
Abstract
Hemorrhagic shock (HS) is associated with an excessive activation of inflammation, contributing to multiple organ failure in numerous medical or surgical conditions. To explore the therapeutic potential of crocin, a natural compound with anti-inflammatory properties, we administered crocin to rats during resuscitation following HS induced by withdrawing blood. Compared with control animals which were sham-treated, HS-operated rats showed organ damages as manifested by enhanced markers of multiple organ dysfunctions. Crocin treatment substantially reduced these parameters in rats subjected to HS, suggesting an alleviation of tissue injuries such as in the kidney, liver, pancreas, and muscle. The activation of NF-κB (nuclear factor κB) pathway in lung tissue by HS, as shown by increased nuclear translocation of p65 and IκBα phosphorylation, was diminished by crocin treatment. The crocin administration also significantly decreased the serum levels of proinflammatory cytokine TNF-α (tumor necrosis factor-α) and interleukin (IL)-6, whereas increased the level of anti-inflammatory cytokine IL-10 in HS-operated rats. These studies indicate that crocin administration may reduce inflammation-driven tissue damage in patients with HS.
Introduction
H
An essential mediator of such devastating conditions in HS is the transcription pathway of nuclear factor κB (NF-κB), which correlates with the severity of systemic inflammatory response in HS-treated animals (Altavilla and others 2001; Yang and others 2002; Guarini and others 2003). A classic stimulation of NF-κB signaling pathway includes the activation of IκBα kinase and increased phosphorylation-associated IκBα protein degradation, thereby promoting the nuclear translocation of p65, resulting in transcriptional activation of its target genes such as inflammatory cytokines (Altavilla and others 2001; Yang and others 2002; Guarini and others 2003). The molecular basis of NF-κB activation by HS is still unclear, but NF-κB pathway is known to be associated with oxidative stress. Indeed, it has been previously suggested that reactive oxygen species (ROS) may be critical activators of NF-κB signaling, and reducing ROS level could prevent NF-κB activation in many occasions (Li and Karin 1999). Importantly, the blockade of NF-κB signaling is known to be able to protect HS-induced organ injury, which is associated with the reduction of proinflammatory mediators, including inducible nitric oxide synthase (iNOS) and cyclooxygenase-2 (COX-2) (Mauriz and others 2001; Guarini and others 2003). Particularly, activation of NF-κB pathway in the lung plays an essential function in initiating acute lung injury in HS through increased expression of proinflammatory cytokines and induced production of nitric oxide (NO). Moreover, NF-κB levels in lung tissue are also sensitive indicators of systemic inflammation that is tightly associated with local macrophage activation and neutrophil infiltration in the lung.
Owing to their anti-inflammatory activities, natural antioxidants may offer therapeutic promise for HS patients. For instance, a carotenoid compound named crocin, found in the crocus flowers and gardenia, could exhibit antioxidant and anti-inflammatory effects (Poma and others 2012). Prior studies have shown that crocin and its closely related chemical metabolite crocetin possess potential activities to reduce hyperlipidemia, block microglial activation, or protect cytotoxicity induced by ROS (Lee and others 2005; Mousavi and others 2010; Nam and others 2010). The beneficial effects of crocetin on HS have been well studied in animal models of HS. The increased whole-body oxygen consumption and reduced mitochondrial oxidative damage by crocetin may be responsible for these actions (Gainer and others 1993; Venkatraman and others 2008; Yan and others 2010; Wang and others 2012). In contrast, very little is known about the biological effects and therapeutic potential of crocin on HS. Previously, crocin has been demonstrated to exert protective effects against ischemic injuries in multiple organs, possibly through its free radical scavenging and antioxidant functions (Zheng and others 2007; Goyal and others 2010; Vakili and others 2014). For instance, in ischemic stroke (Ochiai and others 2007; Zheng and others 2007; Vakili and others 2014; Oruc and others 2016), crocin administration was shown to protect brain injury. In addition, the cardioprotective potential of crocin has been studied in an ischemia-reperfusion model of isolated rat hearts (Dianat and others 2014). Recently, the possible protective effect of crocin against IR-induced injuries in the kidney and liver has also been suggested by the studies of animal models of ischemia (Adali and others 2016; Ozkececi and others 2016). Potentially, the preventive effects of crocin against ischemia-induced organ injuries could be led by its potent inhibition on NF-κB activation, which has been extensively reported in studying the anti-inflammatory actions of crocin (Xu and others 2009; Nam and others 2010; Hemshekhar and others 2012). Based on these observations and strong associations between ischemia, oxidative stress, and multiorgan injuries in HS, we hypothesized that crocin may also have an effect in relieving HS-induced organ damage. To test these hypotheses, we administered crocin in a rat model of HS followed by resuscitation. We reasoned that crocin could create an immunomodulatory milieu that, in turn, may abrogate the systemic inflammatory response and protect multiorgan injuries induced by HS.
Methods and Materials
Animal model of HS and resuscitation
The animal protocols used in the study have been approved by the Animal Ethics Committee of Cangzhou City Central Hospital in accordance with the guidelines of the institutional animal care. Male Wistar rats, which were housed in a temperature-controlled environment with a 12-h light–dark cycle, were administered by general anesthesia (using intraperitoneal injection of 120 mg/kg sodium thiopentone, and maintained using 10 mg/kg intravenous). Following anesthesia, the trachea, bladder, carotid artery, jugular vein, femoral artery, and femoral vein were cannulated with polyethylene catheters (PE50) filled with heparin. The mean arterial pressure (MAP) and heart rate (HR) were recorded by a pressure transducer (Columbus Instruments) which was connected with the right femoral arterial catheter. HS was induced by withdrawing blood (through carotid artery, 2.5 mL/kg) in 10 min to keep MAP at 30 ± 5 mm Hg. The shock duration was maintained for 90 min either by further withdrawal of blood during the compensation phase or administration of the shed blood during the decompensation phase. Vehicle (PBS) or crocin (60 mg/kg) were then administered through intravenous injection (jugular vein) right before resuscitation. For resuscitation, the lactated Ringer's solution (1.5 mL/kg/h) was infused via the venous catheter to maintain the MAP for 4 h. For animals which survived the experiments (8 rats per group for sham+veh, sham+crocin, HS+veh, and HS+crocin), the MAP and HR were measured every 30 min throughout the procedures.
Organ injury assessment
The levels of urine creatinine and creatinine clearance were determined from the urine collected at the last 3 h of the resuscitation. Tissues and blood were harvested in the end of the procedures. Serum was then collected and assayed for creatinine, aspartate aminotransferase (AST), alanine aminotransferase (ALT), amylase, and lipase (IDEXX Laboratories). Lactic dehydrogenase (LDH), creatine phosphokinase (CPK), and lactate in the serum were also measured with an autoanalyzer (Roche Diagnostics, Penzberg, Upper Bavaria, Germany).
Western blot
Lung tissues were collected for Western blot analysis after resuscitation. To determine the subcellular localization of NF-κB signaling mediators, subcellular fractionation was performed. Tissues were first homogenized by a hypotonic buffer (250 mM sucrose, 10 mM KCl, 2 mM MgCl2, 1 mM EDTA, 20 mM HEPES pH = 7.4). Nuclear was pelleted by centrifuge at 500 g 5 min, followed by wash and finally lysed in extraction buffer. Supernatant was saved for cytoplasm fractions. The protein was quantitated by BCA assays, and the same amount of total protein lysates were run on sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE) followed by transfer to PVDF membranes. The membranes were blocked by 3% bovine serum albumin (Sigma, St. Louis, MO) and incubated with primary antibodies overnight at 4°C. Primary antibodies include p65, IκBα, p-IκBα, HDAC1 (Santa Cruz, Dallas, TX), and β-actin (Sigma). HRP-conjugated secondary antibodies were used to visualize bands under an electrogenerated chemiluminescence-based imaging system. Signals were analyzed in ImageJ and compared with controls after normalization (repeat 3 times).
Enzyme-linked immunosorbent assays
Serum cytokines tumor necrosis factor-α (TNF-α), interleukin (IL)-6, and IL-10 were determined using commercial immunoassay kits (R&D Systems, Minneapolis, MN) according to the manufacturer's instructions.
Statistics
All data in graphs are expressed as the means ± SDs (standard deviation) as indicated. The one-way or two-way analysis of variance followed by Bonferroni posttests was used. The comparisons with significance were denoted as *P < 0.05, **P < 0.01, and ***P < 0.001 compared with sham +vehicle control, or # P < 0.05, ## P < 0.01, and ### P < 0.001 compared with HS +vehicle control.
Results
Crocin administration does not affect the changes of MAP and HR in rats subjected to HS
Compared with sham groups, the MAP dropped rapidly after HS induced by blood withdrawal from the carotid artery catheter in rats. Even after 4-h resuscitation, MAP levels had recovered but remained lower in HS animals comparing with sham controls. Comparing with vehicle control, treatment of crocin (60 mg/kg, intravenous injection) did not cause significant effects on the MAP levels in both sham and HS groups (Fig. 1a). The dose of crocin used in the study was based on prior reports (Vakili and others 2014) and preliminary experiments (data not shown). Similarly, treatment of crocin had no effect on the HR in the experimental animals (Fig. 1b). The HR was significantly increased following HS, and no apparent differences were seen between vehicle and crocin administration in either sham- or HS-treated rats (Fig. 1b).

Effects of crocin treatment on
Crocin administration attenuates multiorgan dysfunction in rats subjected to HS
HS-related organ injuries were then assessed. First, to assess the extent of kidney dysfunction, we examined the levels of serum creatinine (Fig. 2a) and creatinine clearance (Fig. 2b) in the experimental groups. Compared with sham-operated rats, rats subjected to HS showed significant increases in serum creatinine and decreases in creatinine clearance (Fig. 2a, b). Notably, these signs of renal dysfunction were alleviated by crocin treatment. Second, the development of liver and pancreatic injuries in HS was indicated by the elevated plasma concentrations of AST (Fig. 2c) and ALT (Fig. 2d), as well as amylase (Fig. 2e) and lipase (Fig. 2f), respectively. Compared with vehicle control treatment, in HS-operated rats, crocin also greatly reduced such injuries (Fig. 2c–f). Finally, the plasma concentrations of LDH (Fig. 3a), CPK (Fig. 3b), and lactate (Fig. 3c) were measured. The increases of all these markers in HS groups suggested the occurrence of skeletal muscular injury and global tissue hypoxia. Administration of crocin in HS-operated animals resulted in an attenuation of the damages (Fig. 3a–c), compared with the vehicle treatment.
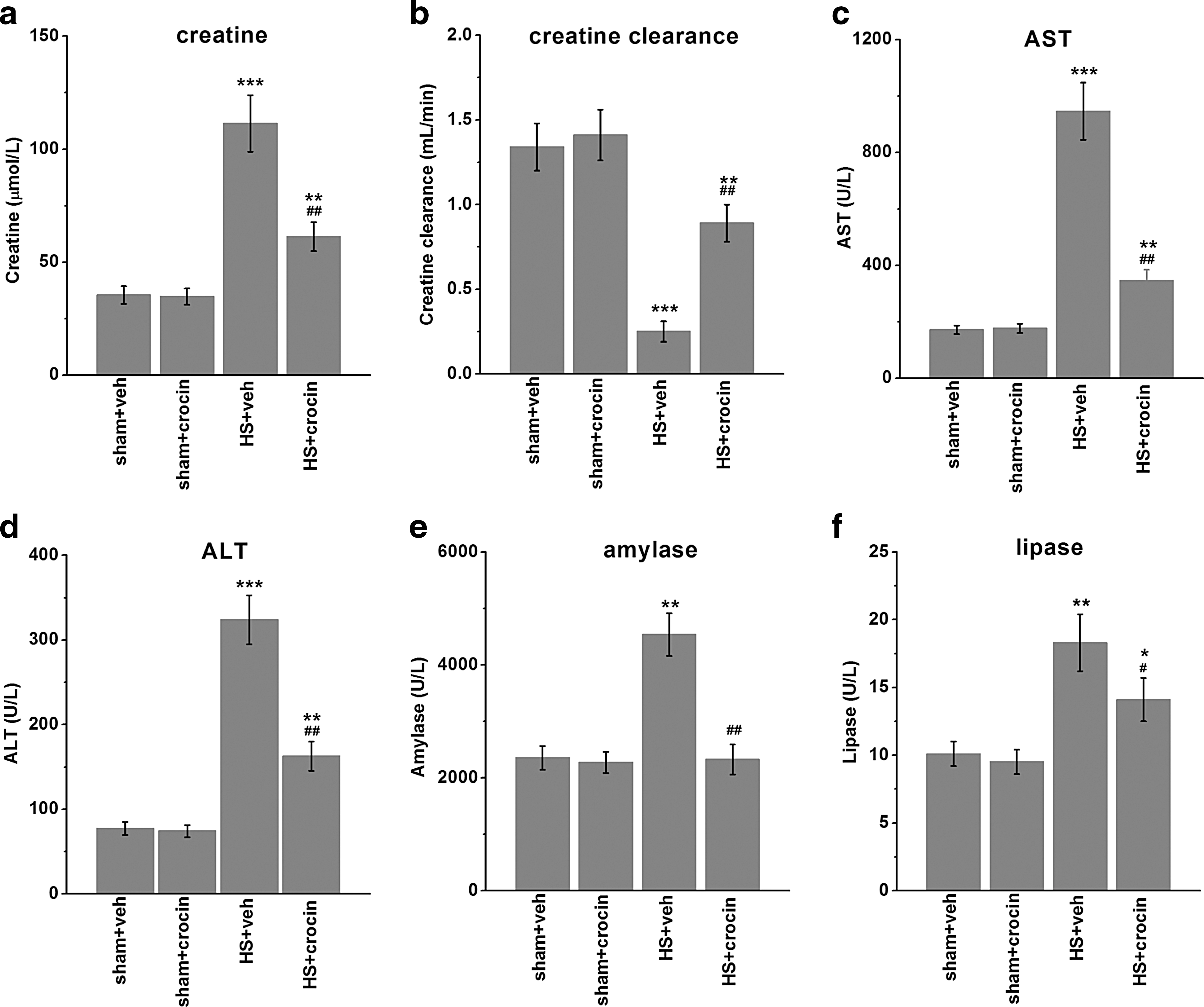
Effects of crocin treatment on multiple organ failure and dysfunction in HS-operated rats. Blood was collected and subjected to examination 4 h after resuscitation. The levels of creatinine
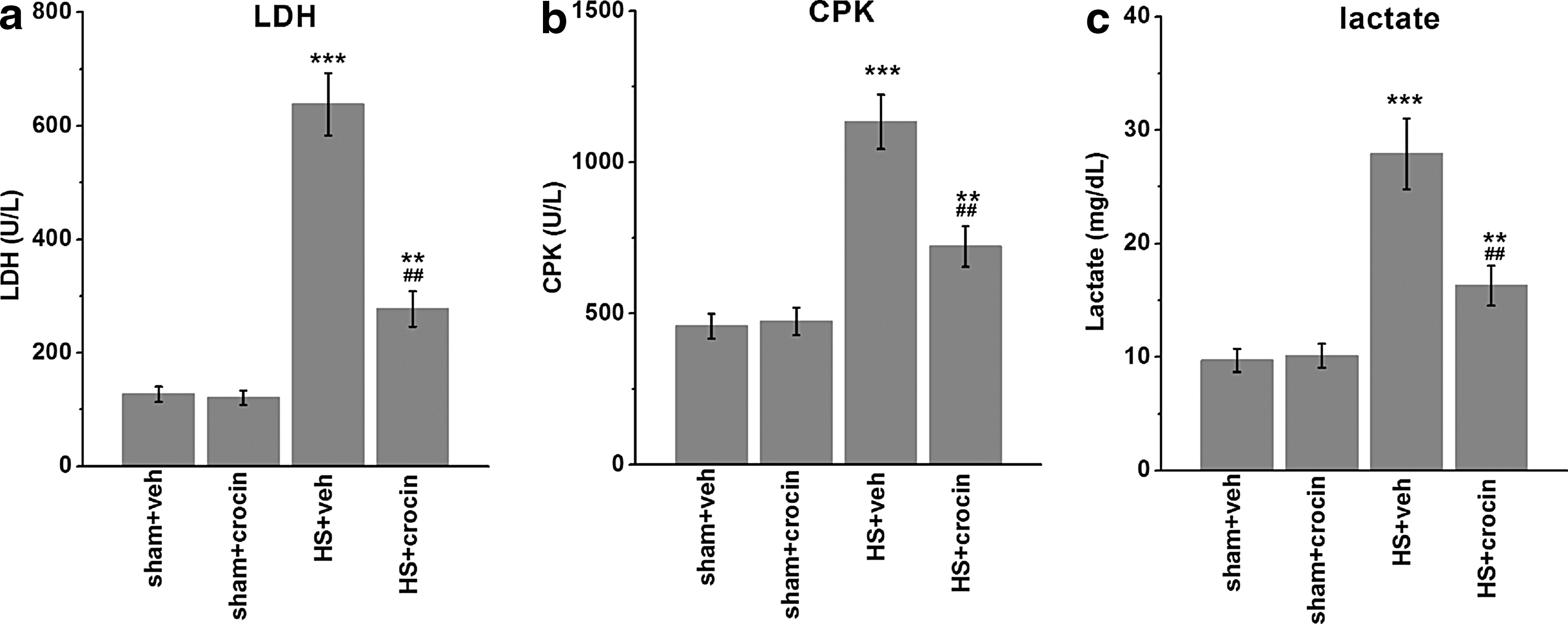
Effects of crocin treatment on serum LDH
Crocin treatment suppresses NF-κB activation in HS-operated rats
As NF-κB activation is a key player of HS-related organ injuries (Mauriz and others 2001; Guarini and others 2003), we examined NF-κB signaling in tissue level. Lung was selected for analysis because lung tissue is one of the earliest affected organs following HS, and reduced lung function is strongly associated with systemic inflammation in numerous inflammatory diseases. After resuscitation following HS, the nuclear translocation of p65 was dramatically induced, accompanied with an enhanced IκBα phosphorylation (Fig. 4a). Interestingly, total IκBα protein levels were unchanged in HS-operated rats (Fig. 4a, b), implicating a negative feedback loop, in which IκBα expression could be induced by sustained NF-κB activation (Covert and others 2005). Importantly, administration of crocin after HS significantly diminished the nuclear shuttling of p65 protein as well as the increased IκBα phosphorylation (Fig. 4a, b). These data suggest an inhibitory effect of crocin on NF-κB signaling activation in HS, which is likely a mechanism underlying the attenuated multiorgan injuries in these animals (Figs. 2 and 3).
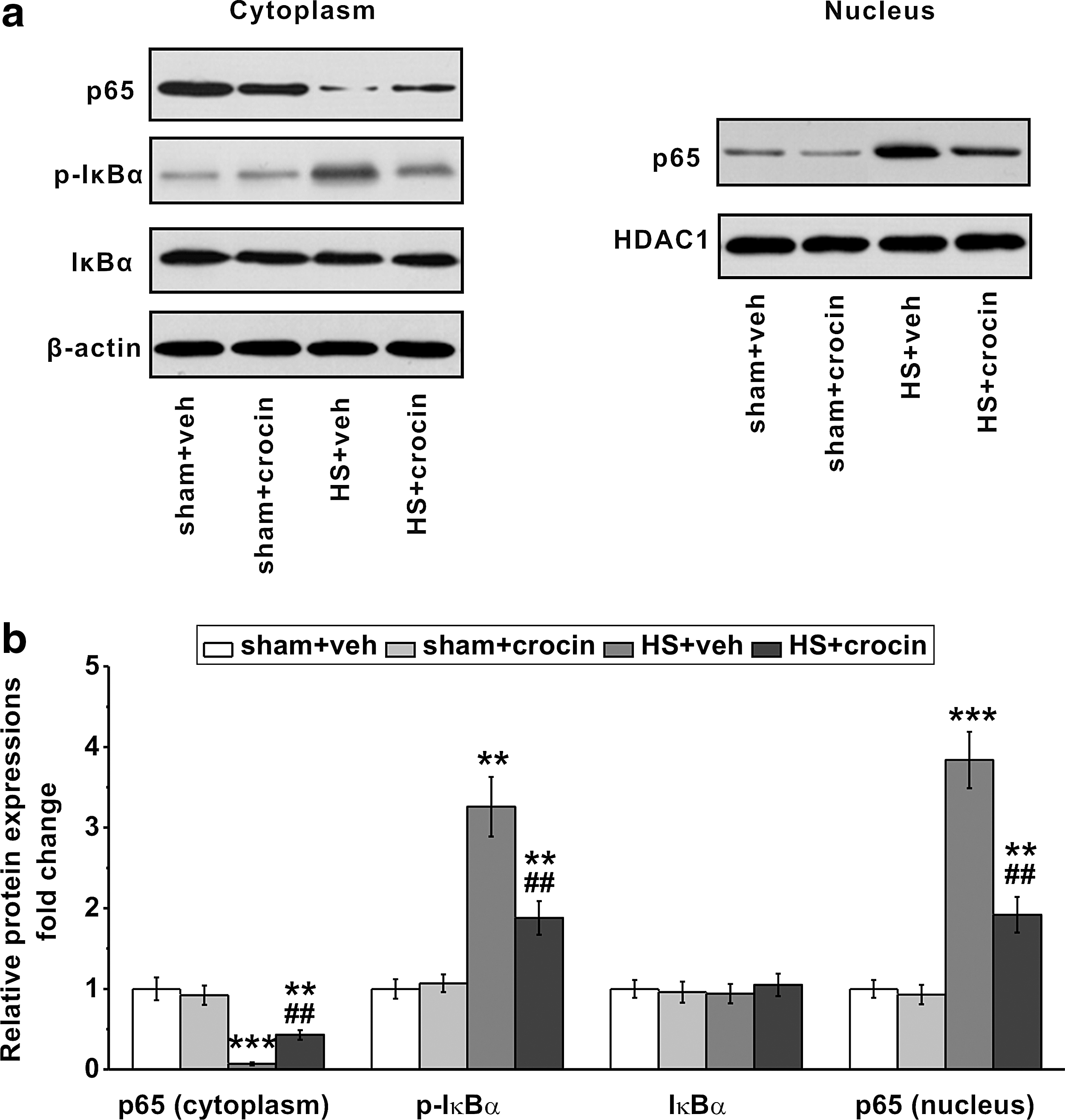
Crocin treatment suppresses NF-κB signaling pathway in HS-operated rats. Lung tissues were collected for Western blot analysis 4 h after resuscitation.
Crocin treatment reduces serum inflammatory markers in HS-operated rats
Since NF-κB signaling is essential for inflammation induction which is a major component of HS, we examined serum markers of systemic inflammatory response. As expected, the serum levels of proinflammatory cytokine TNF-α and IL-6 were greatly enhanced in HS + vehicle treated animals, which were significantly decreased by crocin administration (Fig. 5a, b). In contrast, the level of a key anti-inflammatory cytokine IL-10 was further raised by crocin in HS-operated rats (Fig. 5c).
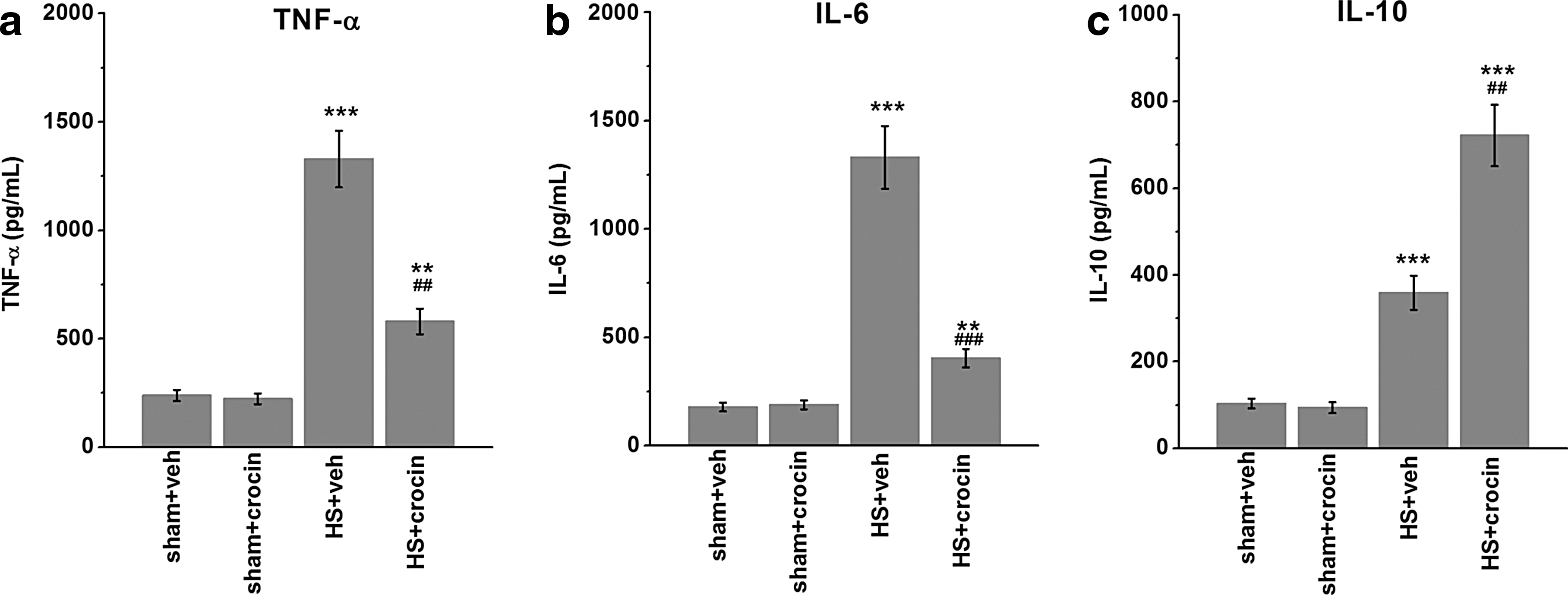
Crocin treatment attenuates serum inflammatory response in HS-operated rats. Serum levels of TNF-α
Discussion
Our findings suggest that intravenous administration of crocin to animals undergoing HS/resuscitation can attenuate or prevent the ensuing multiorgan dysfunction. Crocin treatment has significantly decreased the enhanced levels of creatinine and liver/pancreas enzymes, as well as diminished the induction of LDH, CPK, and lactate in the blood of HS rats. Thus, organ damage caused by HS appeared to be partially prohibited by the crocin administration. These effects are possibly through its anti-inflammatory properties. Indeed, HS-stimulated nuclear translocation of p65 protein and increased IκBα phosphorylation in the lungs were potently reduced in crocin-treated animals, further suggesting that the protective effect of crocin on HS might be driven by the suppression of NFκB signaling. Consistent with this model, our data found that the serum levels of proinflammatory cytokines induced in HS were significantly lower when treated by crocin, while anti-inflammatory cytokine IL-10 was increased. Together, these evidences describe a role of crocin in ameliorating HS-induced multiorgan injuries and suggest that there may be a causal relationship between the anti-inflammatory activity of crocin and the attenuated organ damages in HS-operated rats which received crocin treatment.
The potential role of crocin in HS may be hinted by a previously demonstrated function of crocetin, which is a close compound to crocin (Nam and others 2010; Yan and others 2010; Wang and others 2012). Magnetic resonance spectroscopy data have indicated that crocetin increased oxygen uptake by the muscle, which may be related to an increased diffusion of oxygen through plasma (Gainer and others 1993). Because a chemical shock causes a lack of cellular oxygen supply in tissues, enhancing oxygen extraction rate by crocetin may prevent shock responses, including organ damage. For example, Wang and others (2012) have reported that crocetin has beneficial effects on renal dysfunction caused by HS/resuscitation in rats; Dhar and others (2005) found that crocetin limits hepatocyte apoptosis and improves survival following HS. Beyond its effects on oxygen diffusion, crocetin has also been suggested by prior studies to inhibit inflammatory cascades by decreasing production of ROS, which may be correlated with its protection of oxidative damage during hemorrhage shock and resuscitation. Similarly as crocetin, our data in rats point to a therapeutic potential of crocin in HS. Crocetin is a known metabolite of crocin after oral administration. To specifically test the effects of crocin, we delivered crocin to animals directly through intravenous administration for a relatively short period of time. Nevertheless, we did not measure the levels of crocetin or crocin in the plasma to rule out the possible contribution of crocin metabolites, which can be largely affected by in vivo pharmacokinetics involving different formulations and application modes. The optimal dosage and timing thus still wait for a detailed investigation to determine the appropriate regimen of crocin application in future clinical trial. In addition, since crocin has additional sugar moieties, which play a critical part in its antioxidant behavior, we speculate that crocin should have a superior function than crocetin in its chemical reactivity toward buffering the severe consequences of HS. This hypothesis could be potentially addressed by side-by-side comparison between 2 compounds in future studies.
Our studies have demonstrated that crocin can decrease proinflammatory response caused by HS, which could be the underlying mechanism of the crocin action in ameliorating multiorgan injuries. First, the blockade of NFκB signaling by crocin may be correlated with a reduction of the early immediate response genes such as iNOS and COX-2, which facilitate the rapid production of NO and prostaglandins that initiate the lethal effects of HS/resuscitation (Hierholzer and Billiar 2001). Second, the downstream induction of inflammatory cytokines, which can play a catalyzing role for tissue damage during the resuscitation phase (Murao and others 2003; Ding and others 2014; Leung and others 2015), can be greatly attenuated by crocin via NFκB suppression as shown in our study. The current data in HS-operated animals agree with the prior observations in the cell culture that crocin contains a general anti-inflammatory property. For example, crocin in lipopolysaccharide (LPS)-challenged macrophages was found to significantly decrease the productions of proinflammatory mediator PGE2 (prostaglandin E2), along with an inhibition of NFκB activation (Xu and others 2009). Similarly, crocin effectively reduces the release of NO, TNF-α, and IL-1β from cultured rat brain microglial cells stimulated by LPS (Nam and others 2010). All these data, including ours, indicate that crocin compound exhibits an obvious anti-inflammatory activity that can be potentially utilized to modulate inflammatory processes such as HS.
Alternatively, anti-inflammatory effects of crocin in HS could be secondary to the inhibition of HS-induced organ dysfunction. This is because the tissue damage itself during the ischemic phase of shock often initiates a proinflammatory response, including the activation of NFκB pathway and its downstream target expressions (Covert and others 2005). Our current model clearly shows that the anti-inflammatory function of crocin is associated with, or most likely participates, in the protection of HS-induced organ injuries. However, it is important to note that these experiments cannot test the causality. That is, crocin may also directly act on HS-induced organ damage, which is primarily independent of inflammation. Specifically, it is well known that oxidative stress-mediated direct toxicity or ROS-induced inflammatory cascades contribute to organ injury. Since crocin is one of the active antioxidant ingredients in Gardenia jasminoides Ellis or Crocus sativus L, a leading mechanism for the protective role of crocin on HS-induced organ injuries may well reside in its antioxidative functions (Zheng and others 2007). As the increased oxidative stress and systemic inflammatory response are often tightly connected to each other, it is hard to dissect out the primary cause. It is very likely that the anti-inflammatory and antioxidant effects of crocin interact and cooperate in the protection of HS-induced organ injuries. Therefore, the evaluation of markers for oxidative stress in our model should provide us, at the least to some degree, a more complete picture of the contribution of crocin in HS. Other factors which might account for these results independent of anti-inflammatory functions of crocin (Hosseinzadeh and others 2014) will also be interesting to study by future investigation. For example, in a most recent study by Zeng and others (2016), crocin significantly promoted autophagy during ischemia, while inhibited autophagy during reperfusion of neonatal mouse cardiomyocytes in vitro. Since activation of autophagy pathway has been shown as a protective response to IR injury (Hamacher-Brady and others 2006), enhancing autophagy by crocin may be responsible for its myocardial protective effect. Similarly, such autophagy-mediated mechanisms could explain the crocin effects we observed in HS-induced organ damage, in which toxic waste products build up, such as incomplete removal of mitochondria (Zhang and others 2013). As such, whether or not crocin can modulate autophagy activity during HS needs to be examined.
Taken together, intravenous crocin administration in the current study suggests a protective function of crocin against HS-induced multiorgan injuries. It is possible that these effects are driven by an anti-inflammatory property of crocin, which could lead to a potentially successful treatment of patient with HS by natural antioxidants.
Footnotes
Author Disclosure Statement
No competing financial interests exist.