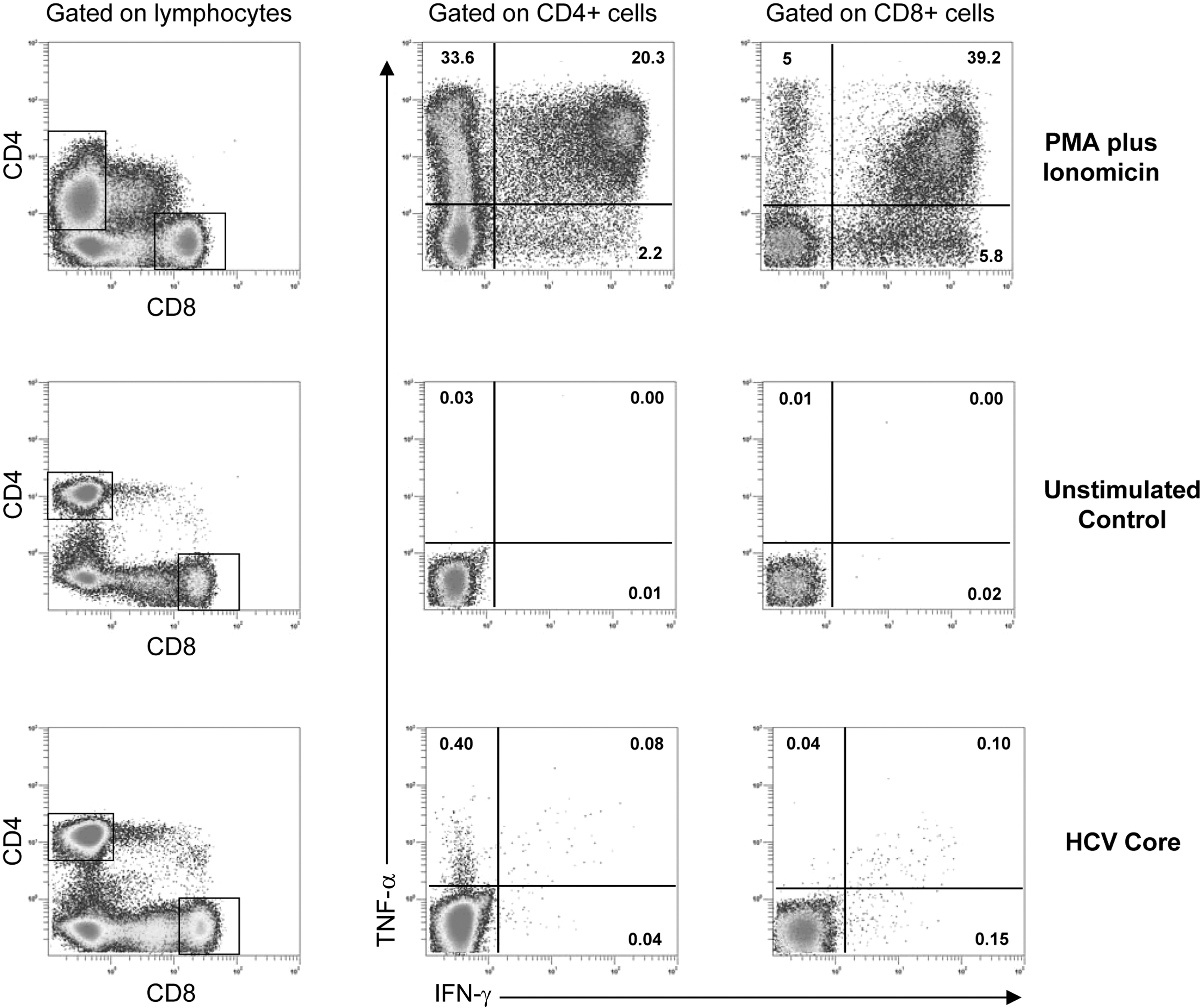
Representative example of flow cytometry data. Patterns of CD4+ and CD8+ T cell production of IFNγ and TNFα in response to polyclonal stimulation (PMA plus ionomycin), CD28 and CD49d stimulation (unstimulated control), and HCV core peptides (HCV core). Numbers in plots represent the percentage of positive events. HCV, hepatitis C virus; PMA, phorbol 12-myristate 13-acetate.
On the basis of expression of the two cytokines examined, three unique CD4+ and CD8+ T cell subsets were defined (cells not producing either of the two cytokines were not considered). For each of these subsets, a threshold for a positive response was defined as the 95 percentile of 160 pooled values obtained from eight different stimulations (with the eight HCV pools) in 20 HCV- and HIV-seronegative healthy donors, as previously described (Rallón and others 2011a). This threshold was applied to each sample after background subtraction (the response of the negative control in each particular sample). Prevalence of response for each HCV protein was defined as the proportion of patients showing detectable (above the threshold level) response for that particular protein. Level of response was defined as the proportion of T cells producing IFNγ and/or TNFα in response to each of the HCV proteins.
Statistical analyses
The main characteristics of the study population and the different parameters evaluated are expressed as median (interquartile range, IQR). Differences between groups were tested using nonparametric tests. Logistic regression analysis was applied to test the influence of T cell responses and IL28B genotype on treatment outcomes adjusting for baseline HCV viremia and presence of HIV coinfection. All statistical analyses were performed using the SPSS software version 13 (SPSS, Inc., Chicago, IL). All P values were two tailed and were considered as significant only when below 0.05.
Results
Study population
Table 1 shows the baseline characteristics of the two groups of patients included in the study. There were no significant differences between HCV and HCV/HIV groups in terms of age, baseline HCV-RNA, or IL28B genotype distribution. There were significantly more males in the HCV/HIV than the HCV group. As expected, the level (expressed as proportion) of CD4 cells was significantly higher in the HCV compared to the HCV/HIV group. Median (IQR) CD4 count in HCV/HIV patients was 442 (377–760) cells/μL. The majority (88%) of HCV/HIV patients were receiving highly active antiretroviral therapy and presented an undetectable level of HIV viremia at the moment of study.
HAART, highly active antiretroviral therapy; HCV, hepatitis C virus.
HCV-specific T cell immune response
In the whole population of patients, HCV-specific CD4 T cell response was detectable in 60%, while CD8 T cell response was detectable in only 37%. HCV core was the protein with the highest prevalence of CD4 response (43%), and HCV-E2/p7 proteins the most frequently targeted by CD8 T cells (prevalence of 24%). Level of response was very low overall, with median values below 0.5% in all cases; the global level of response (the sum of responses to the different proteins) was 0.39% (0.06–1.8) for CD4 and 0.50% (0.19–0.70) for CD8 response. Level of CD4 response for each of the HCV proteins tended to be higher compared to CD8 response, although the differences were not statistically significant (Fig. 2).
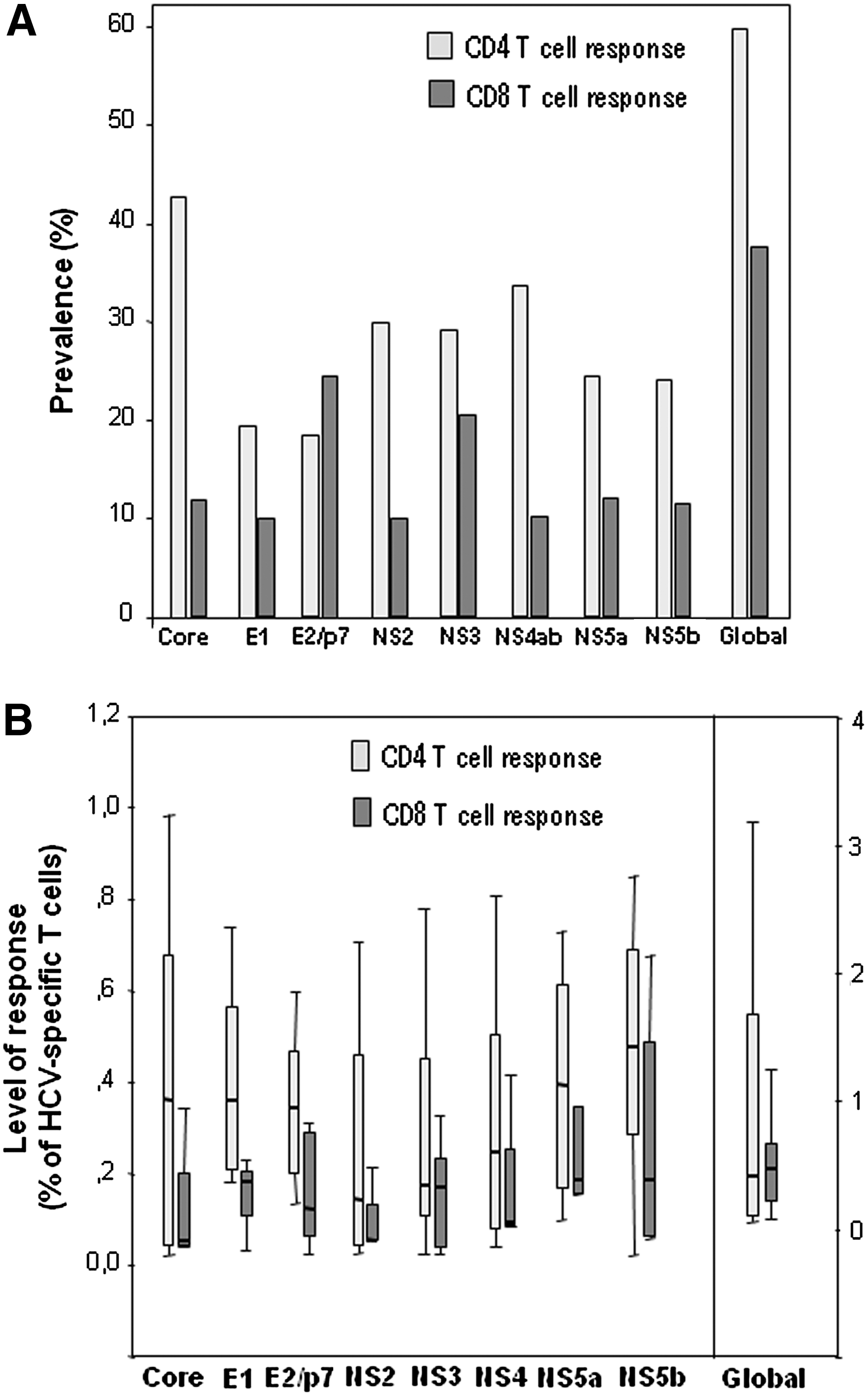
Prevalence
Prevalence of HCV-specific CD4 response tended to be higher in the HCV compared to the HCV/HIV group, although the differences observed did not reach statistical significance. However, prevalence of HCV-specific CD8 response to any of the proteins (global prevalence) was significantly higher in the HCV group when compared to the HCV/HIV group (62% versus 18%, P = 0.02). We observed a trend toward higher prevalence of HCV-specific CD8 response against E1, NS2, and NS5a proteins in the HCV compared to the HCV/HIV group, although the differences did not reach statistical significance.
It has been described that a higher rate of SVR after pegIFNα-RBV therapy occurs in patients carrying the rs12979860 IL28B CC genotype compared to patients carrying the non-CC genotype. Thus, we stratified our study population into CC and non-CC IL28B carriers and compared the prevalence of HCV-specific T cell response according to IL28B genotype. There were no significant differences in the prevalence of CD4 or CD8 T cell response between CC and non-CC patients (Fig. 3).

Prevalence of baseline HCV-specific CD4 and CD8 T cell response against the different HCV proteins in patients carrying IL28B CC or non-CC variants. “Global” indicates the presence of detectable response to any of the HCV proteins. HCV, hepatitis C virus.
Factors associated with virological response to anti-HCV treatment
In the whole population of patients, rates of RVR, EVR, EOTVR, and SVR were 14%, 78%, 66%, and 49%, respectively. The influence of HIV coinfection, IL28B genotype, and baseline HCV-RNA load on on-treatment virological responses is shown in Table 2. HCV/HIV-coinfected patients showed a significantly lower rate of EVR and EOTVR compared to HCV-monoinfected patients. IL28B CC carriers showed a significantly higher rate of EVR and SVR compared to non-CC carriers. RVR and EOTVR were also higher, although the difference did not reach statistical significance. Finally, patients with low baseline HCV-RNA load (<600,000 IU/mL) showed higher rates of RVR and SVR compared to patients with high baseline HCV-RNA load (≥600,000 IU/mL).
Low baseline HCV-RNA: <600,000 IU/mL. High baseline HCV-RNA: ≥600,000 IU/mL.
EOTVR, end-of-treatment virological response; EVR, early virological response; HCV, hepatitis C virus; RVR, rapid virological response; SVR, sustained virological response.
The association of HCV-specific T cell response with virological response was analyzed by stratifying patients according to the presence or absence of CD4 or CD8 T cell response against each of the HCV proteins. Figure 4 shows the rate of RVR and of SVR according to the presence or absence of HCV-specific CD4 response. Overall, the rate of RVR was higher in patients showing detectable HCV-specific CD4 response compared to those lacking this response. This difference reached statistical significance when patients were stratified according to CD4 response against NS2 protein (44% versus 0% of RVR in patients with and without detectable CD4 response, respectively, P = 0.006), and also when patients were stratified according to CD4 response against NS3 protein (36% versus 4% of RVR in patients with and without detectable CD4 response, respectively, P = 0.02). RVR showed a trend toward higher rates when CD4 response against NS4ab protein was present (P = 0.12). When patients were grouped according to the presence or absence of CD8 response, the rate of RVR tended to be higher in patients showing detectable CD8 response, although the differences were not statistically significant (data not shown).
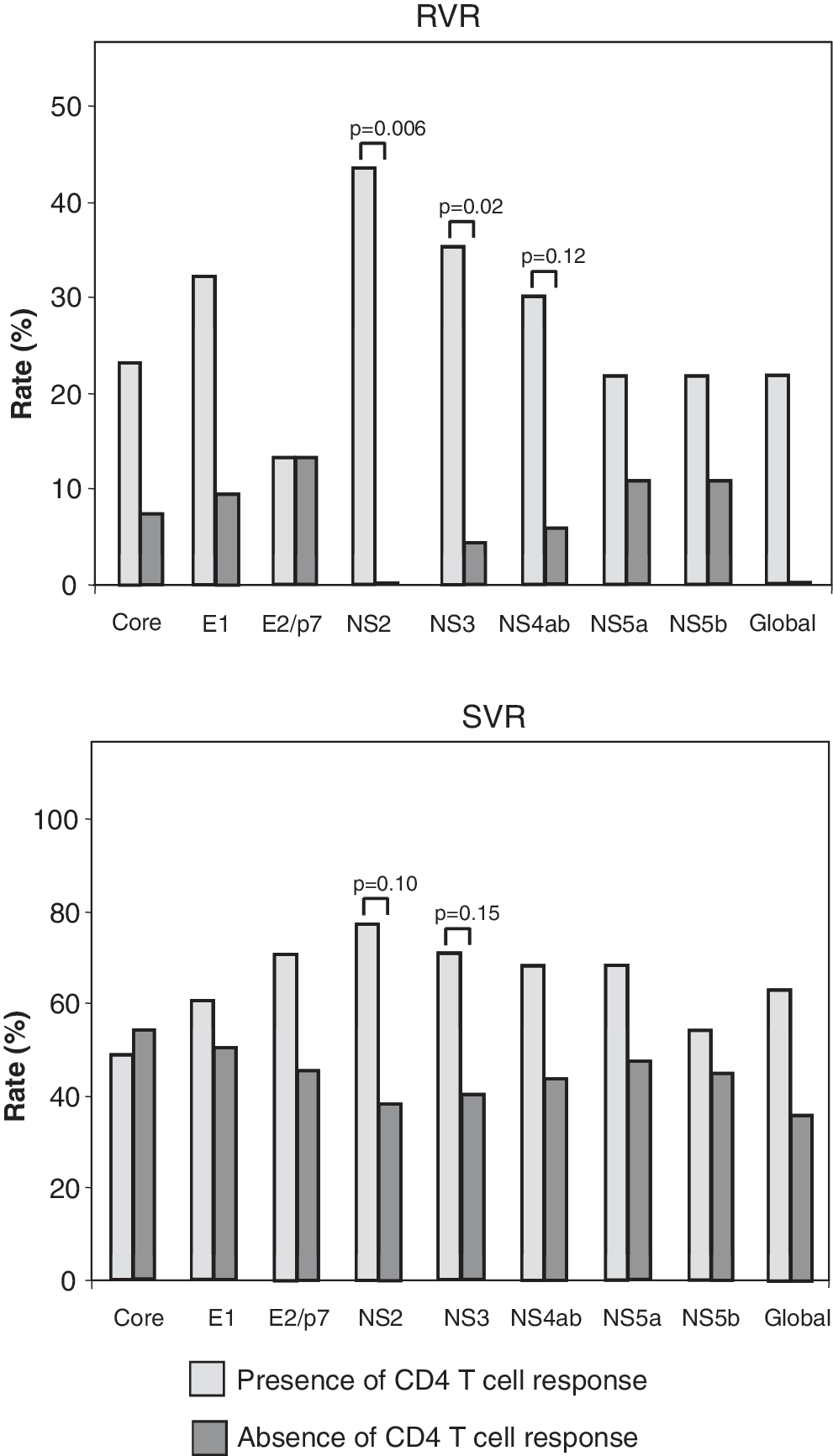
Rate of RVR and SVR according to the presence or absence of hepatitis C virus-specific CD4 T cell response. The term “global” refers to the presence of detectable response to any of the proteins. RVR, rapid virological response; SVR, sustained virological response.
Rates of EVR and EOTVR were similar between patients independently of the presence or absence of HCV-specific CD4 or CD8 response (data not shown). There were no differences in the rate of SVR according to the presence or absence of CD8 response. However, the rate of SVR tended to be higher in patients showing detectable CD4 response. Specifically, CD4 response against NS2 and NS3 proteins exhibited a clear trend (SVR: 78% versus 39%, P = 0.10 in patients with and without detectable CD4 response against NS2; SVR: 70% versus 40%, P = 0.15 in patients with and without detectable CD4 response against NS3) (Fig. 4).
A multivariate logistic regression analysis was performed to ascertain which factors were significantly and independently associated with the probability of attaining RVR (Table 3). Independent (explanatory) variables included in the analysis were IL28B genotype (CC versus non-CC), baseline HCV-RNA load (low versus high), and HCV NS3-specific CD4 response (presence versus absence). Factors showing a clear trend toward an association with RVR were (odds ratio [95% CI], P value) detectable CD4 response against HCV-NS3 protein (11.6 [0.85–160], P = 0.07) and low baseline HCV-RNA load (26 [0.64–1083], P = 0.09), whereas IL28B CC genotype did not show this same trend (P = 0.2). The same analysis applied to SVR and, including IL28B genotype (CC versus non-CC), baseline HCV-RNA load (low versus high), and HCV NS2-specific CD4 response (presence versus absence), revealed that IL28B CC genotype was the variable most strongly associated with SVR (15 [1.5–140], P = 0.02) followed by detectable CD4 response against HCV-NS2 protein, which showed a clear trend (7.3 [0.75–70], P = 0.09), whereas HCV load was not associated (P = 0.9).
CI, confidence interval; HCV, hepatitis C virus; OR, odds ratio; RVR, rapid virological response; SVR, sustained virological response.
Discussion
To our knowledge, this is the first study to analyze the interdependence and relative contribution of two different factors involved in the HCV eradication induced by pegIFNα/RBV treatment: one related to adaptive antiviral immune response and another related to innate antiviral immune response. The main findings of our study are as follows: first, IL28B genotype (as a factor related to innate immunity) is not associated with T cell mediated anti-HCV responses; second, both factors contribute significantly and independently to predictions of virological response after pegIFNα/RBV treatment; third, HCV-specific T cell responses (adaptive immunity) seem to influence viral clearance during the short (RVR) and the long (SVR) term of therapy, whereas the influence of IL28B genotype (innate immunity) may be more relevant in the long term (preliminary results were reported by Rallón and others 2015).
IL28B genotype is a strong predictor of treatment-induced HCV eradication (Ge and others 2009; Tanaka and others 2009; Rallón and others 2010), although the mechanisms underlying this association are not yet understood. A boosting of innate antiviral immunity through modulation of IFNλ3 production, either at baseline and/or after exogenous administration of IFNα, could be involved, although the results surrounding this issue are controversial (Langhans and others 2011; Rallón and others 2012).
Alternatively, IL28B genotype could influence the ability to mount an adaptive T cell-mediated anti-HCV response. In the simian model of AIDS, it has been shown that IFNλ3 increases CD4 response to vaccination with HIV DNA, as well as the killing activity of CD8 cells (Morrow and others 2009, 2010). These findings suggest that IL28B genetic variation could potentially impact cellular immune responses against viral pathogens. However, our study does not support this hypothesis since there was no correlation between HCV-specific T cell responses and IL28B genotype in HCV-infected patients. This finding of ours is in agreement with a very recent study in HCV spontaneous resolvers and in chronically infected HCV patients (Bes and others 2012). Nevertheless, a potential association of IL28B genotype with other characteristics of T cell responses not measured in our study cannot be ruled out.
On the contrary, our study is useful in elucidating the role of HCV-specific adaptive immunity in controlling HCV infection after treatment, which is an issue that has been widely debated over time, with some studies reporting an association (Cramp and others 2000; Sreenarasimhaiah and others 2003; Morishima and others 2006; Aberle and others 2007; Pilli and others 2007; Rosen and others 2007; Caetano and others 2008; Moller and others 2011; Zhang and others 2012) but not others (Lauer and others 2005; Capa and others 2007; Burton 2008; Barnes and others 2009; Arends and others 2010; Pembroke and others 2012). Several reasons may account for these contradictory results, including differences in study design (cross sectional or longitudinal), different patient cohorts (including different HCV genotypes or restricted to only one genotype), differences in ways of measuring treatment outcome (either early or long-term viral kinetics), and different methodological approaches to evaluate T cell responses.
We used a longitudinal design in a cohort of patients infected with HCV genotype 1 (the most difficult-to-treat genotype) and evaluated T cell responses using overlapping peptides that encompassed the whole HCV proteome. Moreover, our cohort of patients also included patients coinfected with HIV (an additional factor of poor response to treatment). Using this approach, we found that the presence of HCV-specific T cell responses at baseline is associated with achievement of virological response to pegIFNα/RBV treatment. Interestingly, this association was restricted to CD4 responses against nonstructural proteins, and was more apparent with viral clearance during the first weeks of therapy (RVR), although it was also present with outcome in the long term (SVR).
Our results regarding the association between T cell responses and early viral kinetics (RVR) are in agreement with two previous studies (Aberle and others 2007; Pilli and others 2007); indeed, this association was independent of other well-known predictors of response, such as the IL28B genotype, which is one of the most interesting and original findings from our study. Our finding strongly supports the role of virus-specific T cell response in viral kinetics during the first weeks of therapy, corresponding to the second phase of HCV viremia decline (Neumann and others 1998), which has been attributed to the death of infected hepatocytes driven by HCV-specific immune responses (Neumann and others 1998; Layden and Layden 2002), and is in agreement with the established role of this type of immune response in the spontaneous clearance of HCV during acute infection (Thimme and others 2001).
A potential mechanism underlying this association could be the boosting of T cell response (present before the starting of pegIFNα/RBV therapy) after the very first reduction of HCV viremia induced by therapy, and this enhanced T cell response may contribute to HCV elimination thereafter. This hypothesis is supported by previous studies showing that HCV-specific T cell responses are boosted in parallel with a rapid reduction of HCV during the first weeks of therapy (Kamal and others 2002; Aberle and others 2007). In fact, our results demonstrating a clear trend toward an association of RVR with T cell response at baseline (before the administration of treatment) provide empirical support of the role of T cell responses as the cause of HCV clearance during the first weeks of therapy.
On the contrary, IL28B CC genotype was the strongest predictor of SVR, suggesting a role played by innate immunity in long-term viral kinetics. Given the anti-HCV action of IFNλ3 (Kotenko and others 2003; Robek and others 2005), a modulated expression of this molecule driven by IL28B genetic variation (Rallón and others 2012) could be involved in this long-term antiviral effect. Further studies measuring the levels of IFNλ3 longitudinally during pegIFNα/RBV treatment are needed to test this hypothesis.
The main strengths of our study are the prolonged observation period in a population of patients being treated for HCV infection, lasting the entire period of follow-up, and the detailed analysis of immunological responses, IL28B polymorphism, and several on-treatment outcomes with clinical relevance. Moreover, we were able to test the contribution and interdependence of both HCV-specific T cell responses and IL28B genotype to anti-HCV therapy outcome.
However, some limitations of our study merit comment, beginning with the relatively small number of patients analyzed together with the fact that some other factors impacting on treatment outcome were not controlled in our study; these likely prevented most of the results from reaching a level of significance below 0.05. In addition, we did not measure other characteristics of T cell responses and thus cannot rule out the existence of other potential associations between T cell responses and virological response. However, it must be noted that even with these limitations, we found a clear trend toward an association between T cell responses and treatment outcome. These findings make the case for further studies with larger cohorts to validate our results.
In summary, our study is the first to explore the interdependence and roles of T cell-mediated immune responses and IL28B genotypes on several clinically relevant on-treatment outcomes as predictors of virological response to pegIFNα/RBV therapy in patients with chronic HCV infection. Our results suggest that HCV-specific T cell responses (adaptive immunity) influence viral clearance throughout HCV treatment (RVR and SVR) independently of IL28B genotype. Within this context, the influence of IL28B genotype (innate immunity) is more significant on the long-lasting effect of therapy (SVR).
Footnotes
Acknowledgments
The authors thank all the patients who participated in the study. This study was funded by projects RD12/0017/0031 and PI14/00518 (part of the Plan Nacional I+D+I) and was cofunded by ISCIII-Subdirección General de Evaluación and the European Regional Development Fund. Norma Rallón is a Miguel Servet investigator from the ISCIII (CP14/00198), Madrid, Spain. Marcial García is a predoctoral student cofunded by the CP14/00198 project and Intramural Research Scholarship from IIS-FJD.
Author Disclosure Statement
No competing financial interests exist.