
Abstract
Although immune dysfunction by space conditions has been reported postflight, as well as during ground-based experiments, the cause(s) and nature of the immunological changes are not completely understood. Microgravity has been suggested as one of the factors responsible for the observed immune dysregulation. The goal of this study was to assess immune changes in simulated microgravity (s-μG) using an in vitro cytokine release assay. The effect of s-μG provided by the desktop random positioning machine on cell-mediated immunity was examined by analyzing interleukin 2 (IL-2), interferon-γ (IFN-γ), tumor necrosis factor alpha (TNF-α), and interleukin 10 (IL-10), in response to immune cell stimulation in whole blood samples (n = 10). Stimuli used were bacterial recall antigens, pokeweed mitogen (PWM), lipopolysaccharide (LPS), or heat-killed Listeria monocytogenes (HKLM). S-μG caused an overall inhibition of the IL-2 and IFN-γ responses to recall antigen and mitogen stimulation. More specifically, s-μG most strongly influenced the levels of all four cytokines elicited by bacterial recall antigen stimulation. In contrast, HKLM-induced TNF-α secretion was elevated. The average concentrations of TNF-α in response to PWM and LPS and IL-10 release stimulated by PWM, LPS, and HKLM were not significantly altered by s-μG. However, a variable response between individual subjects could be observed. In conclusion, our results demonstrate that the in vitro cytokine release assay can detect gravity-related immune alterations. Furthermore, the use of multiple stimuli and the associated changes in cytokine secretion has the potential to reveal information on the underlying mechanisms affected by s-μG.
Introduction
S
Impairment of cell-mediated immunity (CMI) has been reported during spaceflight (Taylor and Janney 1992; Gmunder and others 1994) and could lead to insufficient protection of the human body against infection. Important cytokines that regulate the CMI are the pro-inflammatory cytokines interleukin 2 (IL-2), interferon-γ (IFN-γ), and tumor necrosis factor alpha (TNF-α), as well as the anti-inflammatory cytokine interleukin 10 (IL-10). IL-2 plays a pivotal role during T cell activation and proliferation (Boyman and Sprent 2012). IFN-γ has immunomodulatory properties by controlling the differentiation of T cells and enhancement of leukocyte recruitment (Schroder and others 2004). TNF-α is involved in the initiation, enhancement, and priming of T cell signaling and activation (Hehlgans and Pfeffer 2005). IL-10 is important for the limitation of immune reactions (Ouyang and others 2011). These cytokines are produced by key players in CMI, such as T lymphocytes and macrophages, in response to activation (Belardelli 1995). Since the levels of cytokines in response to stimulation correspond with the immune capabilities of the subject, they can be used as a measure of CMI (van Dooren and others 2013).
In vitro cell culture experiments performed in space or on ground using microgravity simulating devices have shown that (simulated) microgravity accounts for some of the spaceflight-associated changes of the CMI (Gueguinou and others 2009). During the Spacelab missions, in vitro mitogen activation of human lymphocytes was strongly reduced (Cogoli and Tschopp 1985). In addition, when lymphocytes were activated by mitogens during simulated microgravity (s-μG) using the rotating wall vessel or random positioning machine (RPM), the cells showed a suppressed proliferation (Cooper and Pellis 1998; Schwarzenberg and others 1999). In addition, a decreased cytokine response of human T lymphocytes to mitogenic stimulation under (simulated) microgravity has also been reported (Cogoli and others 1993; Cooper and Pellis 1998; Crucian and others 2000). Although these in vitro studies increased our fundamental knowledge, the use of isolated immune cell populations makes it difficult to extrapolate the data to a more clinically relevant situation.
The skin Multitest (Institut Merieux, Lyon, France) was previously used on the US space shuttle and the orbital station MIR (Taylor and Janney 1992; Gmunder and others 1994) to analyze the cutaneous CMI in vivo. These studies demonstrated impaired CMI of astronauts during spaceflight by a reduced skin reaction to common recall antigens. However, due to the risk of antigen sensitization, this assay has been phased out since 2002. An alternative test was developed to evaluate the antigen–dependent cellular immune response in whole blood cell culture, that is, an in vitro cytokine release assay (Feuerecker and others 2013). Immune cell activation and the resulting cytokine secretion can be induced by recall antigens and mitogens associated with different signaling pathways. Recall antigens trigger memory T cells (Farber 2009). The lectin pokeweed mitogen (PWM) induces mitosis in B and T lymphocytes (Ashraf and Khan 2003). Antigens from bacterial origin such as lipopolysaccharide (LPS) and heat-killed Listeria monocytogenes (HKLM) are ligands of toll-like receptor (TLR) 4 and TLR2 (O'Neill and Brint 2005), respectively. The use of whole blood in this assay combines the benefits of an in vitro test with a more truthful representation of the in vivo immune situation compared to isolated immune cells. Furthermore, the assay has been shown to be suitable for monitoring stress-mediated immune modulation and the degree of immunosuppression in clinical conditions (Kaufmann and others 2013). Therefore, it can also be a valuable tool for investigations of space-related immune dysregulation.
To this background, the aim of the present study is to verify the suitability of this in vitro cytokine release assay to detect gravity-related immunological changes. Consequently, the cytokine secretion of immune cells in whole blood in response to 24 h stimulation with recall antigens or mitogens was assessed in both normal gravity and under s-μG using the desktop RPM.
Materials and Methods
Blood collection
Peripheral blood samples were obtained from healthy male subjects (n = 14; age 28 ± 5 years). Volunteers were prescreened with inclusion criteria of no acute medical conditions, no chronic illness, no medication intake, normal blood formula, and nonsmoker. The blood collection procedure was approved by the local SCK•CEN ethics committee and was carried out in accordance with the ethical standards of the Helsinki Declaration of 1975, as revised in 2000. Before blood draw, participants were informed about the aim of the study, and written consent was signed.
Blood samples were drawn in the morning in nonfasted state (between 10:00 am and 11:00 am) into 9 mL heparin-treated tubes (Venosafe, Terumo Europe, Leuven, Belgium). Filled tubes were stored on an orbital rocking platform at room temperature (RT), and assays were performed 2 h after blood sampling.
Whole blood immune cell stimulation
The in vitro cytokine release assay was based on the protocol previously described by Feuerecker and others (2013) with small modifications. Briefly, whole blood aliquots of 750 μL from 10 subjects were transferred under aseptic conditions to the assay tubes (round-bottomed 1.0 mL cryotubes; Nunc, Thermo Scientific, Roskilde, Denmark) prefilled with an equal volume of Roswell Park Memorial Institute 1640 medium (RPMI; Sigma-Aldrich Chemie GmbH, Steinheim, Germany). Used stimuli included a bacterial recall antigen mixture containing diphtheria-, tetanus-, and pertussis-toxoid (combined in 1% Boostrix®; GlaxoSmithKline, Rixensart, Belgium), PWM (0.5 μg/mL; Sigma-Aldrich), LPS (5 μg/mL; Sigma-Aldrich), and HKLM (5 μg/mL; InvivoGen Europe, Toulouse, France). Concentrations used were optimized to the protocol specifications. Fully filled closed assay tubes were subsequently incubated for 24 h at 37°C under normal gravity or s-μG conditions. At the end of incubation, the assay tubes were centrifuged for 15 min at 1,500 g. Supernatant was transferred into Eppendorf tubes and immediately stored at −80°C until further analysis.
Microgravity simulation
A microgravity environment was simulated by placing the assay tubes on the desktop RPM (Dutch Space, Leiden, The Netherlands). Motion mode was set to real random, with random direction and random interval. The respective minimal and maximal frame rotation speed used was 55°/s and 72°/s, resulting in a mean rotation speed of 63.5°/s. During microgravity simulation, the RPM was placed inside an incubator at 37°C. Control samples (1 × G condition) were placed on the base of the RPM to account for vibrational effects.
T cell isolation from peripheral blood samples
T cells were isolated from the diluted whole blood samples (n = 10) before incubation and after 24 h incubation in normal gravity or s-μG on the RPM. Isolation was performed by negative selection using the immunomagnetic EasySep Direct method according to the manufacturer's instructions (STEMCELL Technologies, Grenoble, France). The samples were transferred from the assay tubes to 5 mL round-bottom Falcon tubes (Corning Life Sciences, Amsterdam, The Netherlands). EasySep Direct isolation cocktail and RapidSpheres were added to the whole blood samples at a concentration of 50 μL/mL whole blood. After 5 min incubation and addition of 1 mL RPMI 1,640 medium (Sigma-Aldrich), the tubes were placed in the EasyEights EasySep Magnet (STEMCELL Technologies) for the first round of cell separation. Next, the supernatant along with ±10% pellet was transferred to a new tube. Extra RapidSpheres (50 μL/mL whole blood) were added to the supernatants and incubated, after which the tubes were placed in the magnet for the second round of cell separation. Subsequently, only the clear part of the cell suspension was retained, followed by two more rounds of cell separation on the magnet without addition of RapidSpheres.
Viability assessment of T cells
Viability of the isolated T cells (n = 10) was assessed using an Annexin V-FITC (fluorescein isothiocyanate)/Propidium Iodide (PI) Apoptosis Detection Kit (eBioscience, Bender Medsystems GmbH, Vienna, Austria). The cells were washed in 1 mL phosphate buffered saline and subsequently resuspended in 195 μL 1× binding buffer. Then 5 μL FITC-labelled Annexin V was added, and samples were incubated for 10 min at RT. Unbound Annexin V-FITC was removed by centrifugation and removal of the supernatant. PI was added to the cell pellet following resuspension in 190 μL 1× binding buffer up to a final volume of 200 μL. Data of the viability labeling was collected on an Accuri C6 Flow Cytometer (BD, Becton Dickinson and Company, San Jose, CA) with a blue (488 nm, 20 mW) laser, which was calibrated according to the manufacturer's recommendation. Standard optical filters for FL-1 (530/30 nm) and FL-3 (670 LP) were used for the detection of, respectively, FITC and PI. A quality control with six and eight peak fluorescent beads (BD, Becton Dickinson and Company) and cleaning cycle was performed before experiments to assess both the accuracy (bead count and position) and the cleanliness of the machine. The percentages of viable (Annexin V−/PI−), early apoptotic (Annexin V+/PI−), and dead (Annexin V+/PI+) T cells were analyzed using BD Accuri™ C6 software (version 1.0.264.21).
Measurement of cytokine concentrations
The concentration of four key cytokines controlling the CMI, that is, IL-2, IFN-γ, TNF-α, and IL-10, was simultaneously determined in the supernatant samples (n = 10) using the MILLIPLEX® MAP Human Cytokine/Chemokine Kit (Millipore, Billerica, MA) with respective minimum detectable concentrations of 1.0, 0.8, 0.7, and 1.1 pg/mL, according to the instructions provided by the manufacturer. After washing of the plate, 25 μL of the standards and quality controls together with an equal volume of serum matrix solution were added to the appropriate wells. Assay buffer was used as a background control. The sample wells were filled with 25 μL assay buffer before adding 25 μL of supernatant sample. Subsequently, 25 μL of the antibody-immobilized bead mixture was added to all wells. Then, the plate was sealed, wrapped with foil, and incubated overnight at 4°C on a plate shaker (800 rpm).
The next day, well contents were removed, and the plate was washed twice using a handheld magnet. The plate was placed on the magnet for 60 s to allow complete settling of the magnetic beads. Well contents were removed through gentle decanting of the plate and gentle tapping on absorbent paper to remove residual liquid. The plate was washed by removing the plate from the magnet, adding 200 μL wash buffer, and shaking for 30 min followed by removal of the wash buffer after settling of the beads for 60 s on the magnet. Detection antibodies (25 μL) were added to each well and the plate was sealed, covered with foil and incubated for 1 h at RT on a plate shaker (800 rpm). Next, 25 μL Streptavidin-Phycoerythrin was added to every well and again the plate was sealed, covered, and incubated for 30 min at RT on a plate shaker (800 rpm). After incubation, well contents were removed, and the plate was washed twice as previously described. Beads were resuspended by addition of 150 μL of sheath fluid to the wells and placing the plate on a plate shaker for 5 min. Finally, cytokine levels were measured by the Luminex 200 (Luminex, Austin, TX), and the obtained data were analyzed using xPONENT 3.1 software.
Statistical methods: Data analysis
Viability data are described as mean percentage ± standard deviation. Pre- and postincubation values of the percentage of viable, early apoptotic, or dead T cells were compared, and significance was evaluated with repeated measures one-way analysis of variance (ANOVA) followed by Tukey's multiple comparison test. Cytokine concentrations are presented with single subject data. In the event of levels below the lower assay detection limit, the respective minimum detection limit values were used for statistical analysis.
To achieve normally distributed data, all cytokine concentrations were transformed (log10), and normality was verified by D'Agostino-Pearson omnibus normality test. A repeated measures one-way ANOVA was performed followed by Sidak's multiple comparisons to identify significant differences between basal (no stimulation) cytokine levels and cytokine concentrations after stimulation for both gravity conditions, as well as stimulation in 1 × G versus s-μG. Statistical calculations were performed using GraphPad Prism software version 7.01. Differences between test conditions were considered significant if P values were <0.05.
Results
Viability assessment of T cells in whole blood cultures
Annexin V-FITC/PI staining was performed to determine the influence of the experimental setup on the viability of T cells, that is, incubation of whole blood cultures in closed tubes, with or without microgravity simulation. The percentages of viable, early apoptotic, and dead T cells before incubation (T 0 h) and after 24 h incubation in normal gravity (T 24 h 1 × G) or s-μG (T 24 h s-μG) are listed in Table 1. No significant differences between incubation conditions were observed in the percentage of viable and early apoptotic T cells. Although a significant increase of the fraction of dead T cells occurred after 24 h incubation of whole blood cultures in closed tubes for both gravity conditions, all mean values remained <2%. Preliminary experiments of incubation during 48 h in closed tubes showed a trend toward decreased number of viable cells (data not shown). Therefore, the immune cell stimulation in diluted whole blood cultures, which is discussed further, was limited to 24 h. In addition, incubation of the samples with a stimulus did not affect their viability (data not shown).
Percentage of viable, early apoptotic, and dead T cells before incubation (T 0 h) and after incubation under normal gravity (T 24 h 1 × G) or s-μG (T 24 h s-μG). Values listed as mean (n = 10) ± standard deviation. Significant differences compared to values before incubation are indicated by * P < 0.05, ** P < 0.01 (Repeated measures one-way analysis of variance with Tukey's multiple comparison test).
s-μG, simulated microgravity.
Interleukin 2
IL-2 concentration was determined after 24 h incubation under normal gravity or s-μG in the presence of one of the following stimuli: bacterial recall antigens, PWM, LPS, or HKLM. Basal (without stimulation) IL-2 levels were within the normal physiological range (Fig. 1). Stimulation of the immune cells with bacterial recall antigens led to a significantly increased IL-2 secretion (P < 0.001; Fig. 1a). This rise of IL-2 was strongly inhibited by incubation under s-μG (∼95% decline; P < 0.001). PWM induced a milder but significant IL-2 increment under 1 × G (P < 0.001), which was significantly decreased (∼30%) by incubation under s-μG (P = 0.049; Fig. 1b).
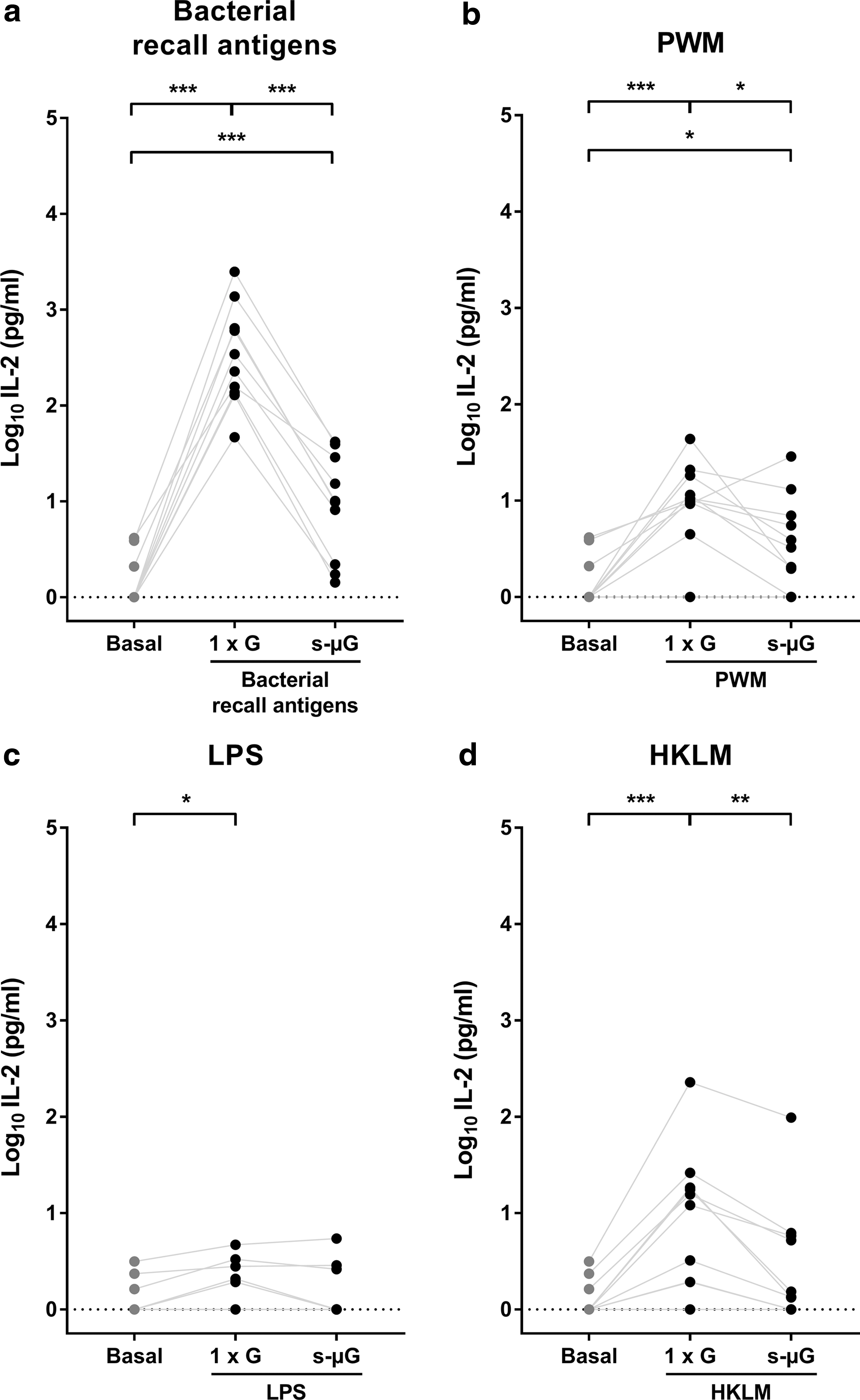
IL-2 concentration in whole blood culture supernatant after 24 h immune cell stimulation under 1
Exposure to LPS increased IL-2 secretion for only 50% of the subjects. However, the mean IL-2 response to LPS was significantly elevated (P = 0.025) and was unaffected by s-μG (P = 0.332; Fig. 1c). The significantly higher IL-2 concentrations after stimulation with HKLM in 1 × G (P < 0.001) were reduced in s-μG by ∼70% (P = 0.012; Fig. 1d). In s-μG, IL-2 levels were only elevated from the normal physiological range for stimulation with bacterial recall antigens (P < 0.001; Fig. 1a) and PWM (P = 0.032; Fig. 1b).
Interferon-γ
Concentrations of IFN-γ were determined after 24 h stimulation in both gravity conditions. Nonstimulated samples contained normal physiological IFN-γ concentrations for all subjects (Fig. 2). These values were significantly increased by all four stimuli, that is, bacterial recall antigens, PWM, LPS, and HKLM, under 1 × G (P < 0.001; Fig. 2 a–d). Incubation under s-μG inhibited this response to different degrees depending on the stimulus used. Stimulation with bacterial recall antigens under s-μG dropped IFN-γ concentration significantly (P < 0.001; Fig. 2a). Furthermore, the IFN-γ response to PWM stimulation was unaffected in s-μG (P = 0.173; Fig. 2b). LPS-induced IFN-γ was significantly reduced by ∼85% in s-μG (P < 0.001; Fig. 2c). Finally, IFN-γ secretion in response to HKLM stimulation under s-μG was significantly decreased by 75% (P = 0.001; Fig. 2d). Despite the differences in IFN-γ secretion between the two gravity conditions, after stimulation in s-μG the values were still significantly higher compared to the normal physiological range for all stimuli (P = 0.023 for bacterial recall antigens; P < 0.001 for PWM, LPS, and HKLM; Fig. 2a–d).

IFN-γ concentration in whole blood culture supernatant after 24 h immune cell stimulation under 1
Tumor necrosis factor alpha
TNF-α measurements in response to stimulation in 1 × G and s-μG were performed after 24 h of incubation. Normal physiological TNF-α concentrations were measured in basal conditions (Fig. 3). Bacterial recall antigens induced a significant elevation of TNF-α (P < 0.001; Fig. 3a). These values decreased in s-μG by ∼50% (P = 0.001). However, the values were still significantly higher compared to the nonstimulated condition (P = 0.002). Concentrations of TNF-α were highly upregulated by PWM, LPS, and HKLM (P < 0.001; Fig. 3b–d). The mean TNF-α induction by PWM and LPS was not significantly affected by incubation in s-μG (Fig. 3b, c). Interestingly, stimulation with HKLM in s-μG led to a significant (∼125%) further increase of TNF-α compared to 1 × G (P = 0.037; Fig. 3d). Consequently, TNF-α remained significantly raised after stimulation in s-μG with PWM, LPS, and HKLM (P < 0.001; Fig. 3b–d).
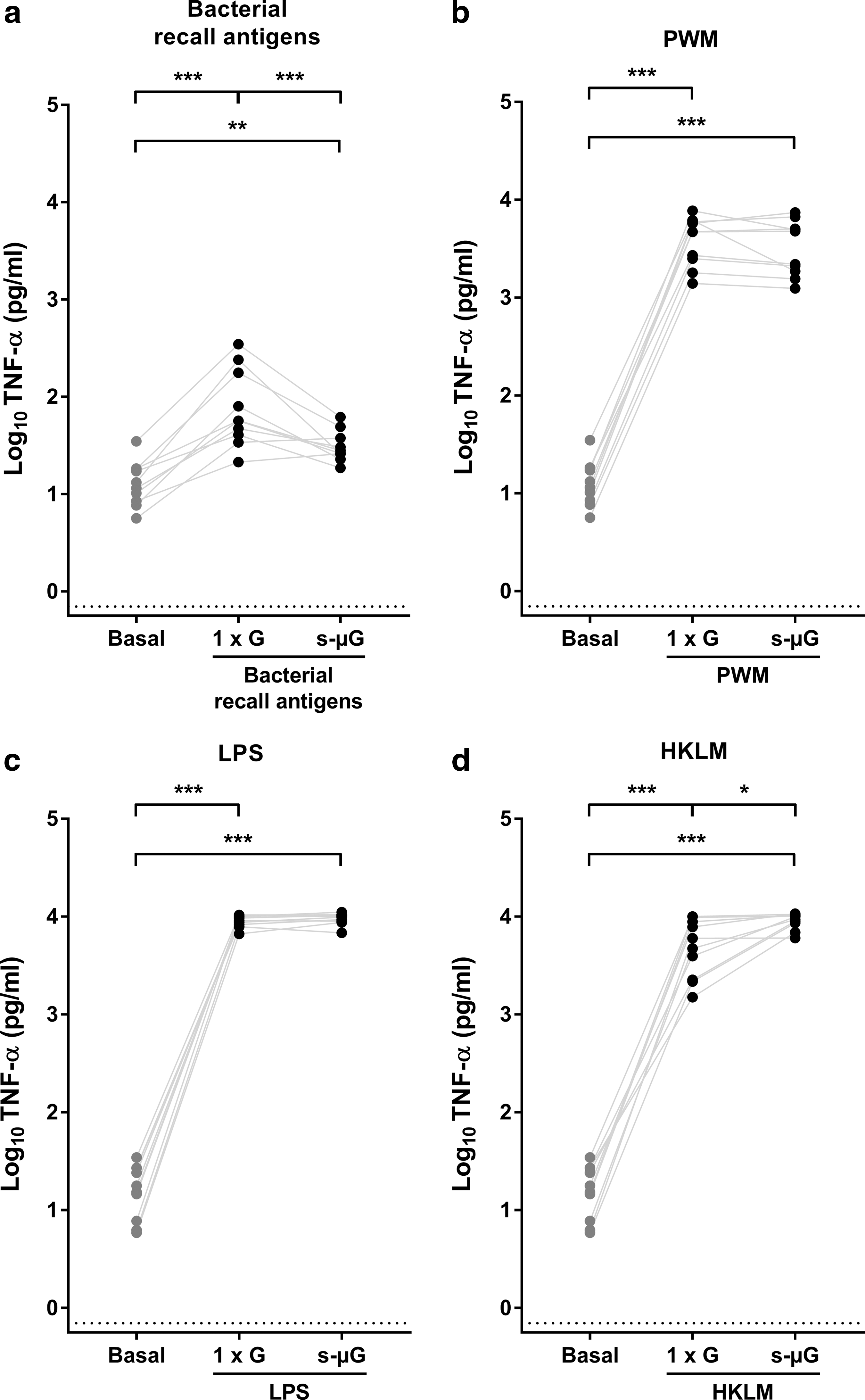
TNF-α concentration in whole blood culture supernatant after 24 h immune cell stimulation under 1
Interleukin 10
In addition to the three pro-inflammatory cytokines, that is, IL-2, IFN-γ, and TNF-α, concentrations of the anti-inflammatory cytokine IL-10 were determined after 24 h of incubation. Without stimulation, IL-10 concentrations were within the normal physiological range (Fig. 4). A slight but significant increase of IL-10 was measured after stimulation with bacterial recall antigens in 1 × G (P = 0.001; Fig. 4a). Incubation under s-μG lowered this response to up to 90% (P = 0.002). PWM, LPS, and HKLM led to a very strong induction of IL-10 in both gravity conditions (P < 0.001), without a discernible difference between the mean IL-10 concentrations after stimulation in 1 × G or s-μG (Fig. 4b–d). When looking at the individual subjects, a variable response between subjects can be observed.

IL-10 concentrations in whole blood culture supernatant after 24 h immune cell stimulation under 1
Discussion
Although it is well known that space travel leads to immune dysfunction, the exact nature of the immunological changes, their specific cause(s), as well as their clinical relevance, has not yet been fully elucidated. Therefore, in the light of future long-duration missions beyond low Earth orbit, it is important to further investigate the impact of space conditions, such as microgravity, on the immune response. The goal of this study was to use an in vitro cytokine release assay in combination with the desktop RPM as a tool to investigate gravity-related immune alterations. The response of the immune cells stimulated with recall antigens and mitogens was measured by analyzing the cytokine response after 24 h. Decreased cytokine secretion of mitogen-stimulated peripheral blood mononuclear cells (PBMCs) isolated from whole blood samples collected during long-duration spaceflight was shown by Crucian and others (2015). However, immune cell isolation and functional testing were performed after return of the samples to Earth. Since the whole blood cytokine release assay does not require cell isolation beforehand, it could present a valuable tool to assess the immune responsiveness of astronauts during space missions. In addition, the use of whole blood instead of isolated cells has the advantage of displaying differences in cytokine secretion due to complex cell–cell interactions and the plasma, thereby better reflecting the in vivo situation.
During the assay, whole blood samples were supplemented with recall antigens or mitogens (PWM, LPS, and HKLM). As expected, all stimuli induced a general cytokine upregulation after 24 h of incubation (Duffy and others 2014). After incubation in s-μG, a shift in the cytokine production in response to stimulation occurred. This shift was dependent on both the cytokines measured and the stimuli used. Incubation under s-μG caused an overall inhibition of IL-2 and IFN-γ responses to recall antigen and mitogen stimulation. A significant decrease of TNF-α and IL-10 was also observed in s-μG conditions for stimulation with bacterial recall antigens. Interestingly, TNF-α levels after stimulation with HKLM were elevated in s-μG compared with 1 × G. The high increase of TNF-α by PWM and LPS and the great upregulation of IL-10 by PWM, LPS, and HKLM under normal gravity was not affected by s-μG.
The secreted cytokine levels in response to in vitro stimulation can be used as a measure of the CMI, since levels of different cytokines correspond with the ability of the subject to induce an effective immune response (van Dooren and others 2013). Cytokines are involved in multiple biological functions, such as the control of cellular homeostasis, the regulation of inflammation, and the response to both pathogens and self-antigens (Yoshimoto and Yoshimoto 2014). Consequently, excessive or insufficient production has been shown to contribute to the pathophysiology of a range of diseases (Fridman and Michon 1990). IL-2 is predominantly produced by activated T cells and is one of the key cytokines involved in T cell activation and proliferation (Boyman and Sprent 2012). Therefore, the inhibited secretion of IL-2 indicates a suppressed T cell activation in s-μG. This is supported by experiments with human PBMCs and primary T cells exposed to real or s-μG showing reduced IL-2 secretion in response to T cell stimuli (Cogoli and others 1993; Pippia and others 1996; Cooper and Pellis 1998; Hashemi and others 1999; Risso and others 2005). IFN-γ has immunomodulatory effects. It plays a role in cellular immunity against viral and intracellular bacterial infections by controlling the differentiation of T cells and enhancing lymphocyte recruitment. Our results show an inhibition of IFN-γ secretion in s-μG for all four stimuli. Previous studies in real and s-μG described both increases and reductions of IFN-γ. In vitro stimulation of PBMCs during spaceflight displayed higher IFN-γ levels compared to ground controls (Chapes and others 1992). Mitogenic stimulation of isolated PBMCs from astronauts' blood samples collected immediately following spaceflight showed a reduced IFN-γ production by CD4+ T cells, whereas CD8+ T cells were unaffected (Crucian and others 2000). Furthermore, the percentage of IFN-γ producing T cells was reduced in the blood of astronauts returning from short-term mission and unchanged after long-term missions (Crucian and others 2008). IFN-γ secretion of PBMCs in s-μG was initially lowered up to 48 h after stimulation, but recovered after 3 days (Cooper and Pellis 1998). TNF-α is a primary factor initiating, enhancing, and priming T cell signaling and activation (Croft 2009). The reduced TNF-α secretion after bacterial recall antigen stimulation in s-μG could therefore lead to an inefficient acquired immune response. However, the increased TNF-α secretion in response to HKLM in s-μG might result in a heightened immune reaction. In contrast with the pro-inflammatory cytokines, IL-10 has important anti-inflammatory properties and is involved in the prevention and limitation of immune reactions (Moore and others 2001). The observed decrease of IL-10 in response to bacterial recall antigens in s-μG could therefore indicate a failure of inflammation resolving mechanisms under s-μG.
In general, the changes in cytokine secretion between both gravity conditions indicate an impaired immune cell activation after stimulation in s-μG. The clinical significance of the observed changes, however, is not straightforward. Since for many of the tested conditions the cytokine response induced by stimulation is also elevated in s-μG compared to the normal physiological range, the question remains if the CMI will be activated enough to be able to counteract possible infections during spaceflight. The variable effects of (simulated) microgravity on the induction of cytokines between cell populations, stimuli, or time points demonstrate the complexity of the CMI. Therefore, there is a need for standardized assays matching the in vivo situation as closely as possible. The ex vivo use of whole blood can serve as an intermediate step for extrapolating observations of in vitro cell cultures to what truly happens in vivo.
In addition to our ground-based experiments describing altered cytokine secretion in s-μG, the in vitro cytokine release assay is currently in preparation to be implemented onboard the International Space Station (ISS) within the framework of the European Space Agency approved MoCISS (Monitoring Cellular Immunity on the ISS) flight experiment. During the MoCISS project, the impact of different gravity conditions and stress hormones on CMI will be monitored by performing the in vitro cytokine release assay before, during, and after long-duration spaceflight. The establishment of in-flight test platforms is necessary to better understand the events occurring during flight.
Cell stimulation in whole blood allows the interaction of the different immune cell types, thus providing insight into the effect of s-μG on the overall immune response. However, it does not give precise information about the impact on singular cell types. The use of different stimuli, including recall antigens and mitogens, and the associated signaling pathways that are activated can offer insights about the different cell types and mechanisms affecting the immune response during microgravity. Stimulation with recall antigens relies on the antigen recognition and activation of memory T cells (Farber 2009). PWM is a lectin that is mitogenic for both T and B cells by binding to glycoproteins on the plasma membrane (Ashraf and Khan 2003). LPS and HKLM are TLR ligands. LPS, found on the outer membrane of Gram-negative bacteria, binds to TLR4, while HKLM, a Gram-positive bacterium, interacts with TLR2 (O'Neill and Brint 2005). Future in-depth pathway analysis is necessary to uncover the specific molecular mechanisms sensitive to microgravity. These findings might lead to new targets for the development of countermeasures. The efficacy of the countermeasures could then be evaluated using the in vitro cytokine release assay by analyzing the cytokine secretion patterns.
In addition, the assay could be useful to study other space factors which are thought to impact the immune competence of astronauts, such as cosmic radiation and psychological stress. Some components of the immune system are among the most radiation sensitive tissues in the body (Moreels and others 2012). Since only a limited number of humans have traveled beyond Earth's magnetic field, there is a lack of data on the effect of cosmic radiation on the human immune system (Fernandez-Gonzalo and others 2017). By performing the cytokine release assay in s-μG combined with radiation exposure, the effect of different stressors of the space environment can be studied in vitro.
Due to the 3D movements of the samples during microgravity simulation, our specific experimental setup required the use of firmly closed tubes. Prolonged incubation of the samples in closed tubes (no gas exchange), as well as the s-μG condition itself, may affect the viability of the immune cells and, thereby, influence the outcome of further investigations of the CMI response. Therefore, the viability of T cells was determined after incubation of the whole blood culture samples in closed tubes in normal gravity or under s-μG. The rather high percentage of early apoptotic T cells before incubation could be caused by the cell isolation procedure. However, the viability of the T cells was not affected by 24 h incubation in closed tubes in both normal gravity and s-μG, nor by stimulation. These results correspond with a previous study using tubes allowing gas exchange that reported no significant changes in the proliferation and cellular viability of PBMCs to mitogen stimulation in 24 h of s-μG, while after 48 h a significant decrease was observed (Martinelli and others 2009). As a consequence, the immune cell stimulation assay was restricted to 24 h incubation.
Furthermore, there was a large variation in the cytokine response between the different subjects reflecting the naturally occurring variance to immune stimulation (Duffy and others 2014). Although the same volume of blood was used in the assays and the blood cell counts of all subjects were within the physiological normal range, the absolute number of immune cells differed among the subjects, which could account for the interindividual differences. For stimulation with the bacterial recall antigens, involving memory T cells, the time since vaccination of the subjects and its effectiveness could also play a role because of the heterogeneity of vaccine immune responses (Poland and others 2007). Nevertheless, the clear suppression of the cytokine response to bacterial recall antigens in s-μG suggests a severe impairment of the acquired immune response to recall antigens. Consequently, it would be interesting to study other recall antigens from bacterial, fungal, or viral origin.
In conclusion, the in vitro cytokine release assay can be used to detect gravity-related immune changes. Therefore, it is a valuable candidate for immune monitoring during spaceflight or in other extreme conditions where the immune system might be affected. In addition, cytokine-based immune monitoring using different stimuli can contribute to revealing key gravity-sensitive steps of the cellular signaling pathways. Better insight into the gravity-related immune dysfunction can support the successful progress of future long-term spaceflight, as well as improve the understanding of immune disorders on Earth.
Footnotes
Acknowledgments
The authors thank the medical service of SCK•CEN for the support during blood sampling. They also thank the blood donors for their contribution to this study. This work was financially supported by the ESA/BELSPO/Prodex IMPULSE contract (CP-90-11-2801-03 and CO-90-14-3539-00, for Belgium) and by DLR 50WB1317 (for Germany).
Author Disclosure Statement
No competing financial interests exist.