
Abstract
Mycobacterium tuberculosis PE/PPE family proteins play a vital role in antigenic diversity, host–pathogen interactions, and immune evasion. As secreted by ESX-5 system, M. tuberculosis PPE27 is related to the growth and virulence of the bacilli. In this study, we expressed PPE27 in the nonpathogenic fast growing Mycobacterium smegmatis. We found that the recombinant strain exhibits higher survival rate under several hostile conditions in vitro and longer persistence in mouse tissues. The survival of the recombinant strains was also enhanced in the mouse macrophage ANA-1, accompanied by higher level of host cell death, higher secretion of tumor necrosis factor-alpha, and a slightly higher amount of interleukin (IL)-1β, which could be abolished by the nuclear factor-κB, p38, and ERK inhibitors. In addition, the recombinant strain robustly induced larger amount of nitric oxide (NO) and lower amount of IL-6 after infection of mouse macrophages. In brief, our data suggest that PPE27 promotes the survival of nonpathogenic M. smegmatis in vitro by manipulating the expression of multiple cytokines, NO, and affecting host cell necrosis, which provide a new insight to understand the functions of this gene.
Introduction
T
Mtb expresses multiple ligands which bind to different host cell receptors involved in entry, such as complement receptor, scavenger receptors, and Toll-like receptors TLR2 and TLR4 (Carroll 1998; Ryffel and others 2005). Exhaustive bioinformatics screening has suggested some proteins from the PE, PPE, and MCE families to be potentially exposed membrane proteins, many of which can affect host immune responses by binding to host cell receptors (Somvanshi and others 2008; Sampson 2011). The PE and PPE members in the mycobacteria represent about 10% of the total coding capacity of the Mtb genome, which was named by their
Mtb (Rv1790), locating in ESX-5 gene cluster, encodes a PPE family protein (PPE27). It has been reported that the ESX-5 system induces caspase independent cell death in macrophages and is indispensible for mycobacterial cell wall stability (Ates and others 2015; Shah and Briken 2016). ESX-5 systems also have been proven to be involved in protein secretion to allow the bacteria to exit the cells and infect neighboring cells (Voskuil and others 2004; Ates and others 2016; Shah and Briken 2016). The ESX-5 region of Mtb (rv1782–rv1798) harbors 2 pe (pe18/19) and 3 ppe (ppe25/26/27) genes (Shah and Briken 2016). MtbΔppe25-pe19 was found highly attenuated in immune-competent mice (Bottai and others 2012; Sayes and others 2012). MtbΔppe27-pe19 deletion caused a growth defect in standard Pi-rich 7H9 medium and formed smaller colonies on 7H10 solid agar medium (Ramakrishnan and others 2016). The evidence above indicates that PE/PPE proteins of ESX-5 may play important roles in virulence of mycobacterium. Comparative genomics analyses suggest that PPE27 may be associated with pathogenesis (Voskuil and others 2004; Bottai and others 2012; Ramakrishnan and others 2016), and PPE27 mutant can affect an increasing secretion of PPE41 and some PE_PGRS proteins (Houben and others 2012). However, the functions of the PPE27 remain unclear.
The rapid growth and ready expression of heterogeneous genes from pathogenic mycobacteria enable the nonpathogenic Mycobacterium smegmatis a good surrogate for the pathogenic mycobacteria (Daim and others 2011). In the present study, we expressed PPE27 in M. smegmatis and investigated whether PPE27 can enhance the survival of recombinant M. smegmatis in the mouse macrophage cell, various stress conditions, and mouse tissues, and also the release of cytokines in infected mouse, nitric oxide (NO), and the secreted cytokines from the infected macrophages was analyzed in this study.
Materials and Methods
Cell culture, bacterial strains, and animals
Mouse ANA-1 cells were supplied by RPMI-1640 (Hyclone, USA) medium with 10% fetal calf serum (FCS; Hyclone) containing 100 U/mL streptomycin and 100 U/mL penicillin (Hyclone); the cells were incubated in a humidified atmosphere with 5% CO2 at 37°C. Escherichia coli DH5α (TIANGEN, China) was cultured in Lennox LB medium for DNA cloning. M. smegmatis strain mc2155 and recombinant M. smegmatis were cultured at 37°C on Middlebrook 7H9 (MB 7H9; Difco, USA) liquid medium with shaking or on Middlebrook 7H10 (MB 7H10; Difco) agar supplemented with 0.5% (w/v) albumin fraction V, 0.2% (w/v) glucose, 0.85% (w/v) NaCl, 0.2% (v/v) glycerol, and 0.05% (v/v) Tween 80; 25 μg/mL kanamycin was added to detecting the gene as well (Dheenadhayalan and others 2006). Six- to eight-week-aged BALB/c male mice, received from the Sichuan University Laboratory Animal Center (Chengdu, China), were dedicated in experiments according to Laboratory Animal Care and approved by the Ethics Review Committee of SLAS. Appropriate diet and sterilized water were provided for animals, which, at the same time, were bred in the West China Center of Medical Sciences in sterile conditions.
Gene amplification, plasmid construction, and recombinant M. smegmatis
Chromosomal DNA was from M. tuberculosis genome, in which the open reading frame encoding M. tuberculosis (PPE27) Rv1790 was amplified by polymerase chain reaction (PCR) with gene-specific primers. To acquire his6-tagged PPE27 fusing protein, the his6-tagged PPE27 gene was PCR amplified with the forward primer 5′-GGAATTCATGGACT TCGGGGCGTTACCGCC-3′ containing an EcoRI site (underlined) and reverse primer 5′-gtggtggtggtggtgTCCCGC CGACGGAG-3′ containing five His codons (lowercase letters). The containing his5-tagged PPE27 sequence was diluted as the second PCR template, using the same forward primer and a second reverse primer 5′-CCCAAGCTTCT Agtggtggtggtggtggtg-3′, containing a HindIII site (underlined) and six His codons (lowercase letters), then the containing six consecutive His codons PCR product was inserted in frame into the cloning site of pMV361 and generated as pMV361-his6-tagged PPE27 (Daim and others 2011). The recombinant plasmid and empty pMV361 was electroporated into M. smegmatis mc2155, respectively. Recombinant M. smegmatis clones were selected on MB 7H10 agar containing 25 μg/mL kanamycin. The clone containing PPE27 gene was named as Ms_ppe27, and the clone harboring empty pMV361 alone was named as Ms_vec. Ms_ ppe27 and Ms_vec were cultured until the OD600 (the absorbed light of material) reached 0.8–1.0 with shaking, and the bacterial pellets were harvested and disrupted by ultrasonication. Protein samples were subjected to sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE), and the his6-tagged PPE27 protein was detected by Western blotting with mouse anti-His antibody (TIANGEN), while Ms_vec was set as a control.
In vitro growth of M. smegmatis and subcellular location of the protein
To monitor the growth tendency of the recombinant M. smegmatis strains, Ms_vec and Ms_ppe27 were shaken at 37°C in MB 7H9 liquid medium in a starting absorbance (OD600) of 0.02. The OD600 of the culture was continuously determined at an interval of 3 h over 24-h growth period.
Subcellular localization of the PPE27 protein was determined by protocols previously described (He and others 2015). Briefly, M. smegmatis was cultured in 50 mL 7H9 liquid medium until log phase. To prepare the whole cell lysate, the cell pellets were harvested and lysed by sonication, in which intact cells and cell debris were removed by centrifugation at 3,000g for 5 min. Then the supernatant cells were subjected to ultracentrifugation at 12,000g at 4°C for 60 min to obtain the pellets (cell wall fraction), and the supernatants (cytoplasm fraction) were transferred into fresh tube. Proteins from each fraction were subjected to SDS-PAGE and analyzed by Western blotting with mouse anti-His antibody. Native GroEL was used as a positive control for detecting cytoplasm fraction (Daleke and others 2012). Positive GroEL was detected using mycobacterial Hsp65 mouse primary antibody (Abcam), while HRP-conjugated Goat Anti-Mouse IgG (TIANGEN) was used as the secondary antibody.
In vitro infection with recombinant M. smegmatis
ANA-1 cells were seeded in 12 well tissue with a density of 5 × 105 per well in RPMI-1640 medium containing 10% FCS and 100 U/mL streptomycin and 100 U/mL penicillin, and cells were incubated for 24 h. Exponentially growing bacteria cultured in the presence of 25 μg/mL kanamycin were pelleted and suspended in RPMI-1640 medium in a single-cell suspension. Then, ANA-1 cells were infected with M. smegmatis transformants at a multiplicity of infection (MOI) of 10:1 (bacteria-to-ANA-1 ratio) and incubated for 4 h at 37°C under 5% CO2. After phagocytosis, cells were washed thrice with sterile warm PBS to remove extracellular bacteria and then incubated with fresh RPMI-1640 containing 4% FCS and 100 μg/mL gentamicin for 2 h. After that, cells were washed thrice with sterile warm PBS and cultured in fresh medium with 4% FCS (Daim and others 2011).
At different time points (6, 24, or 48 h), the ANA-1 macrophages were collected and washed twice with sterile warm PBS and then lysed with 1 mL of PBS with 0.025% (w/v) SDS. The lysed macrophages were serially diluted in PBS with 0.05% Tween 80 (PBS-T) and plated on 7H10 agar plates containing 25 μg/mL kanamycin; the colony forming units (CFUs) were counted as a measurement of the intracellular survival of recombinant M. smegmatis.
Analysis in vitro survival under different stress conditions
Recombinant M. smegmatis strains were grown into log phase on 7H9 medium containing 25 μg/mL kanamycin. Bacteria were collected and washed with 7H9 by adding HCL and then diluted to an OD600 of 0.5 in 5 mL 7H9 medium (pH 5.0). Cultures were incubated for 0, 3, or 6 h under stress conditions at 37°C (Singh and others 2016). To determine the survival of recombinant M. smegmatis after exposure to peroxide stress, bacteria were washed and suspended on 7H9 and then diluted to OD600 of 0.8 in 7H9. The bacteria were then treated with SDS at the concentration up to 0.1% for 2 and 4 h (Ramakrishnan and others 2016). To measure the sensitivity to hydrogen peroxide (H2O2), the bacteria (OD = 0.5) were incubated at 37°C with 5 mM H2O2 (Sigma, USA) for 3 and 6 h, and then 100 μL samples were removed to determine the viable count. To measure sensitivity to lysozyme, the bacteria (OD = 0.5) were incubated at 37°C with 2,500 mg/L lysozyme (Sigma-Aldrich, USA) for 6 h. To test susceptibility to nitrosative stress, the bacteria (OD = 0.5) were incubated at pH 5.4 with 10 mM sodium nitrite (NaNO2) for 0, 2, or 4 h at 37°C, and then 100 μL samples were removed to determine the viable count (Singh and others 2016).
In vivo infection with recombinant M. smegmatis
To evaluate the in vivo survival of the recombinant M. smegmatis, 6-week-old male BALB/c mice (seven per group) were infected with 2 × 107 CFUs of either Ms_vec or Ms_ppe27 by intraperitoneal injection (Dheenadhayalan and others 2006). All the strains were dispersed in 0.05% PBS-T, and no clumping was observed. At each time point (days 3, 6, and 9), survival of the recombinant M. smegmatis strains in different organs was determined with the methods as described previously (Singh and others 2013). Briefly, the mice were sacrificed, and lungs, livers, and spleens were removed aseptically and homogenized in sterile 0.05% PBS-T. The mouse tissue homogenates were diluted in PBS-T and 100 μL aliquots (or 200 μL for lung homogenates), and then the dilutions were plated on MB 7H10 agar plate containing 10% OADC and 25 μg/mL kanamycin. The plates were incubated at 37°C, and the CFUs were counted after 3 to 4 days.
Measurement of lactate dehydrogenase release from macrophage
Culture supernatants were harvested after infection of macrophages with Ms _ppe27 or Ms_vec for 6, 24, or 48 h. Lactate dehydrogenase (LDH) activity in the culture supernatants was assayed with a LDH Cytotoxicity Detection Kit assay (Promega, USA) according to the manufacturer's instructions. The percentage of LDH release was calculated according to the published method. The percentage of LDH release was calculated as follows: percentage release = 100 × (experimental LDH release-spontaneous LDH release)/(maximal LDH release-spontaneous LDH release). The value of maximal LDH release was obtained from the complete lysis of uninfected ANA-1 macrophages, and the Spontaneous LDH release was measured with supernatants of uninfected cells.
Assay for cytokine and NO production
Cultured supernatants were harvested at various time points (6, 18, or 48 h) after infection of ANA-1 cells with Ms_ppe27 or Ms_vec, respectively. Concentrations of IFN-γ, tumor necrosis factor-α (TNF-α), IL-1β, IL-2, IL-4, IL-6, IL-10, and IL-12 in the culture supernatants were determined with a two-site sandwich EIA (eBioscience, USA) according to the manufacturer's protocol. NO concentration in the culture supernatants was detected at varying time intervals (6, 24, or 48 h) after infection of ANA-1 cells with Ms_ppe27 or Ms_vec using the Griess reagent system (Promega) according to the manufacturer's instructions.
Quantitative real-time PCR (qRT-PCR) was used to detect the messenger RNA (mRNA) levels of the cytokines and inducible NO synthase (iNOS). After infection of macrophages with Ms_ppe27 or Ms_vec for 24 h, total RNA was extracted with the Simply P Total RNA Extraction Kit (Bioer, China). The RNA was then transcribed into complementary DNA (cDNA) with random hexamers using the RevertAid First Strand cDNA Synthesis Kit (Thermo, USA). qRT-PCR was performed with primers listed in (Supplementary Table S1; Supplementary Data are available online at
Pharmacological inhibition of signaling
All the pharmacological inhibitors were reconstituted in sterile dimethyl sulfoxide (DMSO; Sigma) and then used in the following concentrations: 10 μM U0126 (a MEK1/2 inhibitor; Sigma), 20 μM SB202190 (a p38 inhibitor; Sigma), and 20 μM Bay11-7082 (a nuclear factor-κB (NF-κB) inhibitor; Sigma); DMSO at 0.1% concentration was used as the vehicle control. One hour after treatment with the inhibitor, the macrophages were infected with Ms_vec or Ms_ppe27. The culture supernatants were harvested after 24 h treatment, and enzyme-linked immunosorbent assay (ELISA) was performed to determine the concentrations of TNF-α and IL-1β.
Assay for serum cytokines in infected BALB/c mouse
The BALB/c mouse was infected with Ms_ppe27, Ms_vec, or PBS-T by intraperitoneal injection; all the strains were dispersed in 0.05% PBS-T and delivered to each animal at 1 × 108/CFU (Singh and others 2016). At days 3, 6, 9, and 12 after infection, the blood was collected from the mice eyes, and the serum was collected by centrifugation at 2,500g for 5 min (Sha and others 2017). A total of three mice were sacrificed for each time point of individual group. The level of serum cytokines TNF-α, IL-1β, and IL-6 was determined using an ELISA Kit (eBioscience) according to the manufacturer's instructions.
Statistical analysis
All the experiments were performed in triplicate. Significance analysis was performed by SPSS (Statistical Package for the Social Sciences, version 19.0; SPSS, Inc.). Statistical significance was assessed by one-way analysis of variance, followed by Bonferroni's post hoc test. P value <0.05 was considered as statistically significant, and P < 0.01 was considered as highly significant.
Results
The expression of PPE27 in recombinant M. smegmatis
To investigate the effect of PPE27 on the macrophage response, we expressed it in vitro. We generated a recombinant M. smegmatis strain Ms_ppe27 that expresses his6-tagged PPE27. A control strain Ms_vec that only contains vector pMV_361 was also constructed. We confirmed that the his6-tagged PPE27 gene was electroporated into M. smegmatis successfully and absent in the control strain Ms_vec by PCR (Supplementary Fig. S1). The expression of his6-tagged PPE27 fusing protein (∼41.0 kDa) in strain Ms_ppe27 was confirmed by Western blotting with the mouse anti-His antibody (Fig. 1A). To determine whether the protein exists in cell wall or in cytoplasm fraction, strain Ms_ppe27 was lysed and protein fractions were separated and analyzed by Western blot. The results suggested that the PPE27 protein was exposed on the cell wall. As controls for fractionation, the cytosolic heat-shock protein GroEL was only detected in the cytoplasm fraction (Fig. 1B).

Expression of PPE27 in recombinant Mycobacterium smegmatis.
PPE27 enhances the survival of recombinant M. smegmatis in macrophages
To explore whether PPE27 can enhance the survival of the pathogen within host macrophages, we investigated the intracellular survival of the M. smegmatis harboring the PPE27. ANA-1 macrophages were infected by strain Ms_vec and Ms_ppe27 at a MOI of 10:1 as described in the methods part. At 24 and 48 h postinfection, compared with the strain Ms_vec, strain Ms_ppe27 had a greater number of bacteria residing within macrophages (Fig. 2A). We also found that there was no significant difference in growth kinetics between the two recombinant strains (Fig. 2B), suggesting that expression of PPE27 did not influence the growth of M. smegmatis.
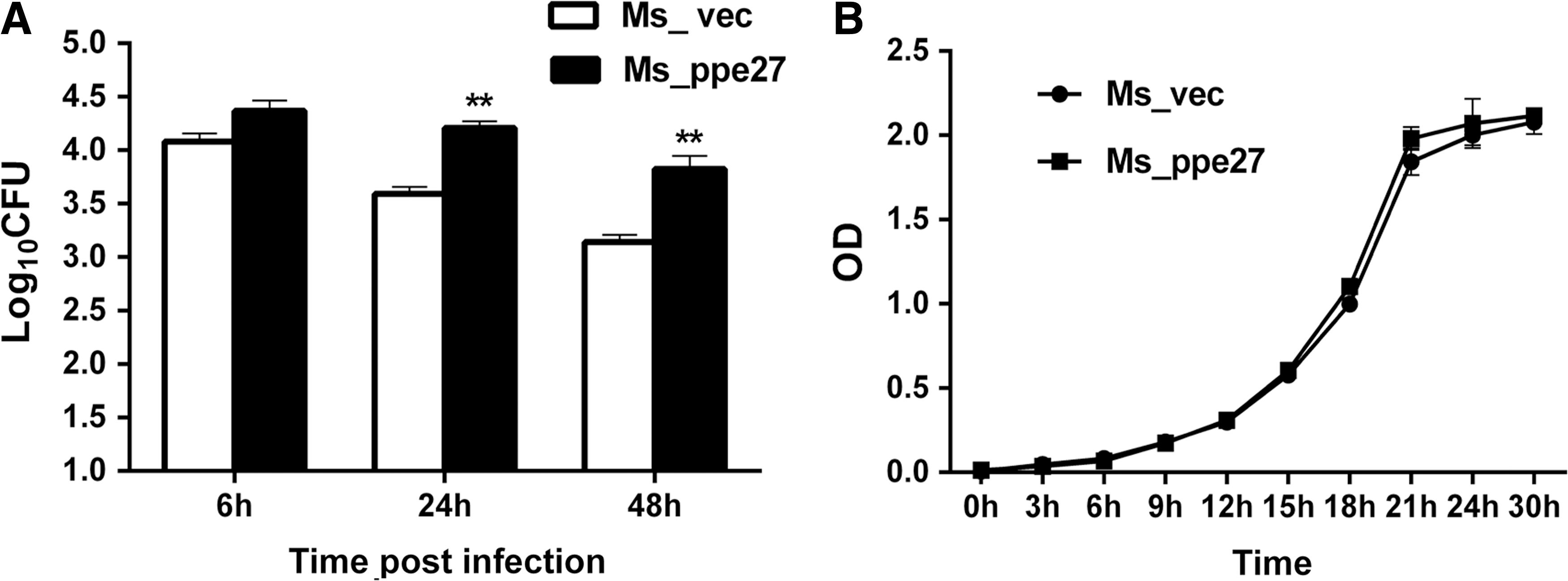
Intracellular survival of Ms_vec and Ms_ppe27 in ANA-1 macrophages and the growth curve of the recombinant strains.
PPE27 promotes the survival of recombinant M. smegmatis resistance to hostile factors
To understand the mechanism of PPE27 mediated enhancement of M. smegmatis survival in macrophage, we evaluated the resistance capacity of the recombinant strain in several stress conditions, including low pH, 0.1% SDS, 5 mM H2O2, and 2,500 mg/L lysozyme. The results showed that Ms_ppe27 was more resistant than Ms_vec on 7H9 medium at pH5.0 after being incubated for 3 or 6 h (Fig. 3A). SDS susceptibility test showed that the survival rate of Ms_ppe27 was significantly higher compared with Ms_vec at 2 or 4 h after exposure (Fig. 3B). Similarly, we found that strain Ms_ppe27 was more resistant to H2O2, lysozyme, and NaNO2 than the control strain (Fig. 3C–E). Taken together, these results suggested that PPE27 enabled M. smegmatis a higher resistance to several hostile factors.
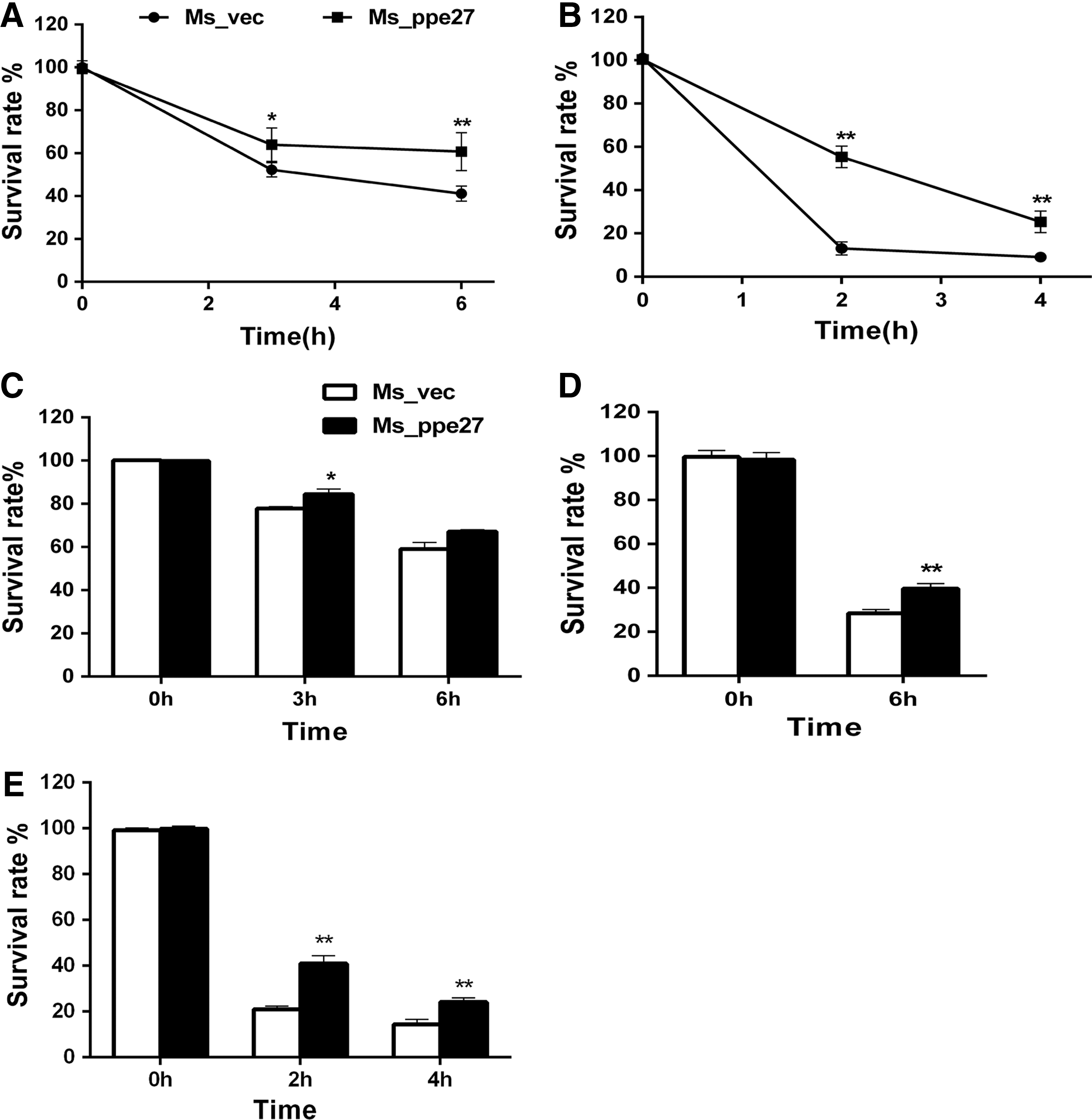
Survival of Ms_vec and Ms_ppe27 under different stress conditions.
PPE27 confers a growth advantage to M. smegmatis in mouse
To observe the survival of Ms_ppe27 in mouse tissues, we infected BALB/c mice with Ms_ppe27 or Ms_vec strains and compared their CFU in different tissues. Strain Ms_vec and Ms_ppe27 were dispersed into single cell suspensions and administration of equal number of bacteria into mice (Fig. 4A). We found that a significantly greater number of Ms_ppe27 presents in the liver, spleen, and lung tissues compared to Ms_vec (Fig. 4B–D). The recombinant M. smegmatis strains were cleared more slowly in the liver (Fig. 4B) and spleen (Fig. 4C). In the lung tissues, the bacteria were almost cleared out at 9 days postinfection (data not show). No difference was observed in the size of spleen between the two groups. These results indicated that the expression of PPE27 in nonpathogenic M. smegmatis resulted in enhanced survival of the bacilli in host tissues.
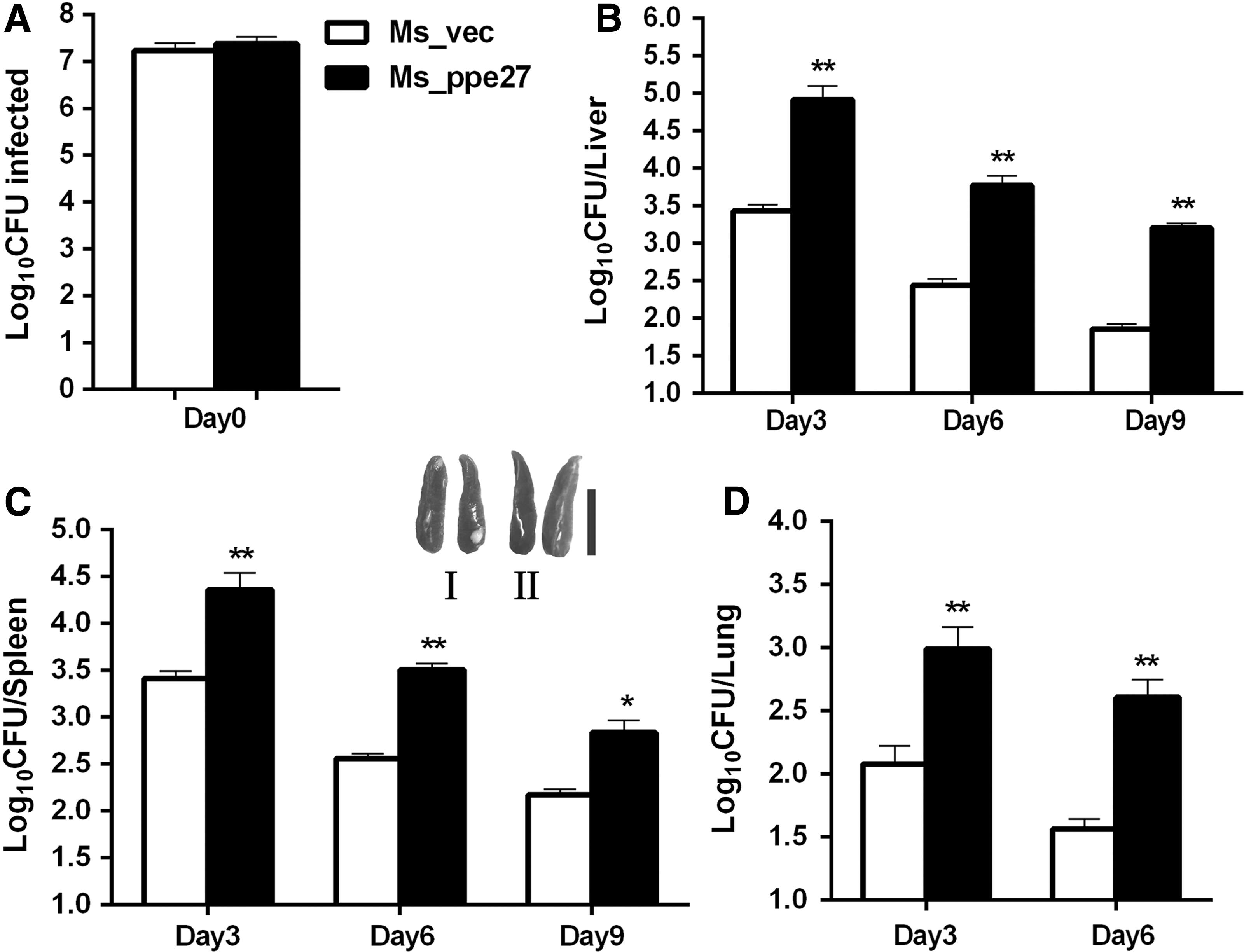
The persistence of Ms_vec and Ms_ppe27 in mouse tissues.
PPE27 triggers cell necrosis during infection with M. smegmatis
To test whether PPE27 has effectiveness on macrophage viability, we infected ANA-1 macrophages with recombinant M. smegmatis and detected the amount of LDH in the culture supernatants to quantify cell necrosis. The results showed that both the experiment and control groups released comparable low amounts of LDH in 6 h postinfection. At 24 and 48 h postinfection, macrophages infected by Ms_ppe27 showed higher LDH release amount than the control group with appreciable difference, 22.7% versus 31% in 24 h and 41.5% versus 51.2% in 48 h (Fig. 5).

Assay of cell death in macrophages infected with Ms_vec and Ms_ppe27. Culture supernatants were collected from ANA-1 infected at a MOI of 10:1 with Ms_vec and Ms_ppe27, respectively; the release of LDH as a measure standard of macrophage death was estimated at different time points. Error bars represent (±SEM) from three biological replicates. *P < 0.05, **P < 0.01. LDH, lactate dehydrogenase; MOI, multiplicity of infection; SEM, standard error of the mean.
PPE27 upregulates TNF-α and downregulates IL-6 in ANA-1 macrophages
To explore the influence of PPE27 on host innate immunity, we investigated the levels of several pro-inflammatory cytokines upon infections of ANA-1 macrophages. Macrophages infected with Ms_ppe27 secreted significantly higher level of TNF-α (Fig. 6A) at 6 h postinfection, and a higher level of IL-1β was secreted by the Ms_ppe27 infected after 6 and 18 h compared with the Ms_vec, although the difference was around 3 pg/mL (Fig. 6B). Furthermore, Ms_ppe27 induced a lower level of IL-6 in infected macrophages compared with Ms_vec (Fig. 6C). No difference was observed in the levels of IL-4 and IL-10 between two groups (data not show), and the level of IFN-γ, IL-2, and IL-12 could not be detected in the groups. The levels of above cytokines were further determined by qRT-PCR. Consistent with the results of ELISA, we detected higher mRNA levels of TNF-α and lower level of IL-6 in macrophages after 24 h for the infection of Ms_ppe27. No significant difference was observed in transcript levels of IL-1β (Fig. 6D). These results suggest that PPE27 might play a role in modulating pro-inflammatory cytokine balance to alter the outcome of the infection.

Cytokines and nitric oxide release of ANA-1 macrophage infected by the recombinant strains. TNF-α
Macrophages can defense the M. tuberculosis infection by producing superoxide radicals such as NO (Bhat and others 2013). NO and the transcript level of iNOS were measured to identify whether nitrite accumulation contributes to intracellular survival of recombinant M. smegmatis strains in macrophages. We observed an upregulation of the iNOS transcript level (Fig. 6D) and a significant increase in NO production (Fig. 6E) in ANA-1 cells after Ms_ppe27 infection compared to Ms_vec after 24 h infection, which indicated that the enhanced intracellular survival by PPE27 is associated with the increased production of NO.
PPE27 possibly activates macrophage ERK, p38, and the NF-κB signaling
It was reported that the MAPKs and the NF-κB signaling pathways have participated in production of some pro-inflammatory cytokines in M. smegmatis-infected macrophages (Collart and others 1990; Lee and Schorey 2005). We investigated the possible involvement of the MAPKs and NF-κB pathways in PPE27 inducing pro-inflammatory responses by ELISA. Treated with pharmacological inhibitors, macrophages were infected with Ms_ppe27 or Ms_vec for 24 h to assess TNF-α and IL-1β production. Data showed a lower level of the TNF-α (Fig. 6F) and IL-1β (Fig. 6G) in macrophages pretreated with the ERK1/2 inhibitor (U0126), the NF-κB inhibitor (Bay11-7082), or the p38 inhibitor (SB203580). Based on these results, we suggest that induction of cytokines by Ms_ppe27 in macrophages was possibly mediated by MAPK and NF-κB signaling pathway.
PPE27 modulates circulating cytokine secretion in serum of infected mouse
To investigate the effect of PPE27 in infected mouse response, we analyzed the circulating cytokine secretion in mouse serum after being infected by the recombinant strains at 3, 6, 9, and 12 days. It was found that the levels of TNF-α were significantly higher in the mouse infected with Ms_ppe27 compared to those of mice infected with Ms_vec during 6–12 days postinfection (Fig. 7A). Meanwhile, Ms_ppe27 reduced the secretion of pro-inflammatory cytokine IL-6 during 6–12 days postinfection compared with Ms_vec (Fig. 7B). Besides, there was no difference in the levels of IL-1β during the infection (Fig. 7C). The results indicated that the enhanced survival rate in mouse tissues by PPE27 is associated with the medicating of the cytokine release in infected mouse.

Serum cytokines in Ms_ppe27 and Ms_vec infected mice. The serum of the mice infected with Ms_ppe27, Ms_vec, or PBS with 0.05% Tween 80 was collected at different time points (3, 6, 9, and 12 days) postinfection. The levels of cytokines TNF-α
Discussion
The capability of M. tuberculosis survival and persistence within host macrophages is the major factor of the pathogenesis of the M. tuberculosis (Liu and Modlin 2008; Cambier and others 2014b). Many investigations indicate that PPE proteins have participated in such process (Dong and others 2012; Tundup and others 2014). In the present study, we found that expression of PPE27 in M. smegmatis enhanced the survival in ANA-1 macrophage cultures in vitro, promoted the resistance in stress conditions, and enabled M. smegmatis longer persistence in mouse tissues in vivo. These results indicate that PPE27 confers M. smegmatis with potential pathogenic properties, consistent with the report that PE/PPE proteins of ESX-5 secretion system are vital to M. tuberculosis survival and virulence (Bottai and others 2012; Sayes and others 2012; Ramakrishnan and others 2016).
M. tuberculosis can induce necrosis to survive and disseminate in the infection site (Cambier and others 2014a). It was reported that PE25/PPE41 proteins induce macrophage necrosis to help the bacterium to survive in the host (Tundup and others 2014). Our study has shown that macrophage viability is impaired by expression of PPE27. Such result implies that PPE27 could induce necrosis to allow the bacteria to survive in macrophage and evade host immunity.
The cell wall constituent of M. tuberculosis has been implicated in the pathogenesis and resistance to hostile factors (Akhter and others 2012; Singh and others 2016). Several PPE proteins, such as Rv2108 (PPE36), Rv1787 (PPE25), and Rv1196 (PPE18), have been observed to be cell wall associated and may play crucial roles in host–pathogen interactions (Le Moigne and others 2005; Bhat and others 2012; Mi and others 2016). The evidence shows the host macrophages defense against M. tuberculosis, including reactive oxygen and reactive nitrogen compounds, iron-deprived conditions, and low pH conditions (Singh and others 2016). In this study, we observed that PPE27 is associated with the cell wall. Furthermore, overexpression of PPE27 in M. smegmatis enhances the survival rates in the environments of SDS, H2O2, lysozyme, low pH, and NaNO2, which further support their role in the host interaction and resistance to different stress conditions.
Macrophage is the primary target for M. tuberculosis infection and persistence (Liu and Modlin 2008). The reaction of innate immunity and cytokines secreted by macrophages is crucial for the outcome of M. tuberculosis infection (Arango Duque and Descoteaux 2014). M. tuberculosis can subvert the cytokine release for its survival (Cambier and others 2014a). Many PPE proteins have been revealed to have the capability to induce inflammatory cytokines. PPE38, a virulent associated protein, can induce high levels of TNF-α and IL-6 secretion in macrophage (Dong and others 2012); PPE57 is able to recognize TLR2 and induce pro-inflammatory cytokines like TNF-α, IL-6, and IL-12p40 within macrophages (Xu and others 2015). In our study, high level of TNF-α was secreted from ANA-1 macrophage cultures infected with M. smegmatis expressing PPE27. High levels of TNF-α are thought to promote the doom of M. smegmatis and assist hosts to terminate the invading nonpathogenic M. smegmatis (Clay and others 2008). However, we found that PPE27 increases the survival of recombinant M. smegmatis strains in macrophages, which support the complicated interactions between the M. tuberculosis PPE27 and host cell. In addition, the secretion of cytokine of IL-1β and IL-6 from infected macrophage also can be affected by PPE27, consistent with the report that macrophages can secrete some pro-inflammatory cytokines infected with M. smegmatis (Daim and others 2011; Mi and others 2016). Meanwhile, PPE27 promoted the secretion of TNF-α and reduced the release of IL-6, accompanying higher survival rate in infected mouse tissues. These results indicate that PPE27 can alter the levels of multiple cytokines that enable M. tuberculosis to survive and persist after infection.
Cells can respond to extracellular signals by transmitting intracellular instructions to coordinate appropriate responses, such as the NF-κB and MAPK signaling pathways, regulating production of a number of cytokines by the macrophages infected with M. tuberculosis (Collart and others 1990; Clay and others 2008). Xu and others (2015) demonstrated that PPE57 can lead to activation of MAPK pathways. In this study, we found that the NF-κB, ERK, and p38 pathways are involved in PPE27 induced TNF-α expression and IL-1β induction. The observations suggest that PPE27 is likely to disrupt the host signal transduction in an alternative manner to maintain the balance of cytokines, which enforce the bacteria to breakdown the host immune response.
Researches proved that NO plays an important role in innate immunity and defenses the microbiology to spread in active macrophage, while NO is generated by iNOS in the inflammatory response, appearing in the inflammatory-macrophage cells (Fang 1997; Bhat and others 2013). It was reported that PPE2 and PPE26 can inhibit NO in activated macrophages (Bhat and others 2013; Mi and others 2016). We found that PPE27 can promote the expression of iNOS and NO in macrophages, accompanying higher intracellular survival by Ms_ppe27 infection. As high concentrations of NO lead to cell damage and death, specifically, NO production inhibits respiration and induces dormancy of M. tuberculosis in the infection process (Voskuil and others 2003), which suggests that PPE27 affects the release of NO to mediate the survival of the mycobacterium.
In summary, our results suggest that the expression of PPE27 in nonpathogenic M. smegmatis shows some novel properties that can promote the persistence in vivo and in vitro. Such effects may be explained by the changed pro-inflammatory cytokine response and the viability of macrophages. Also these results intensify the acquaintance of the gene function of ESX-5 secreting system and ascertain the key role of PPE27 in the complex interplay between mycobacteria and host. Yet, many detailed functional roles of PPE27 remain to be determined. Further study using the PPE27-knockout M. tuberculosis to study the function of PPE27 is needed.
Footnotes
Acknowledgments
This work was supported by Grants from the Chinese National Key Project of Infectious Disease (2012ZX10003008004), the Fund of Doctoral Scientific Research of Ministry Of Education (MOE) (20110181110046), and the Natural Science Foundation of China (31301033).
Author Disclosure Statement
No competing financial interests exist.