
Abstract
Neutralizing antibodies may affect interferon (IFN)-β treatment efficacy, but mechanisms of neutralizing anti-drug antibody (ADA) evolution are not fully elucidated. We investigated the relationship between ADA titers, IgG subclass profile, and binding affinity with the development and persistency of neutralizing ADA in relapsing-remitting multiple sclerosis (MS) patients treated with subcutaneous IFN-β. A total of 94 patients, who had blood sampling at months 6, 12, 24, and 36 during IFN-β therapy, were included into this retrospective study and stratified to the following: non-neutralizing, transient, and persistent neutralizing ADA status. Patients without or with transient neutralizing ADA displayed predominantly IgG1 and IgG3 subclasses, lower ADA titers, and antibody binding affinity compared with patients having persistent neutralizing ADA, in whom the predominant IgG subclasses were IgG2 and IgG4. Overall, ADA binding affinity positively correlated with IgG4 and neutralizing ADA titers, but negatively with IgG3 titers. Persistency of neutralizing ADA was predicted by their titers at month 24 and month 36 of treatment and by an increase of antibody affinity within the second year of IFN-β treatment. The humoral immune response to IFN-β observed in MS patients as a result of IFN-β therapy is a multifactorial process that is influenced by ADA titers, affinity maturation, and IgG subclass switching.
Introduction
I
The reasons that neutralizing ADA develop and, if present, persist in some patients and revert in others are still not fully elucidated. It has been shown that the development of neutralizing ADA is preceded by the occurrence of non-neutralizing ADA (Perini and others 2004) and that (non-neutralizing) ADA titers are significantly higher in those patients who develop neutralizing ADA after extended treatment (Mayr and others 2003; Hegen and others 2014). Furthermore, in neutralizing ADA-positive patients, immunoglobulin (Ig) G subclass profile is different (Deisenhammer and others 2001; Gibbs and Oger 2007) and antibody binding affinity is significantly higher compared with neutralizing ADA-negative patients (Gneiss and others 2006a).
Persistency of neutralizing ADA correlates with neutralizing ADA titer (Gneiss and others 2004; Hegen and others 2012) and depends on the type of IFN-β (with higher reversion rates and earlier reversion to seronegativity in patients on IFN-β-1b as compared with those on IFN-β-1a treatment) (Herndon and others 2005; Sorensen and others 2005; Hegen and others 2012). Neutralizing ADA against IFN-β-1a shows later peak titers (Hegen and others 2012), probably explaining why some authors reported higher titers as compared with IFN-β-1b after at least 2 years of treatment (Gneiss and others 2006b).
Here, we hypothesize that ADA against IFN-β undergoes IgG subclass switch and affinity maturation in a titer-dependent manner in those patients who develop neutralizing ADA. As such, a relationship between these parameters has yet to be investigated within a single, large, and well-characterized patient cohort, so we performed this longitudinal study.
Materials and Methods
Patients and samples
Patients with relapsing-remitting (RR) MS who were on IFN-β treatment were included from 2 different sources: the pivotal IFN-β-1b trial (n = 57) (The IFNB Multiple Sclerosis Study Group and the University of British Columbia MS/MRI Analysis Group 1996) and the Austrian ADA registry (n = 37). Inclusion criteria were as follows: age of at least 18 years, a continuous subcutaneous (s.c.) IFN-β-1a or IFN-β-1b treatment over 3 years (without switches between different preparations), and the availability of serum samples at months (mo) 6, 12, 24, and 36 after the start of therapy.
Patients were divided into 4 groups according to their ADA status: (1) NEGB (non-neutralizing ADA against IFN-β-1b): Patients on IFN-β-1b tested ADA positive at least at mo12, but neutralizing ADA negative at any time point (source: Austrian ADA registry); (2) TRANSB (transient neutralizing ADA against IFN-β-1b): Patients on IFN-β-1b tested neutralizing ADA positive at mo12 and/or mo24, but neutralizing ADA negative at mo36 (source: pivotal IFN-β-1b trial); (3) PERB (persistent neutralizing ADA against IFN-β-1b): Patients on IFN-β-1b tested neutralizing ADA positive at mo24 and mo36 (source: pivotal IFN-β-1b trial); (4) PERA (persistent neutralizing ADA against IFN-β-1a): Patients on s.c. IFN-β-1a tested neutralizing ADA positive at mo24 and mo36 (source: Austrian ADA registry).
Terminology of ADA is based on the recommendations of the Innovative Medicines Initiative ABIRISK Consortium (Rup and others 2015). All blood samples were collected by peripheral venous puncture. Serum was isolated from blood by centrifugation; afterward, the blood samples were allowed to clot for ≥30 min and stored at −20°C until performing the assays.
The study was approved by the Ethics Committee of the Medical University of Innsbruck, Austria.
ADA assay
We measured ADA by a direct enzyme-linked immunosorbent assay (ELISA) as described earlier (Gneiss and others 2008) apart from the following modifications (to get more precise ADA titers): Samples initially tested to have ADA titer <1:50 were re-tested starting with a serum dilution of 1:10 (ie, ADA positivity was considered in case of a titer ≥1:10); samples with a screening titer ≥1:6,400 were re-tested by using serial dilutions up to 1:51,200.
Coefficients of variation (CV) of positive controls (calculated for log-transformed ADA titers) run in all assays ranged from 3.9% to 4.6%.
ADA IgG subclass assay
ADA IgG1–4 subclasses were determined by ELISA as previously reported (Deisenhammer and others 2001) apart from the following modifications: As detection antibody, we used a mouse anti-human biotin-conjugated IgG1–4 antibody (Invitrogen; diluted at 1:2,000 with 0.15% bovine serum albumin [BSA] in phosphate-buffered saline [PBS]), and we used horseradish peroxidase-conjugated streptavidin (diluted 1:10,000 in 3.0% BSA/PBS, incubation for 1 h). Samples were run at dilutions as for ADA assay. ADA IgG1–4 titers ≥1:10 were considered positive.
Only sera tested positive for ADA were used for determination of IgG1–4 subclasses; for ADA negative samples, titers of ADA IgG1–4 were considered negative (without testing).
CV of positive controls run in all assays was as follows: 0%–5.8% for ADA IgG1, 6.0% for IgG2, 8.2% for IgG3, and 4.5%–8.5% for IgG4.
ADA affinity assay
Affinity of ADA was determined by a previously described immunoassay, which returns relative affinity values (RAV) based on disrupting interactions between antibody and antigen using increasing concentrations of the chaotropic agent sodium thiocyanate (Gneiss and others 2006a). Here, we deviated from the original protocol (Gneiss and others 2006a) by using only IFN-β-1b for ELISA plate coating. Furthermore, for samples that did not reach a differential optical density (OD) of 0.7–1.3 in dilutions of ≥1:50 in the ADA assay, we used decreasing serum dilutions down to 1:10.
Neutralizing ADA assay
Neutralizing ADA titers of samples from the IFN-β-1b pivotal trial (groups TRANSB and PERB) were determined by a cytopathic effect (CPE) assay (results kindly provided by Bayer HealthCare, San Francisco, CA) (The IFNB Multiple Sclerosis Study Group and the University of British Columbia MS/MRI Analysis Group 1996). Neutralizing ADA titers of samples from the groups NEGB and PERA were measured by a cell-based, myxovirus protein A (MxA) induction assay as previously described (Millonig and others 2009). In a subset of samples (n = 11), which have been also tested by the CPE assay, we determined neutralizing ADA titers by the MxA induction assay and found a favorable concordance [Kendall's Correlation Coefficient (CC): 0.782; P = 0.001]. For both assays, neutralizing ADA titers ≥20 neutralizing units (NU) were considered positive.
Statistical analysis
We performed statistical analysis by using SPSS 22.0 (SPSS, Inc., Chicago, IL) and GraphPad Prism 6.01 (GraphPad, Inc., La Jolla, CA). Distribution of data was assessed by the Kolmogorov-Smirnov test. Variables that did not follow normal distribution were log-transformed for further analyses. For overall group comparisons, we used the Kruskal-Wallis test with Dunn's multiple comparison post-test (for ADA titers and IgG1–4 titers) or one-way ANOVA with Tukey's multiple-comparison post-test (for RAV). For 2 group comparisons (PERB versus TRANSB or PERA), we applied the Mann-Whitney test (for ADA titers and IgG1–4 titers) or Student's t-test (for RAV). Correlation between neutralizing ADA titers returned by CPE and MxA induction assay was assessed by Kendall's tau rank CC. All other correlations were expressed by Spearman correlation coefficient (R). To identify independent predictors for neutralizing ADA persistency, we performed a multivariate linear regression analysis [including only patients from the groups TRANSB and PERB (to avoid the influence of different IFN-β preparations) and including only variables that had been identified as significant predictors in the univariate regression analysis].
Results
A total of 94 patients with RRMS were included into this retrospective, longitudinal study. All patients were on IFN-β treatment during the observation period of the median of 17 months, that is, a median of 3 longitudinally collected samples was available per patient. Patients were classified according to their ADA status (see Materials and Methods section); numbers of patients per group as well as numbers of samples per visit are shown in Table 1.
for definition of patient groups, see Materials and Methods section.
Eight MIU IFN-β-1b s.c. e.o.d.
Fifteen patients received 8 MIU IFN-β-1b s.c. e.o.d, and 14 patients received 1.6 MIU IFN-β-1b s.c. e.o.d.
Twelve patients were on 8 MIU IFN-β-1b s.c. e.o.d, and 16 patients were on 1.6 MIU IFN-β-1b s.c. e.o.d.
Significantly higher than in TRANSB (Kruskal-Wallis test with Dunn's post test, adjusted P = 0.0075).
Significantly higher than in PERA (Mann-Whitney test, P = 0.002).
Significantly higher than in PERA (Mann-Whitney test, P = 0.0005) or TRANSB (Kruskal-Wallis test with Dunn's post test, adjusted P = 0.0052).
Significantly higher than in TRANSB (Kruskal-Wallis test with Dunn's post test, adjusted P = 0.0021).
Significantly higher than in TRANSB (Kruskal-Wallis test with Dunn's post test, adjusted P = 0.0157).
Significantly higher than in TRANSB (Mann-Whitney test, P = 0.0031).
Significantly higher than in TRANSB (Mann-Whitney test, P < 0.0001).
Sixteen patients received 22 μg IFN-β-1a s.c. t.i.w., and 10 patients received 44 μg IFN-β-1a s.c. t.i.w.
ADA, anti-drug antibodies; e.o.d, every other day; IFN-β, interferon beta; IQR, interquartile range; MIU, million international units; NU, neutralizing units; s.c., subcutaneous; t.i.w, 3 times in a week.
Longitudinal development of ADA titers
In the NEGB group, median ADA titers peaked at mo12 and decreased to a median titer of 0 at all later time points. In TRANSB and PERB groups, median ADA titers peaked at mo6 and decreased thereafter. Median ADA titers were negative in TRANSB at mo24 and mo36, whereas patients of the PERB group still showed positive median titers at these time points. ADA titers in PERB were significantly higher than in TRANSB at each visit. In the PERA group, median ADA titers remained elevated and similar at all time-points, but were significantly lower than in PERB at mo6 and mo12. All ADA titers (and neutralizing ADA titers) per patient group and visit are displayed in Table 1.
Longitudinal evolution of ADA IgG1–4 subclass titers
In the NEGB and TRANSB groups, median IgG1 titer returned to 0 at mo24 and mo36, although it remained high in patients with persistent neutralizing ADA (PERB and PERA groups). At mo6 and mo12, median IgG1 titer was significantly higher in PERB than in PERA (Fig. 1). Median IgG2 titers were negative in all patient groups at all visits except for the PERB group at mo6 and mo12 (Fig. 2). Median IgG3 titers were positive in all 4 groups at mo6 and reached negativity at mo24 and mo36 in the NEGB and TRANSB groups. In PERB, titers decreased over time but remained elevated until the last visit. Median IgG3 titers were significantly lower in PERA than in PERB at all study visits (Fig. 3). Median IgG4 titers were 0 in NEGB and TRANSB at all visits, but elevated in PERB and PERA at all visits except for PERA at mo6 and for PERB at mo36 (Fig. 4).

Longitudinal profile of ADA IgG1 subclass titers in different patient subgroups. Mo6: In PERB significantly higher than in PERA (Mann Whitney test, P = 0.0007), NEGB, or TRANSB (Kruskal-Wallis test with Dunn's post test, adjusted P = 0.0063 and P = 0.001, respectively); Mo12: In PERB, significantly higher than in PERA (Mann Whitney test, P < 0.0001) or TRANSB (Kruskal-Wallis test with Dunn's post test, adjusted P = 0.0017); Mo24: In PERB, significantly higher than in NEGB or TRANSB (Kruskal-Wallis test with Dunn's post test, adjusted P = 0.0319 and P = 0.0012, respectively). Mo, month. For definition of patient groups (NEGB, TRANSB, PERB, PERA), see Materials and Methods section. Only statistically significant differences between groups are indicated. ADA, anti-drug antibody.
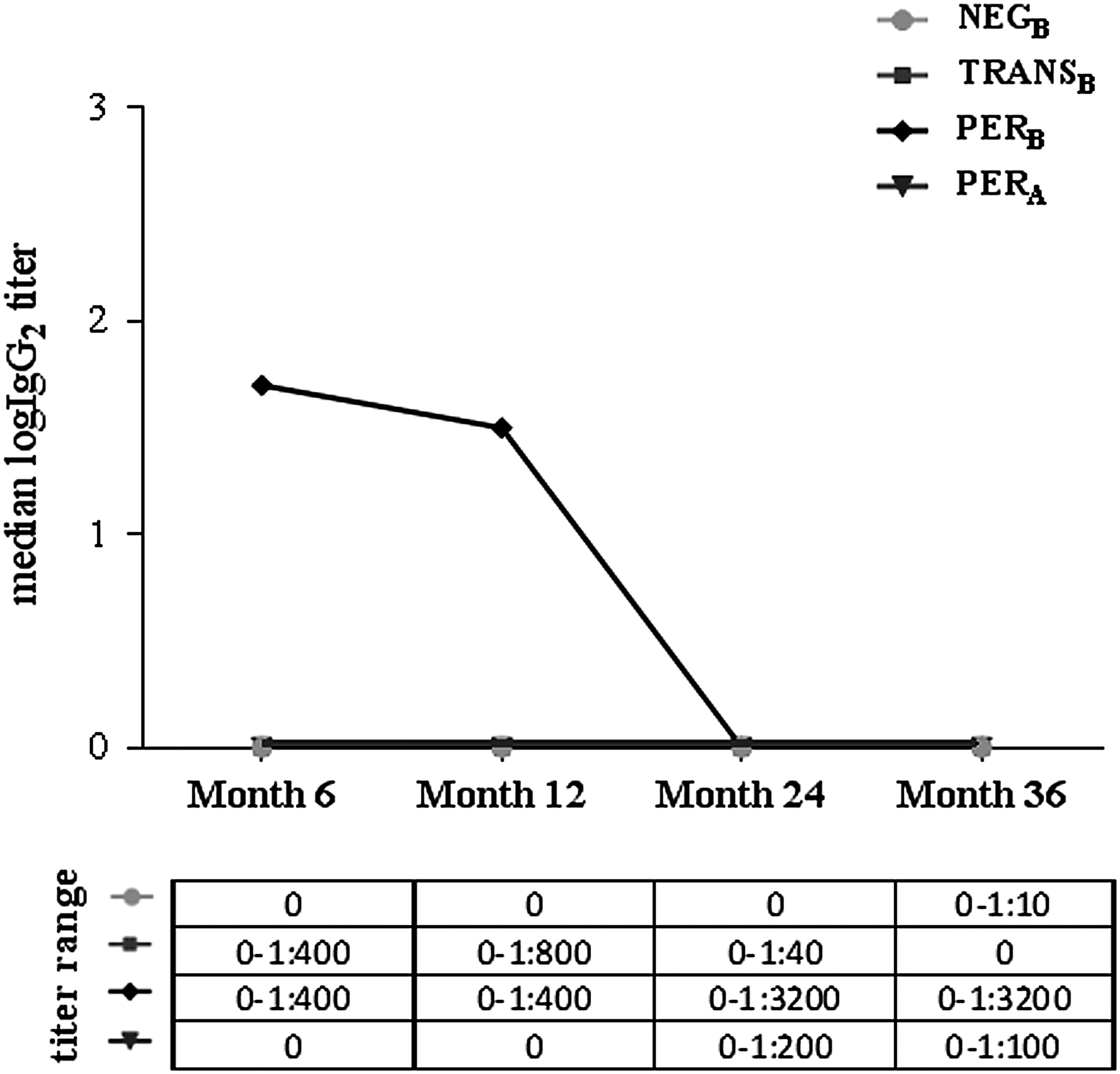
Longitudinal profile of ADA IgG2 subclass titers in different patient subgroups. Mo12: In PERB, significantly higher than in PERA (Mann Whitney test, P = 0.0013), NEGB, or TRANSB (Kruskal-Wallis test with Dunn's post test, adjusted P = 0.0186 and P = 0.0291, respectively); Mo24: In PERB, significantly higher than in TRANSB (Kruskal-Wallis test with Dunn's post test, adjusted P = 0.0145). Mo, month. For definition of patient groups (NEGB, TRANSB, PERB, PERA), see Materials and Methods section. Only statistically significant differences between groups are indicated.
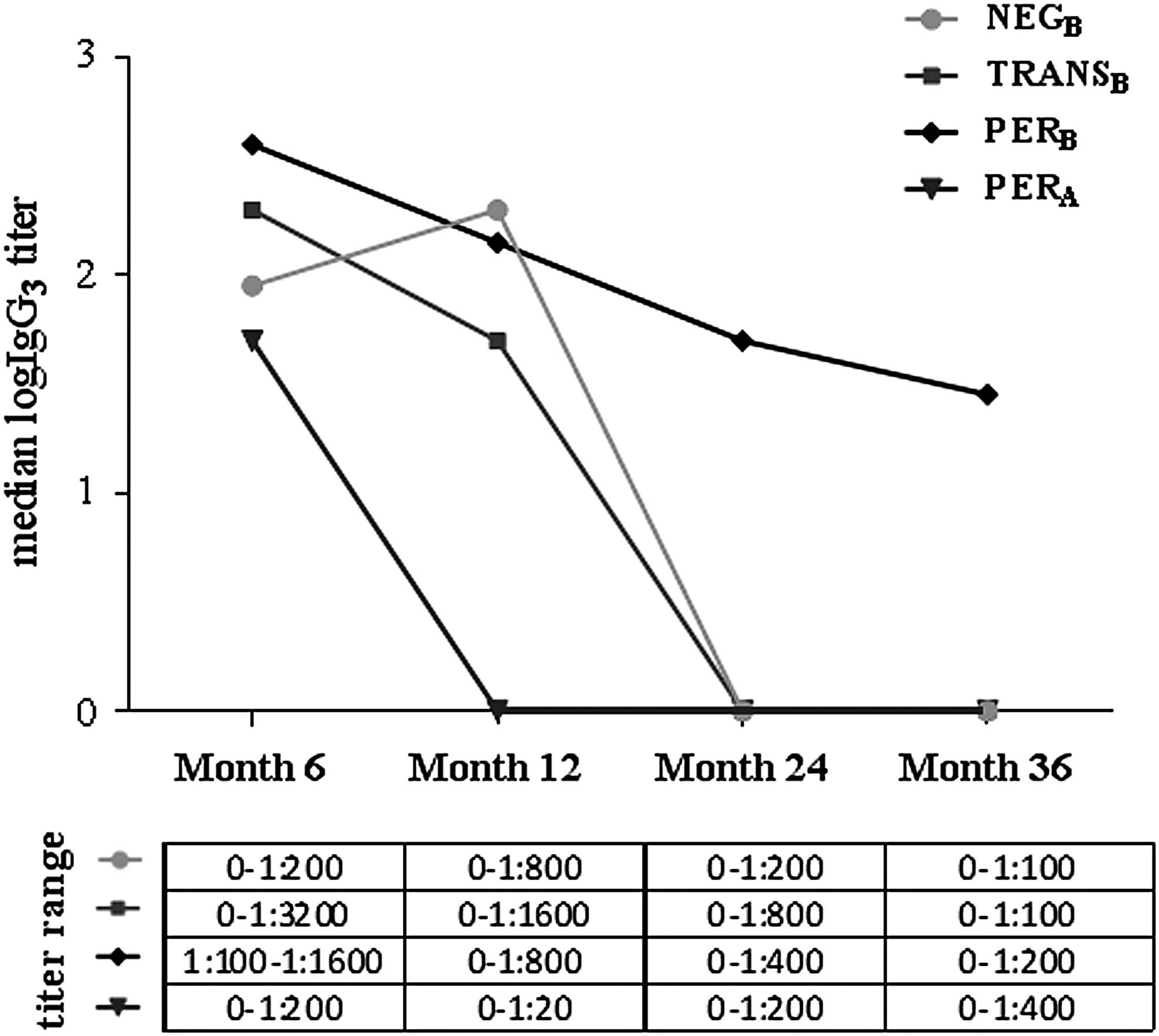
Longitudinal profile of ADA IgG3 subclass titers in different patient subgroups. Mo6: In PERB, significantly higher than in PERA (Mann Whitney test, P = 0.0129); Mo12: In PERB, significantly higher than in PERA (Mann Whitney test, P < 0.0001); Mo24: In PERB, significantly higher than in PERA (Mann Whitney test, P = 0.0082); Mo36: In PERB, significantly higher than in PERA (Mann Whitney test, P = 0.0459). Mo, month. For definition of patient groups (NEGB, TRANSB, PERB, PERA), see Materials and Methods section. Only statistically significant differences between groups are indicated.

Longitudinal profile of ADA IgG4 subclass titers in different patient subgroups. Mo12: In PERB, significantly higher than in TRANSB (Kruskal-Wallis test with Dunn's post test, adjusted P = 0.0429); Mo24: In PERB, significantly higher than in TRANSB (Kruskal-Wallis test with Dunn's post test, adjusted P < 0.0001). Mo, month. For definition of patient groups (NEGB, TRANSB, PERB, PERA), see Materials and Methods section. Only statistically significant differences between groups are indicated.
Longitudinal change of ADA binding affinity
In the NEGB group, mean RAV peaked at mo24 and was significantly lower than in both TRANSB and PERB at mo6, mo12 and significantly lower than in PERB at mo36 (Fig. 5). In TRANSB, mean RAV reached its peak value at mo12, decreased thereafter, and showed significantly lower values than in PERB at mo24 and mo36. In the PERB group, mean RAV gradually increased over 2 years of IFN-β treatment and remained high at mo36; at this visit, mean RAV was significantly higher as compared with both, NEGB and TRANSB. Mean RAV in PERB and PERA was similar at all study visits except at mo6 (Fig. 5).
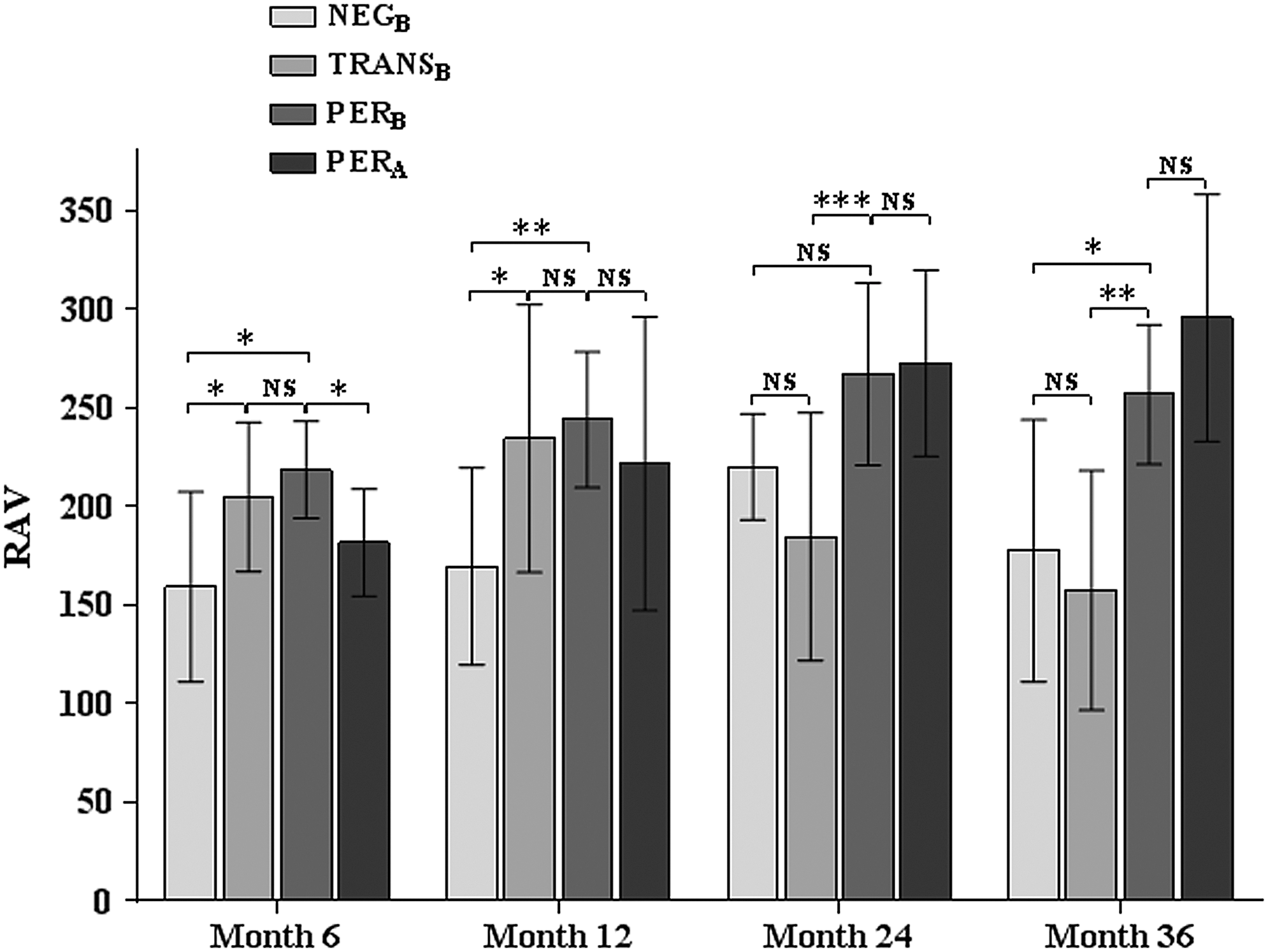
Longitudinal changes of ADA binding affinity in different patient subgroups. NEGB, TRANSB, and PERB were compared by using ANOVA with Tukey's post test, and PERB and PERA were compared by using Student's t-test; NS, non-significant; *, **, *** indicate a statistical significance at a P value <0.05, <0.01 and <0.001; data are shown as mean ± standard deviation. For a definition of patient groups (NEGB, TRANSB, PERB, PERA), see Materials and Methods section.
Correlations between ADA titers, neutralizing ADA titers, ADA IgG1–4 subclass titers, and ADA binding affinity
Overall, there was a significant correlation of ADA titers with all IgG subclass titers (eg, IgG1 R = 0.92, IgG2 R = 0.53, IgG3 R = 0.69, IgG4 R = 0.52; all P values <0.0001). Overall, neutralizing ADA titers correlated significantly with ADA titers in general and titers of the IgG subclasses 1, 2, and 4 (Table 2). A correlation of RAV was observed with neutralizing ADA titers, but not ADA titer. There was a significant positive correlation of RAV with the IgG4 subclass, and a negative correlation with the IgG3 subclass titers (Table 2). Correlations with treatment duration were found for general ADA titer, IgG subclasses 1, 2, and 3, and neutralizing ADA titer; for RAV, it was found only in the patient groups having persistent neutralizing ADA (Table 3).
Data are shown as the Spearman's correlation coefficient.
For definition of patient groups, see Materials and Methods section.
The correlation was assessed in study subgroups with neutralizing ADA.
The correlation was assessed in all patients.
Indicates statistical significance at a P value <0.001.
Indicates statistical significance at a P value <0.01.
Indicates statistical significance at a P value <0.05.
n.s., statistically non-significant; RAV, relative affinity values.
Data are shown as the Spearman's correlation coefficient.
For definition of patient groups, see Materials and Methods section.
Indicates statistical significance at a P value <0.05.
Indicates statistical significance at a P value <0.001.
Indicates statistical significance at a P value <0.01.
n.s., statistically non-significant.
Using a multivariate regression model, neutralizing ADA titers at mo24 and mo36 (Beta 0.914 and 0.823, respectively, both P ≤ 0.001) as well as an increase in RAV between mo12 and mo24 (Beta 0.459, P = 0.007) were determined to be independent predictors for neutralizing ADA persistency.
Discussion
Here, we investigated the development of ADA titers, IgG1–4 titers, and ADA binding affinities with regard to the evolution and persistency of neutralizing ADA in s.c. IFN-β-treated RRMS patients. Although some of the facts shown in the present study have been described, in principle, earlier, for example, titer-dependent kinetics of neutralizing ADA (Hegen and others 2012) or differences in antibody affinity (Gneiss and others 2006a) and IgG subclasses between neutralizing ADA-positive and -negative patients (Deisenhammer and others 2001; Gibbs and Oger 2007), this study revealed important further information on ADA activities and their evolution by providing the first longitudinal examination of the relationship between 3 critical ADA characteristics in a single, large, and well-characterized RRMS patient cohort.
It is generally accepted that the biological effects of antibodies depend on their specificity to the target epitope, titer, and binding affinity. We observed that IFN-β-1b-treated patients developing persistent neutralizing ADA (after 2 years of therapy) show early (within 6 months) and high peak ADA titers, similar as in previous studies (Mayr and others 2003; Hegen and others 2014). We also confirmed that antibody binding affinity is significantly higher in neutralizing ADA-positive than in -negative patients (Gneiss and others 2006a) and directly correlates with neutralizing ADA titer (Gibbs and Oger 2008). Of interest is that ADA binding affinity increases with treatment duration only in the ultimately persistent neutralizing ADA patient group, and not in patients with transient neutralizing ADA or non-neutralizing ADA. Such an observation has never been reported.
Based on these observations, one might hypothesize that ADA titer plays a role in regulating mechanisms of antibody affinity maturation (Pierson-Mullany and others 2002; Jackola and others 2005). Indeed, it has been previously suggested that, given a relatively constant antigen load, there should be some optimal combination of antibody concentration and binding affinity, since the most suitable immune effectiveness may be gained by either increasing the production of specific antibodies or enhancing antibody binding affinity (Jackola and others 2005). This mechanism is further supported by our observation in patients who will eventually develop neutralizing ADA titers that ADA titers show a strong increase at a time of lower antibody affinity. Thereafter, ADA titers decrease when neutralizing activity (ie, neutralizing ADA) becomes established, as shown by the negative correlation between antibody binding affinity and ADA titers, and a positive correlation with neutralizing ADA titers. The latter phenomenon has also been observed in other conditions, for example, in allergen-sensitive atopic individuals showing an inverse relationship between allergen-specific Ig concentration and allergen-Ig-binding affinity (Pierson-Mullany and others 2002), observed as a negative feedback loop between antibody concentration and affinity (Germain 2001; Jackola and others 2005). In such an immunological milieu, when the antigen becomes limited, B cells bearing high-affinity receptors are preferentially stimulated (so-called antigen driven selection) (Kaattari and others 2002). Accordingly, we observed an increase of antibody binding affinity (RAV) within year 2 in patients with later persistent neutralizing ADA; this increase was an independent and significant predictor of persistent neutralizing ADA. Such an observation is consistent with the typical time period known to be associated with the evolution of neutralizing ADA (Deisenhammer 2009; Polman and others 2010).
Another determinant for developing (persistent) neutralizing ADA is probably the IgG subclass profile. We observed that patients with non-neutralizing or transient neutralizing ADA predominantly showed ADA within the IgG1 and IgG3 subclasses, whereas IgG2 and IgG4 were undetectable or maintained at very low levels over 3 years of IFN-β treatment. Suppression of B cell activation within the immune response against IFN-β could, at least in part, be mediated by antibody subclass feedback mechanisms, as IgG1 and IgG3 subclasses are believed to have the higher capacity to negatively regulate B cell activation (Gibbs and Oger 2007). Our finding of a weak, but statistically significant negative correlation between ADA IgG3 titers and antibody affinity further supports this hypothesis (Gibbs and Oger 2007). In persistently neutralizing ADA-positive patients, there was a predominance of IgG2 and IgG4 production, and such findings are consistent with previous studies (Deisenhammer and others 2001; Gibbs and Oger 2007). Since we found a positive correlation of ADA IgG4 titers with both antibody affinity and neutralizing ADA titers, we hypothesize that the shift in the immune response toward IgG4 production is a main contributing factor to neutralizing ADA persistency.
ADA characteristics of patients with persistent neutralizing ADA against s.c. IFN-β-1a were similar compared with the s.c. IFN-β-1b group. However, one has to consider a different temporal antibody evolution developing later peak ADA titers, a later increase in neutralizing capacity, and different ADA reversion rates in RRMS patients treated with IFN-β-1a (Perini and others 2004; Hegen and others 2012). One might speculate that the relative lack of ADA IgG3 subclass (Gibbs and Oger 2007), also shown in our study, might contribute to a lower reversion rate to neutralizing ADA negativity in IFN-β-1a-treated patients (Sorensen and others 2005; Hegen and others 2012).
The limitations possibly associated with this study include the retrospective study design and selection of patients, which was restricted to include available serum samples (IFN-β-1b pivotal trial and the Austrian ADA registry). With regard to laboratory methods, there are also some aspects that have to be further illuminated. Two different assays, the CPE and MxA induction assay, were used for the detection of neutralizing ADA. We considered them to give comparable results since a correlation between neutralizing ADA titers determined by the 2 methods has been previously reported (Pungor and others 1998), and we found a favorable concordance between neutralizing ADA titers obtained by the MxA induction assay and the CPE assay in a subset of samples. Regarding ADA and the antibody affinity assay, IFN-β-1b was used as an antigen. This may have resulted in lower antibody-binding responses than if IFN-β-1a would have been used (Files and others 2007). However, such an effect would not be expected to impact the observed differences found between patient groups, as it is similarly applied to all results (also independent of the type of IFN-β used for patient treatment). Regarding antibody affinity, these analyses [and all previous (Gneiss and others 2006a, Gibbs and Oger 2008)] were limited by the fact that actual ADA concentrations are unknown precluding the calculation of absolute affinity; thus, RAV (based on dissociation rates) were used. We are convinced that different antibody affinities found between the patient groups are very unlikely to result from different ADA titers per sé, as the amount of antibodies used in the RAV assay was similar across all samples due to the design of the assay (using in each case that serum dilution with a differential OD of 0.7–1.3). The observed differences in the longitudinal RAV pattern between IFN-β-1b-treated patients with transient and persistent neutralizing ADA could not be attributed to treatment duration, as treatment duration in study subgroups was similar (Table 1). Furthermore, RAV did not significantly differ between the 2 IFN-β-1b dosing subgroups in both transitory and persistently neutralizing ADA-positive patients (data not shown). Finally, the present findings mainly apply to IFN-β-1b-treated patients, as only for this IFN-β preparation we had patients with different ADA (extending from non-neutralizing, transient to persistent neutralizing ADA).
Here, we demonstrated that neutralizing ADA evolution and persistency depend on a complex interplay between ADA titers, affinity maturation, and IgG subclass switch. Why some patients show certain dynamics of ADA and others do not needs further investigation. Nevertheless, it appears likely that a given MS patient's genetic disposition determines ADA development (and probably persistency), as certain human leukocyte antigen alleles and single nucleotide polymorphisms have been found to be associated with neutralizing activity and titer of anti-IFN-β antibodies (Hoffmann and others 2008; Buck and others 2011; Weber and others 2012).
Footnotes
Acknowledgment
The study was supported by a research grant from Bayer HealthCare.
Author Disclosure Statement
I.D. received lecture fees and/or travel grants from Merck Serono, Bayer Pharma AG, Medis, Teva, and Boehringer Ingelheim; received honoraria for acting as an advisor for Bayer Pharma AG; and was supported by the “Prof. Risto Bokonjic Foundation” and The Ministry of Education, Science and Technological Development of the Republic of Serbia (Grant No. 175031). H.H. has participated in meetings sponsored by, received speaker honoraria or travel funding from Bayer Pharma AG, Biogen, Merck Serono, and Novartis, and received honoraria for acting as consultant for Teva Pharmaceuticals Europe. P.P. is employed by Bayer HealthCare. E.C. has participated in meetings sponsored by, received consultant fee, speaker honoraria, or travel funding from Bayer Healthcare, Teva Pharmaceuticals, Atara Biotherapeutics, and Glialogix. F.D. has participated in meetings sponsored by or received honoraria for acting as an advisor/speaker for Bayer Healthcare, Biogen Idec, Genzyme-Sanofi, Merck, Novartis Pharma, and Roche.