
Abstract
Follistatin-like protein-1 (FSTL1) is an inflammatory factor that can induce an inflammatory response and is expressed in cancers. However, little is known about its content and function in nasopharyngeal carcinoma (NPC). Interleukin (IL)-12 and IL-4 are primarily secreted by dendritic cells (DCs) and activated T lymphocytes, respectively; these factors can induce Th cell differentiation and cytotoxic lymphocyte production, both of which facilitate tumors through the STAT4 and STAT6 pathways, respectively. In this study, the relationship between FSTL1 and both IL-12 and IL-4 as well as the functional mechanism of these cytokines was explored. Enzyme-linked immunosorbent assay, flow cytometry, and Western blotting were used to assess the levels of key inflammatory factors and DC markers as well as elucidate the mechanism by which FSTL-1 mediates and exerts it antitumor effects. The results revealed that serum FSTL1 and IL-12 levels were significantly decreased in NPC patients compared with those in the control group (P < 0.05); conversely, IL-4 levels were increased (P < 0.05). Supernatants from the experimental groups (EGs) contained higher IL-4 and IL-12 levels than those from the control groups (P < 0.05). Additionally, phosphorylated-STAT6 and phosphorylated-STAT4 were increased in the EGs (P < 0.05). These results suggest that DC-mediated immunity was activated by FSTL1, which leads to an increase of IL-12 and IL-4 production and consequently activates the STAT4 and STAT6 pathways through upregulation of STAT4 and STAT6 phosphorylation, respectively.
Introduction
T
Immature dendritic cells (DCs) consume antigens, whereas mature DCs express costimulatory molecules and secrete cytokines that convey immune information to other immunocytes. First, T cells can be activated by costimulatory molecules on DCs, including CD40, CD86, and HLA-DR, to initiate a cytotoxic T lymphocyte (CTL) response (Guo and others 2003). Then, DCs are the primary source of interleukin (IL)-12 in the early immune response. IL-12 has been proven as having the strongest ability to induce T lymphocytes (particularly activated T cells and NK cells), to produce interferon (IFN)-γ, and to prevent EBV infection and tumor cell metastasis (Lesinski and others 2004; Sarhan and others 2015). The primary mechanisms by which IL-12 participates in immune regulation is the differentiation of Th naïve (Th) cells into Th1 cells and the promotion of Th1 growth and proliferation through the STAT4 pathway. Researchers found that IL-12-mediated p35 expression silenced DC-inhibited tyrosine phosphorylation of STAT4 in T cells and inhibited IL-12-induced activation of JAK2 in T cells (Xu and others 2007). As a result, translocation of STAT proteins to the nucleus is blocked and they cannot mediate the transcription of many IL-12 response genes associated with proliferation, cytokine production, and cytotoxic activity of T cells. Furthermore, DCs are necessary for mediating an immune response and generating IL-4 from T lymphocytes. IL-4 can concurrently promote proliferation of DCs/cytokine-induced killer cells and upregulate the proportion of tumor-killing cells through IFN-γ (Zhao and others 2016; Yin and others 2016). IL-4 upregulates the differentiation of CTLs and lymphokine-activated killer cells (Puri and others 1994; Fabricius and others 2011). The most important function of IL-4 is that it promotes Th0 cell differentiation into Th2 cells as well as their growth and proliferation.
Notably, the occurrence, development, and immune escape of NPC were involved in an imbalance of Th1/Th2 cells. When the concentrations of Th1 cytokines [IL-2, tumor necrosis factor (TNF)-α, and IFN-γ] and Th2 cytokines (IL-4, IL-5, and IL-10) in the serum of 207 NPC samples were studied, the results indicated that serum Th1 cytokine levels were much lower in NPC patients than in control subjects, whereas Th2 cytokine levels were higher in NPC (Weifeng and others 2005). These results were replicated and suggest that either lower serum IL-2 levels or serum TNF-α concentrations predict an unfavorable prognosis for patients with NPC (Lu and others 2012). These results also demonstrate the existence of an imbalance of Th1/Th2 cells in NPC patients and that IL-12 and IL-4 play important roles in regulating the Th1/Th2 ratio.
Follistatin-like protein-1 (FSTL1) is an extracellular matrix glycoprotein that participates in the immune response by regulating ILs, IFNs, and other immune molecules through signaling pathways. For example, FSTL1 was previously shown to induce the production of ILs in macrophages and fibroblasts through the Toll-like receptor 4/nuclear factor kappa b (NF-κB) signaling pathway (Whiteside 2006). FSTL1 promotes the cellular immune response in the synovium of patients with rheumatoid arthritis by enhancing the function of antigen-presenting cells (APCs) surrounding the synovium (Chaly and others 2014). FSTL1 secretion from fibroblasts increases the secretion of TNF-α and IL-1β from macrophages, and injecting an adenoviral vector encoding FSTL1 into mice induces excessive IL-6, IL-1β, and TNF-α expression in the liver (Miyamae and others 2006). In vitro, FSTL1 activates the electron transport chain in mitochondria, which consequently increases ATP production of ATP and IL-1β secretion from monocytes/macrophages (Chaly and others 2014). These results suggest that FSTL1 is highly associated with immunocytes. Importantly, our preliminary experiments confirmed that FSTL1 activated DCs in NPC patients and increased IFN-γ secretion (Wang and others 2016). Exogenous FSTL1 regulates the immune response by enhancing the antigen presentation ability of APCs and the cytokine secretion ability of immunocytes. However, few publications have explored the relationship between FSTL1 and either IL-4 or IL-12.
In addition, the JAK-STAT4/STAT6 signaling pathways have been demonstrated to be critical for the action of IL-4 and IL-12 on lymphocyte differentiation and proliferation (Chaly and others 2012; Chapoval and others 2010; Stark and others 2013; Weaver and others 2015). IL-12 induces the development and differentiation of Th1 cells, and STAT4 phosphorylation induces the transient production of IFN-γ by Th1 cells (Berenson and others 2006). Additionally, IL-4 signaling through IL-4 receptors on naïve CD4+ T cells leads to the phosphorylation and activation of STAT6, which has been shown to mediate the differentiation and proliferation of T lymphocytes into Th2 effectors in vitro (Shimoda and others 1996; Kelly-Welch and others 2003). Therefore, we hypothesized that FSTL1 could affect the function of DCs by influencing IL-4 and IL-12 secretion, which exert their activity through the STAT6 and STAT4 pathways to affect the Th1/Th2 balance in NPC patients.
Materials and Methods
Specimens
From August 2013 to May 2015, 167 NPC cases were confirmed in patients diagnosed with undifferentiated-type noncornification NPC by the Pathology Department of the First Affiliated Hospital of Guangxi Medical University. These patients included 126 males and 41 females and had an average age of 47 ± 11.26 years. The control group consisted of 140 non-NPC patients, including 76 males and 64 females, with an average age of 44.39 ± 15.53 years. All subjects were recruited from the outpatient Otorhinolaryngology Head and Neck Surgery Department (ENT & HN Surgery Department).
Separation of peripheral blood mononuclear cells and induction of DCs
Before blood collection, the participants were clearly informed such that they understood the study and provided their written informed consent. The project was approved by the ethics committee of the First Affiliated Hospital of Guangxi Medical University. Twenty milliliters of blood were collected from each subject in EDTA-anticoagulant tubes, and 5 mL of coagulant blood was collected. The peripheral blood mononuclear cells (PBMCs) were separated using Lymphoprep™ (STEMCELL™ Technologies) according to the manufacturer's instructions. Approximately 1.3 × 106 to 3.2 × 106 PBMCs/mL were cultured in 6-well flat-bottom plates in RPMI 1640 medium (HyClone; Thermo Fisher Scientific) supplemented with 10% fetal bovine serum and 1% penicillin–streptomycin at 37°C 5% CO2. Four hours later, the adhesion method was used to separate the lymphocytes and PBMCs, and the lymphocytes were stored at −80°C. Recombinant human granulocyte–macrophage colony-stimulating factor (1,000 U/mL; American PeproTech, Inc.), recombinant human IL (rhIL)-4 (500 U/mL; American PeproTech, Inc.), and recombinant human TNF-α (1,000 U/mL; American PeproTech, Inc.) were used to induce the DCs. The culture medium was refreshed every 3 days with fresh RPMI 1640 medium containing growth factors, and the cultures were split upon reaching confluence. The DCs were harvested on the eighth day of culture.
Detection of cell surface molecule expression by flow cytometry
On the third, fifth, and eighth days, CD40, CD86, and HLA-DR expression was examined on the DC surfaces by flow cytometry to judge the maturity of DCs. The cells were labeled with antibodies against the above surface markers by resuspending 1 × 105 cells in 100 μL of 1 × phosphate buffer saline containing 5 μL of antibody; additionally, 5 μL of the isotype control was used as a control (FITC-conjugated mouse IgG1K isotype control). The flow cytometry analysis was performed on an FACSAria (BD Biosciences) instrument.
Experimental groupings
Mature DCs were divided into the following 4 groups: the NPC-experimental group (EG) (cells + FSTL1), the NPC-control group (CON1), the control EG (CON2) (cells + FSTL1), and the control–control group (CON3). FSTL1 (American R&D, Inc.) was added to a final concentration of 40 ng/mL. After 24 h, cell culture supernatants were discarded, and the cells were stimulated with LMP2 (PepMix-EBV-LMP2; JPT Peptide Technologies GmbH, Berlin, Germany), which is a peptide fragment with antigenic characteristics of EBV. PepMix-EBV-LMP2 (497aa) is an antigen peptide, derived from the B95-8 cell line, containing 122 peptides with EBV-specific antigen signals. It does not contain complete EBV particles or other antigen information. The cells were then cocultured with lymphocytes from the same NPC patients and healthy donors.
DC and lymphocyte coculture
The ratio of the 2 cell types was ∼1:8. During coculture, rhIL-7 (American PeproTech, Inc.) was added to a final concentration of 10 mg/mL. Half of the medium was changed every 3 days, and 20 mg/mL IL-7 was added to all groups. On the seventh day, new medium containing IL-7 (20 mg/mL) and IL-2 (40 U/mL; American PeproTech, Inc.) was added to all groups, which were cocultured in a 5% CO2 incubator at 37°C. Finally, all of the cells and cultures were harvested on the 15th day.
Detection of FSTL1, IL-4, and IL-12 expression by enzyme-linked immunosorbent assay
After thawing on ice for 40 min, all serum samples and supernatants were evaluated using Human FSTL1, Human IL-4, and Human IL-12 Sunny ELISA kits [MultiSciences (Lianke) Biotech Co., Ltd., Hangzhou, China] following the manufacturer's instructions. The Curve Expert 1.4 software was used to analyze the OD values and create the standard curve (r > 0.99).
Detection of p-STAT4 and p-STAT6 expression by Western blotting
The total proteins from the cells of each group were extracted. Western blotting was performed to detect the expression of phospho-STAT4 [phospho-STAT4 (Tyr693) (D2E4) rabbit mAb, CST], phospho-STAT6 [phospho-STAT6 (Tyr641) antibody, CST], and β-actin [β-actin (14C10) rabbit mAb, CST]. The primary antibodies were diluted with Western blotting primary antibody diluent to 1:1,000, and the secondary antibody [anti-rabbit IgG (H + L) (DyLight 680 conjugate), CST] was diluted in 1 × Tris-buffered saline with Tween-20 (TBST, pH 8.0) to 1:15,000.
Data analysis
The data were analyzed using SPSS 17.0 and GraphPad Prism v. 5 software. The data are shown as the mean ± standard error of the mean from at least 3 independent experiments. Significant differences were determined using the 2-tailed unpaired T-test. A P-value less than 0.05 was considered significant.
Results
FSTL1 and IL-12 levels in NPC patients' serum are downregulated, while IL-4 level is upregulated
We evaluated the serum levels of FSTL1, IL-12, and IL-4 in NPC patients and control subjects. As shown in Fig. 1, the FSTL1 and IL-12 expression levels were markedly lower in NPC patients than in control subjects, whereas the IL-4 expression levels were significantly higher in NPC patients. Interestingly, FSTL1 levels in NPC patients were ∼3 times lower than those of the control subjects, and there was a significant decrease when NPC occurs (P < 0.05). Similarly, IL-12 levels of NPC patients were also decreased, but not to the extent as FSTL1—there was only a 40 pg/mL difference between the NPC and control groups (P < 0.05). However, the serum level of IL-4 in NPC patients was much higher than that of the control subjects (90 and 55 ng/mL, respectively); this difference was significant (P < 0.05).

The 3 bar charts in this figure show the serum levels of FSTL1, IL-12, and IL-4 as measured by ELISA. The data are presented as the mean ± SEM of duplicates, and these results are representative of at least 3 independent experiments.
Induced DCs from NPC patients and control groups have the same maturity
To reveal the detailed maturity of DCs, we analyzed the percentage of DC surface markers. In Fig. 2, to observe the maturity of DCs induced from PBMCs from NPC patients and control subjects, we measured CD40, CD86, and HLA-DR expression on the surface of DCs using flow cytometry. The line graphs (Fig. 2A, B) show that the expression of these 3 cell surface molecules dramatically increased. During the induction period, CD40 expression increased from ∼30% (the third day) to ∼80% (the eighth day). HLA-DR increased from ∼20% (the third day) to ∼85% (the eighth day), and CD86 increased from 30% to 96% (Fig. 2C, D). There were significant upward trends when DCs were cultured and induced by cytokines (P < 0.05). Nevertheless, upon comparison of the difference in the percentage of DC surface markers between the NPC patient and control groups, there was little difference (P > 0.05). The results suggest that the DCs matured during the induction and cultivation periods and that DCs from both NPC patients and control subjects had matured without any significant differences between the 2 groups.
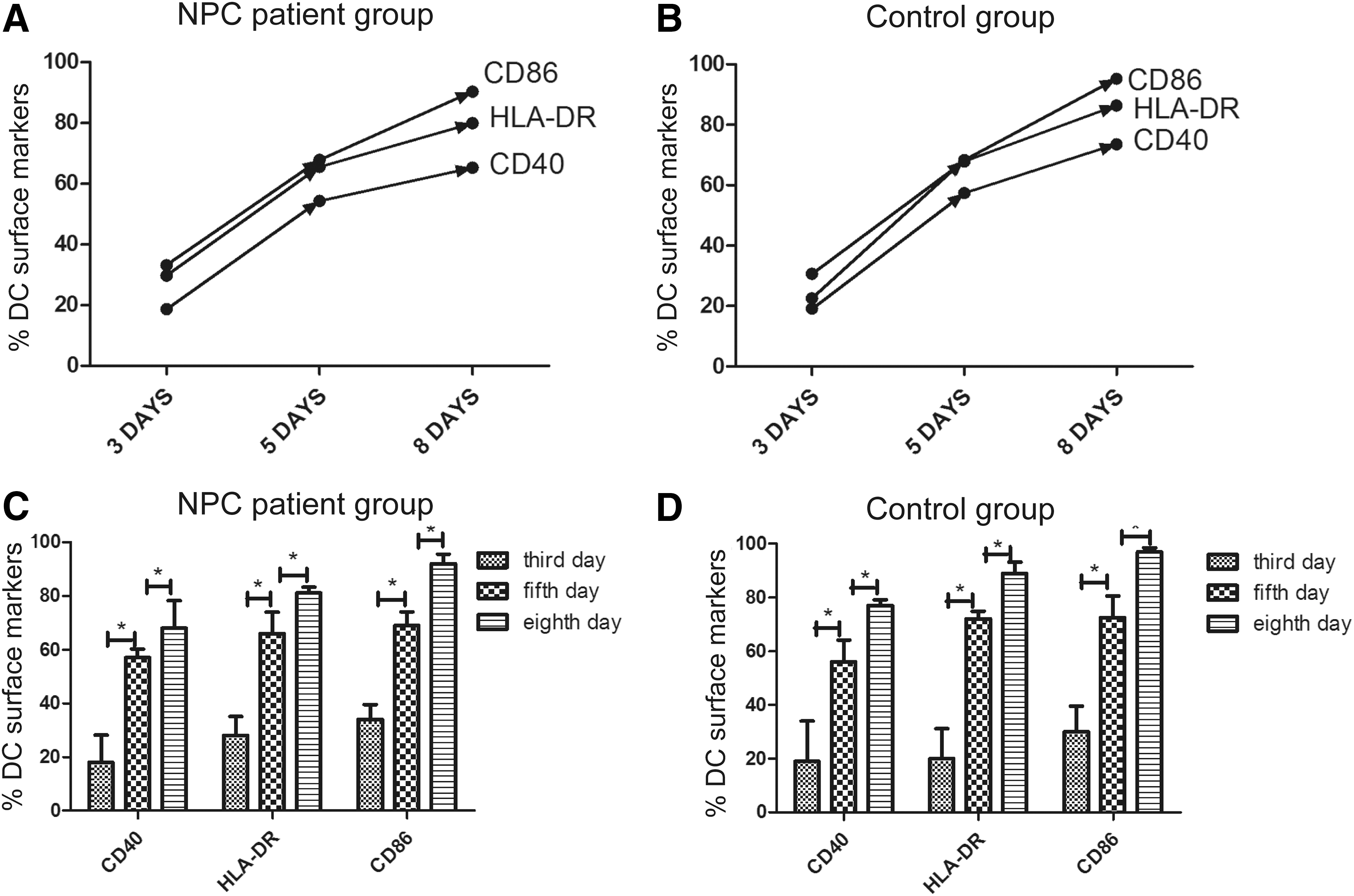
The images in this figure illustrate the relative expression of DC surface markers—CD40, CD86, and HLA-DR—in the NPC patient and control groups.
Exposure to recombinant FSTL1 protein promoted the generation of IL-4 and IL-12
As FSTL1 is a secreted protein, it may function in an extracellular environment. We treated matured DCs with recombinant FSTL1 protein at 40 ng/mL and detected IL-4 and IL-12 levels in cell supernatants of cocultured DCs and lymphocytes. The results illustrate that extracellular FSTL1 upregulates the generation of IL-12 in NPC-EG and healthy subjects. However, FSTL-1 does not regulate IL-4 of control subjects, although it advances that of an NPC patient. Figure 3A and B shows the IL-4 and IL-12 levels in the experimental and control groups. The bar chart in Fig. 3A indicates that IL-4 expression was markedly higher in the cell culture supernatants from NPC-EG cells than in those from CON1 cells, which demonstrated that FSTL1 treatment enhanced the antigen presentation ability of DCs and promoted lymphocyte activity to generate more IL-4. However, this type of upregulation has no effect on healthy people, as indicated by the CON2 and CON3 groups. Moreover, in comparison with CON2 cells, which were activated in response to FSTL1 in a similar manner as NPC-EG cells, NPC-EG cells most likely had lower IL-4 levels because CON2 cells had a higher baseline level than NPC-EG. Therefore, we emphasize that FSTL1 can improve the immunity of DCs from NPC patients, but not from healthy donors in terms of IL-4 generation. Additionally, in Fig. 3B, measurement of IL-12 levels in the cell culture supernatants revealed that CON2 cells had the highest levels and that CON1 cells had the lowest levels. The higher IL-12 levels in culture supernatants from NPC-EG cells compared with those from CON1 cells showed that FSTL1 enhanced IL-12 secretion. Interestingly, the IL-12 levels were markedly higher in CON2 cells than in CON3 cells, which indicated that FSTL1 had functions in the generation of IL-12 in healthy humans.
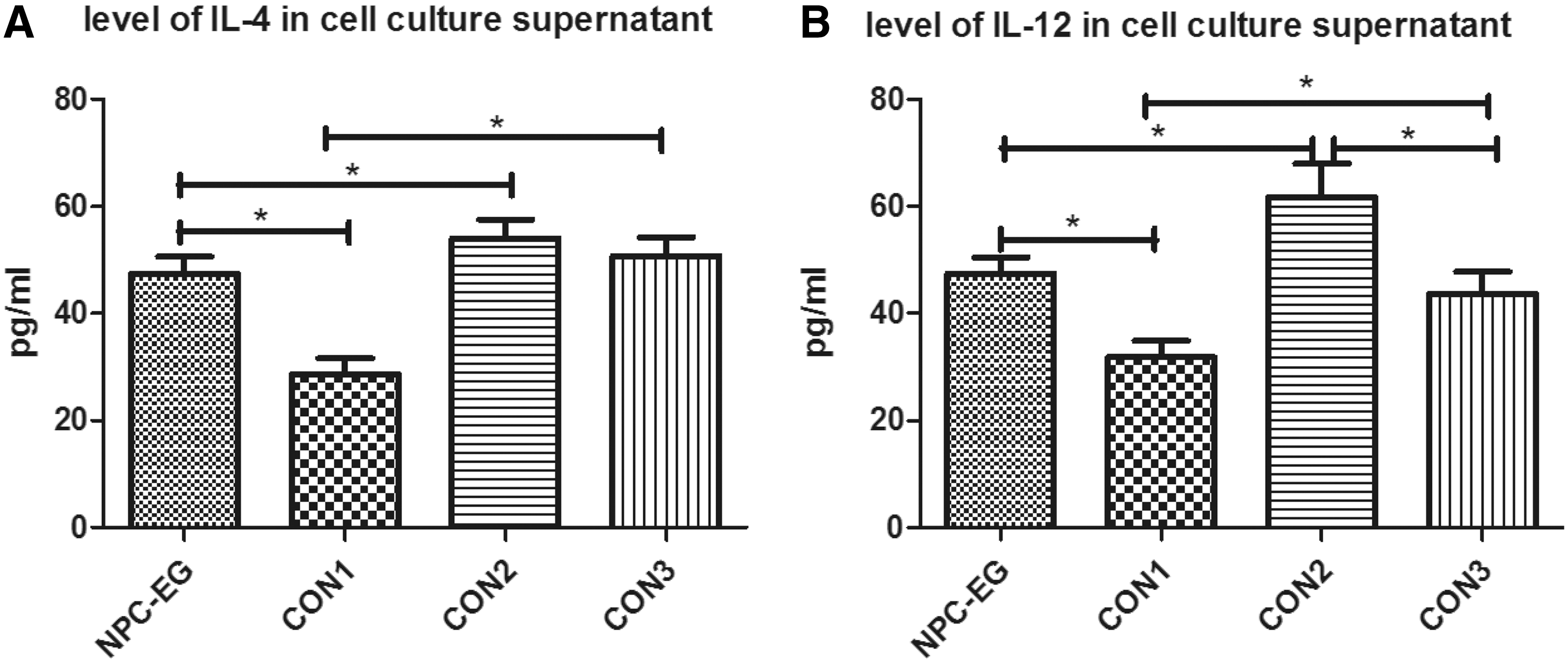
This bar chart shows increases in IL-4 and IL-12 expression in NPC-EG cells.
The increases of p-STAT4 and p-STAT6 are the mechanisms underlying functions of IL-12 and IL-4, respectively
To explore the mechanisms underlying the functions of IL-4 and IL-12, the levels of p-STAT4 and p-STAT6 were measured by Western blotting as shown in Fig. 4. Higher protein levels of p-STAT6 (110 kDa) and p-STAT4 (81 kDa) were detected in NPC-EG, CON2, and CON3. After adjusting for β-actin (45 kDa) expression, the relative expression values of these 2 proteins are shown in Fig. 4B and C. The p-STAT4 and p-STAT6 levels were significantly upregulated in NPC-EG cells compared with CON1 cells (P < 0.05). The highest p-STAT4 expression levels were detected in the CON2 group, whereas the lowest expression levels were found in the CON1 group. Similarly, p-STAT6 expression was also highest in the CON2 group and lowest in the CON1 group. However, the differences between CON2 and CON3 were not significant. These results illustrate that FSTL1 activated IL-12 and IL-4 secretion from DCs and lymphocytes, respectively, from NPC patients; these secreted molecules promoted STAT4 and STAT6 phosphorylation and functioned as immune activators in vitro.

Discussion
According to our results, the downward FSTL1 level in NPC patient serum could be considered a biomarker of NPC. FSTL1 is a tumor-inhibiting factor that has been found to be downregulated in cancers. In the process of researching 45 endometrial and 36 ovarian cancers, reduced FSTL1 expression was identified. They cloned the full-length cDNA of FSTL1 and transiently transfected it into ovarian cancer and endometrial cell lines; the results suggested that migration and invasive abilities of these cell lines were reduced by FSTL1 (Chan and others 2009). When clear cell renal cell carcinoma samples from 84 patients were cultured in vitro and in vivo, the FSTL1 gene was observed to be downregulated (Tan and others 2008). Furthermore, in NPC cell lines and primary tumor biopsies, FSTL1 was frequently downregulated owing to promoter hypermethylation. The tumorigenicity of NPC cells was also decreased by overexpression of FSTL1 in vivo. In addition, the colony-forming, proliferative, migratory, and invasive abilities of NPC cells were obviously inhibited by ectopic expression of FSTL1, which induced NPC cell apoptosis. Interestingly, after NPC cells were treated with soluble recombinant FSTL1 protein, their in vitro proliferation was significantly suppressed (Zhou and others 2016). In this article, when we compared the serum FSTL1 levels in NPC patients (n = 167) and non-NPC patients (n = 140), we observed lower FSTL1 expression levels in serum samples from NPC patients than in serum samples from the control groups (Fig. 1). Combined with previous studies, decreased serum FSTL1 expression could be considered a marker of NPC occurrence.
According to IL-12 and IL-4 levels in serum, we thought that the Th1/Th2 balance in NPC patients was disrupted. In this article, we found that there were much lower levels of serum IL-12 in NPC patients than in control subjects, whereas the serum IL-4 levels of NPC patients were higher than those of control subjects (Fig. 1). In fact, IL-12 is generated by DCs, macrophages, monocytes, neutrophils, and B cells in vivo; additionally, it induces the proliferation and differentiation of Th cells to Th1 phenotype; promotes the production of IFN-γ, IL-2, and TNF-α by T cells and NK cells, and enhances CTL activity (Sarhan and others 2015). Based on the observed reduction of serum IL-12 levels in NPC patients, we speculated that there was a decline in the percentage of Th1 cells. However, there was an obvious increase of IL-4 levels in the serum samples from NPC patients compared with the levels in the control group; IL-4 is secreted by activated lymphocytes, B cells, and NKT cells and is the key factor for the differentiation of Th0 cells into Th2 cells. Thus, an increase in the number of Th2 cells can also be speculated. Indeed, the balance of Th1 and Th2 cells plays a critical role in regulating NPC immune escape and EBV infection (Kidd 2003). Researchers found that in NPC patients, the balance of Th1/Th2 shifted to Th2, and this type of shift would correspond to an advanced TMN grade of NPC (Xiang and Yin 2009). Our speculation was in agreement with these study results—the Th1/Th2 balance in NPC patients was disrupted, and one underlying mechanism was the changes of IL-12 and IL-4 levels, which were affected by decline of FSTL1 expression.
FSTL1 is a soluble secreted protein that induces host immune cell function. Interestingly, FSTL1 had been observed to increase the synthesis of proinflammatory cytokines and chemokines in immune cells both in vitro and in vivo (Kawabata and others 2004). Our previous studies demonstrated that FSTL1 improved the antigen-presenting ability of DCs and enhanced immunity of T lymphocytes through the NF-κB pathway, which causes an increase of IFN-γ expression (Wang and others 2016). Researchers found that during early stages of arthritis, FSTL1 is highly overexpressed in mouse paws, especially at the interface of the synovial pannus and eroding bone. Transfecting the FSTL1 gene into macrophages and fibroblasts resulted in the upregulation of proinflammatory cytokines, including IL-1β, TNF-α, and IL-6 (Sumitomo and others 2000). Similarly, decreased FSTL1 protein levels in NPC patients were associated with downregulated IL-1β and TNF-α expression. IL-1β and TNF-α production in in vitro macrophage cultures was induced by exogenous FSTL1 protein expression (Chaly and others 2014). Therefore, FSTL1 might activate macrophages and attenuate the immune evasion of NPC cells (Zhou and others 2016). In this article, mature DCs from NPC patients and non-NPC donors (as indicated by surface marker expression in Fig. 2), following been stimulated by FSTL1 and activated by LMP2-MIX antigen peptide fragment under identical conditions, showed significant differences in IL-12 and IL-4 expression. Figure 3 shows that IL-12 and IL-4 expression in NPC-EG cells was much higher than that in the control groups, and the difference in IL-12 and IL-4 levels in the supernatants gradually shrank. If the IL-12 and IL-4 levels accurately represent the respective number of Th1 and Th2 cells, the difference between IL-12 and IL-4 expression can indirectly indicate an upward trend of the Th1/Th2 ratio, which suggests that FSTL1 participates in regulating the Th1/Th2 balance.
The JAK-STAT4/STAT6 pathway is critical for the differentiation and proliferation of lymphocytes. Although one JAK kinase can participate in a variety of cytokine signal transduction processes, there is a certain selectivity as to which cytokines activate the STAT molecules. For example, IL-4 activates STAT6, whereas IL-12 activates STAT4; this is because the IL-12 and IL-4 receptors corresponding to their respective STAT pathways are expressed on the surface of naïve T cells and Th2 cells, respectively (Hallett and others 2012; Chistiakov and others 2015). IL-12 and STAT4-mediated signaling are the major instructive cytokine and pathway responsible for increasing CD40 L expression in CD8+ T cells, which forms a self-sustaining positive feedback loop with TCR signaling to facilitate the efficient priming of T cells stimulated by APCs (Stark and others 2013). IL-4 and its downstream transcription factor STAT6 modulate the differentiation of Th2 cells and promote inflammation (Maier and others 2011). To explore the mechanisms underlying the changes in IL-4 and IL-12 production, p-STAT4 and p-STAT6 expression was assessed by Western blotting as shown in Fig. 4. The p-STAT6 and p-STAT4 expression levels were upregulated in NPC-EG and CON2 cells after FSTL1 activation. Notably, the Western blotting results were relatively consistent with the IL-4 and IL-12 secretion results. Our results showed that FSTL1 increased IL-12 and IL-4 production and they promoted STAT4 and STAT6 phosphorylation in Th1 and Th2 cells in vitro, respectively. The mechanism has been shown in Fig. 5.

This picture shows the mechanism behind this article. Naive T cell is activated by mature DC, which has been promoted by FSTL1 and generates IL-12. Activated T cells differentiate into Th1 and Th2 cells, which are promoted by IL-12 and IL-4 separately. On the Th1 cell surface, IL-12 conjuncts to IL-12 receptors, advancing phosphorylation of STAT4 and further increasing generation of IFN-γ and TNF. STAT6 phosphorylation is from IL-4 combining with IL-4 receptor α on the Th2 cell surface. These are benefits to IL-5, IL-6, and IL-10 production. IFN, interferon; TNF, tumor necrosis factor.
To sum up, our results show that the addition of recombinant FSTL1 protein advances the immunity of DCs from NPC patients, by increasing IL-12 secretion, and enhances the antigen presentation ability of DCs. In addition, FSTL1 closes the gap between IL-12 and IL-4 expression to regulate the Th1/Th2 ratio, and the upregulation of STAT4 and STAT6 phosphorylation is a critical underlying mechanism. Taken together, these make FSTL1 an ideal immunotherapeutic method especially suitable to be developed as an anticancer immunity drug. However, further study is necessary to explore this potential.
Footnotes
Acknowledgments
The authors thank Zhe Zhang and Zhiwen Xu (the First Affiliated Hospital of Guangxi Medical University) for their comments. They also thank the staff at the ENT outpatient clinic of the First Affiliated Hospital of Guangxi Medical University for their assistance with specimen collection. The authors also express their heartfelt thanks to the teachers and students in the experimental center of Guangxi Medical University for their experimental comments. This work was funded by the Nature Science Foundation of Guangxi (No. [2015]139) and the Innovation Project of Guangxi Graduate Education (YCBZ2017038).
Author Disclosure Statement
No competing financial interests exist.