
Abstract
Besides initiation of tumor-specific T cell immunity, dendritic cells (DCs) are endowed with tumoricidal activity. Previously, we showed that monocyte-derived DCs of high-grade glioma patients generated in the presence of interferon alpha (IFNα) (IFN-DCs) have impaired cytotoxic activity against tumor necrosis factor alpha (TNFα)-sensitive HEp-2 tumor cells. Herein, we demonstrate that decreased transmembrane TNFα (tmTNFα) expression, but not soluble TNFα (sTNFα) production by high-grade glioma patient IFN-DCs, determines the defective tumoricidal activity against TNFα-sensitive HEp-2 cells. Blocking TNFα-converting enzyme or stimulation of patient IFN-DCs with rIL-2 or dsDNA enhances tmTNFα expression on IFN-DCs and significantly increases their cytotoxicity. Decreased tmTNFα expression on patient IFN-DCs is not caused by downregulation of pNFκB. Neither rIL-2 nor dsDNA upregulates tmTNFα expression on patient IFN-DCs via an increase of pNFκB. The current study shows an important role of tmTNFα as mediator of IFN-DC tumoricidal activity and as molecular target for the restoration of defective DC killer activity in high-grade glioma patients.
Introduction
D
DC tumoricidal activity is mediated through different proapoptogenic ligands, including tumor necrosis factor alpha (TNFα) (Lu and others 2002). TNFα exists in 2 forms, membrane bound or transmembrane (tmTNFα) and soluble TNFα (sTNFα) (Kriegler and others 1988). Both forms can bind to TNF-R1 receptor (p55) and cause apoptosis in target cells (Vandenabeele and others 1995).
Recent data demonstrated that type I interferons (IFNs), in particularly IFNα, in combination with granulocyte-macrophage colony-stimulating factor (GM-CSF), promote the differentiation of monocytes into DCs endowed with natural killer (NK) cell properties (Korthals and others 2007; Papewalis and others 2008). Our previous study also demonstrated that healthy donor monocyte-derived DCs generated in vitro in the presence of GM-CSF and IFNα (IFN-DCs) induce apoptosis and lysis of НЕр-2 tumor cells through the TNFα/TNF-R1 signaling pathway (Tyrinova and others 2013). At the same time, in high-grade glioma patients, cytotoxic activity of IFN-DCs against НЕр-2 cells is significantly decreased (Tyrinova and others 2013), which may be the result of an impaired TNFα-mediated DC cytotoxicity.
Gliomas are the most common type of primary brain tumors (Ostrom and others 2014) that virtually always recur, despite initial treatment with surgical resection, radiotherapy, and chemotherapy (Stupp and others 2009). Nowadays, new promising approaches in treating malignant gliomas based on DC vaccines for the enhancement of the antitumor immune response are being developed. Clinical trials revealed that vaccination with tumor antigen-loaded DCs induces specific immune responses; however, only some of the patients showed prolonged survival (Lasky and others 2013; Finocchiaro and Pellegatta 2016). Therefore, the studies of DC cytotoxicity and mechanisms of its impairment are of basic and applied importance.
Given that DC cytotoxic effect against НЕр-2 cells requires cell-to-cell contact and is not registered when DCs are substituted with their supernatants (Leplina and others 2011), DC cytotoxicity failure found in high-grade glioma patients seems to be related to impairment of tmTNFα expression.
In the present study, we demonstrated that impairments in TNFα expression underlie the defect of monocyte-derived DC cytotoxicity in high-grade glioma patients. In addition, we described the possibility of restoring DC antitumor cytotoxicity in high-grade glioma patients through an upregulation of tmTNFα expression.
Materials and Methods
Patients
The study was conducted in 45 patients with high-grade gliomas, including 24 male and 21 female ranging from 11 to 83 years (median age 51 years). All patients were managed according to established common diagnostic protocol and underwent neurosurgical treatment at the Neurosurgery Department, Novosibirsk Research Institute of Traumatology and Orthopedics, and Federal Neurosurgical Center in the 2013–2016 period. The histological analysis of brain tumor tissues was based on the revised World Health Organization criteria. There were 11 patients with histologically verified glioma Grade III (anaplastic astrocytoma), including 7 patients with newly diagnosed tumor and 4 patients with recurrent glioma. Thirty-four patients had glioma Grade IV (glioblastoma multiforme), including 25 newly diagnosed gliomas and 9 recurrent gliomas. Patient characteristics are shown in Table 1. The control group consisted of 28 age-matched healthy individuals. Informed consent was obtained from all patients and donors according to the Declaration of Helsinki (World Media Association 2000). The protocol for a research project has been approved by a suitably constituted Ethics Committee of Institute of Fundamental and Clinical Immunology, Institute of Traumatology and Orthopedics, Federal Neurosurgical Center, and Institute of Cytology and Genetics, within which the work was undertaken.
Data are present as number (n) of patients (n/total number) with frequencies (percentage) in parentheses.
Patient age at the time of performing experiments.
In vitro differentiation and maturation of DCs
Peripheral blood mononuclear cells (MNCs) were obtained by density gradient centrifugation (Ficoll-Paque; Sigma-Aldrich, Germany) of heparinized whole blood samples. IFN-DCs were generated by culturing of plastic-adherent MNC fraction in RPMI-1640 medium (Sigma-Aldrich), supplemented with 0.3 mg/mL L-glutamine, 5 mM HEPES buffer, 100 μg/mL gentamicin, and 2.5% fetal calf serum (FCS; Sigma-Aldrich), in the presence of rhGM-CSF (40 ng/mL; Sigma-Aldrich) and rhIFN-α (Roferon-A, 1,000 U/mL; Roche, Switzerland) for 4 days at 37°C and 5% CO2 atmosphere. The maturation of DCs was induced by further exposure with a standard lipopolysaccharide concentration (Loison and Gougeon 2014) (10 μg/mL, LPS E. coli 0114: B4; Sigma-Aldrich) for an additional 24 h.
In some experiments, double-stranded human DNA (dsDNA, 5 μg/mL) or rIL-2 (50 U/mL) was added into DC culture simultaneously with LPS. Double-stranded DNA was obtained by the way described in previous reports (Alyamkina and others 2015). Briefly, human DNA preparations were isolated from placentas of healthy women using a phenol-free method. DNA was fragmented in an ultrasonic disintegrator at a frequency of 22 kHz to obtain a mixture of DNA fragments with a size of 200 to 6,000 bp. DNA preparations were dissolved in saline and stored at −20°C. This is a pharmacopeial drug (Registration certificate Medical Drugs of Russia No. 004429/08 of 09.06.2008) and is manufactured under a trademark of Panagen, Ltd.
All experiments were performed with freshly generated DCs. The viability of obtained patient and healthy donor DCs determined by trypan blue exclusion was more than 93%–95% in all cases.
Flow cytometric analysis
Phenotypic analysis of DCs
Phenotype of DCs was determined on an FACSCalibur flow cytometer in large granular lymphocyte gate using CellQuest software (BD Becton Dickinson). Phenotypes of DCs were determined following their direct single- or 2-color staining with fluorochrome-conjugated mAbs specific for CD1a (FITC, Clone HI149), CD11c (FITC, Clone B-Ly6), CD14 (FITC, Clone M5E2), CD86 (FITC, Clone 2331(FUN-1)), HLA-DR (FITC, Clone G46-6), and CD83 (PE, Clone HB15e) (all BD Pharmingen). In each experiment, isotype-matched control mAbs (FITC, Clone MOPC-21 for anti-CD1a, -CD11c, and -CD86 mAbs; FITC, Clone G155-178 for anti-CD14 and -HLA-DR mAbs; PE, Clone MOPC-21 for anti-CD83 mAbs) were included to determine nonspecific background staining. A minimum of 10,000 events was measured from each DC preparation.
tmTNFα expression
tmTNFα expression on IFN-DCs was determined following their direct 2-color staining with FITC- and APC-conjugated mAbs specific for HLA-DR (FITC, Clone G46-6; BD Pharmingen) and TNFα (APC, Clone 6401.1111; BD Pharmingen). As driving monocyte differentiation into either immature or mature DCs is accompanied by high expression of HLA-DR on IFN-DCs (90% frequency), we used HLA-DR molecule as an additional marker of DCs. To distinguish tmTNFα from sTNFα bound to cellular TNF receptor, DCs were treated with glycine buffer (pH = 3) for 90 s at 4°C as described by Gerspach and others (2000). Glycine buffer removed soluble molecules bound to the cells while the transmembrane cytokine remained intact (Fig. 1). Subsequently, cells were washed and incubated with the respective antibodies. In each experiment, isotype-matched control mAbs (FITC, Clone G155-178 for anti-HLA-DR mAbs; APC, Clone MOPC-21 for anti-TNFα; BD Pharmingen) were included to determine nonspecific background staining. A minimum of 10,000 events were measured from each DC preparation.

Effect of glycine buffer treatment on tmTNFα expression by IFN-DCs. IFN-DCs were treated with glycine buffer (pH = 3) for 90 s at 4°C. Subsequently, cells were washed and incubated with FITC- and APC-conjugated mAbs specific for HLA-DR and TNFα. In each experiment, isotype-matched control mAbs were included to determine nonspecific background staining.
Detection of phosphorylated NFκB by flow cytometry
Unstimulated and LPS-stimulated DCs were labeled by HLA-DR-FITC-conjugated mAbs (BD Pharmingen) at 4°C for 30 min. After washes, DCs were intracellularly stained with PE-labeled anti-pNFκB p65 antibody (sc-8008 PE; Santa Cruz Biotechnology) using transcription factor buffer set (BD Pharmingen) according to the manufacturer's instructions. Analysis of pNFκB was performed on a FACSCalibur flow cytometer in large granular lymphocyte gate using CellQuest software following IFN-DC direct 2-color staining with FITC- and PE-conjugated mAbs specific for HLA-DR and pNFκB. In each experiment, isotype-matched control mAbs (FITC, Clone G155-178 for anti-HLA-DR mAbs BD Pharmingen and sc-2866 PE Santa Cruz Biotechnology for anti-pNFκB p65 mAb) were included to determine nonspecific background staining. A minimum of 10,000 events were measured from each DC preparation.
Determination of TNFα production by DCs
DC supernatants obtained from stimulated IFN-DCs were collected and measured for TNFα in duplicate by ELISA using a commercial kit (R & D Systems) according to the manufacturer's recommendations. The level of TNFα production in DC supernatants was calculated to 105 DCs [(pg/mL)/105 DCs].
Cytotoxicity assay
HEp-2 tumor cells (50 × 103/well) were cultured with LPS-activated IFN-DCs in 96-well flat-bottomed plates at a ratio of 1:1 in triplicate. In some experiments, DCs were preincubated for 1 h with rhTNFR1/TNFRSF1A Fc chimera (10 μg/mL; R & D Systems). After a 20-h incubation, 20 μL of 5 mg/mL MTT (3-(4,5-dimethylthiazol-2-yl)2,5-diphenyl tetrazolium bromide; Sigma-Aldrich) solution was added to each well, and the plates were incubated in the dark for 4 h at 37°C. Then, the plates were centrifuged at 1,500 rpm for 5 min, the medium was removed, and 150 μL of dimethyl sulfoxide (DMSO; MP Biomedicals, LLC, France) was added to each well to dissolve the formazan crystals. The absorbance at 492 nm was determined using multiwell spectrophotometer (Thermo Scientific Multiskan FC, Finland). The percentage of cytotoxicity (%) was calculated using the following formula: [1−(absorbance in experimental well with target and effector cells−background absorbance of effector cells/absorbance of untreated target cells)] × 100.
TNFα-converting enzyme inhibition assays
The synthetic metalloproteinase inhibitor TAPI-0 (25 μg/mL; Calbiochem) was added into immature 4-day IFN-DC cultures of glioma patient simultaneously with LPS. After 18–24 h of culture, DCs and DC supernatants were harvested and were determined for tmTNFα expression, TNFα production, and DC cytotoxic activity against HEp-2 cells in MTT assay.
Statistical analysis
Statistical analysis was performed using Statistica 6.0 software for Windows (StatSoft, Inc.). The data are presented as median (Me) and interquartile range (LQ-UQ). The nonparametric Mann–Whitney U test was used to determine statistical significance. Correlations were assessed using the Spearman correlation coefficient. Multiple linear regression was used to determine several factors with possible influence on the cytotoxic activity of DCs. Differences were considered statistically significant for P < 0.05.
Results
High-grade glioma patient IFN-DCs display phenotype with tendency to a delay of maturation
IFN-DCs generated from monocytes of healthy donors are semimature cells (Leplina and others 2015). Despite the high MHC II class- and costimulatory molecule expression, there was a higher number of cells expressing monocyte CD14 molecule and a lower number of cells among IFN-DC population compared with IL4-DC population (Leplina and others 2015). According to our findings, most glioma patient IFN-DCs expressed HLA-DR and costimulatory CD86 molecules reaching the level similar to those of healthy donor DCs (Fig. 2A, B). However, patient IFN-DCs showed higher expression of monocyte marker CD14 (P < 0.05) and immature DC molecule CD1a (P < 0.05). The number of CD83+ cells among patient IFN-DCs was slightly decreased compared with donor IFN-DCs (P < 0.05). Thus, high-grade glioma patient IFN-DCs are characterized by phenotypic profiles with delay of cell differentiation but express equal levels of molecules involved in antigen presentation (HLA-DR, CD86) to donor IFN-DCs.

The phenotypic characteristics of donor and high-grade glioma patient IFN-DCs. LPS-stimulated IFN-DCs were analyzed by flow cytometry for expression of DC-related surface antigens. The histograms represent the expression of the indicated molecules (bold-line histograms) and the corresponding isotype controls (gray-filled histograms) in 5 individual experiments of 18 healthy donors and 5 individual experiments of 18 high-grade glioma patients. The relative percentage of positive cells (%) among the total DC population is given as mean (±SE).
Analysis of membrane and sTNFα expression by high-grade glioma patient IFN-DCs
IFN-DCs derived from high-grade glioma patients were characterized by a 2-fold decreased number of tmTNFα-positive cells (P = 0.00002) compared with IFN-DCs of healthy donors (Fig. 3A, B). A low number of tmTNFα+DCs (hereinafter <5.3%, low quartile of donor range) was observed in 83% of patients and was registered in 18/24 (75%) patients with newly diagnosed gliomas and in 11/11 (100%) patients with recurrent gliomas. Patients with Grade III and Grade IV gliomas experienced similarly low expression of tmTNFα by IFN-DCs (Table 2).

Surface TNFα expression and sTNFα production by high-grade glioma IFN-DCs. Day 4 unstimulated donor and patient IFN-DCs were treated with LPS for additional 24 h.
The data are presented as median and interquartile range (in parentheses) of tmTNFα
DCs, dendritic cells; IFN, interferon; tmTNFα, transmembrane tumor necrosis factor alpha.
Notably, patient IFN-DCs with low tmTNFα expression displayed low cytotoxicity against НЕр-2 cells (Ме 10.6%; Fig. 3С), whereas patient IFN-DCs with intact tmTNFα expression (>5.3%) induced НЕр-2 cell lysis similar to donor IFN-DCs (Ме cytotoxicity 31.2% and 36.3%, respectively). This fact pointed out that impaired IFN-DC cytotoxicity of high-grade glioma patients was associated with low tmTNFα expression.
As shown in Fig. 3D and E, patient IFN-DCs demonstrated a trend to a higher level of sTNFα compared with donor IFN-DCs (P = 0.10). Of interest, concentrations of sTNFα produced by IFN-DCs of newly diagnosed patients (Fig. 3F) were not significantly changed (476 pg/mL versus 634 pg/mL in donors; P = 0.77). However, patients with recurrent glioma (6/7) showed almost 4-fold higher levels of sTNFα compared with donors and newly diagnosed glioma patients (Ме 2333 pg/mL; P = 0.00008 and P = 0.018, accordingly).
Correlation analysis did not reveal any relationships between tmTNFα expression and sTNFα production in either the total group or patients with newly diagnosed and recurrent gliomas. Nevertheless, multiple regression analysis showed a marked positive dependence (coefficient of determination R2 79%, F-criterion 9.4, P = 0.02) of DC cytotoxicity with tmTNFα expression on IFN-DCs (P = 0.009) and the absence of such dependence with sTNFα level (P = 0.59).
The inhibition of TNFα-converting enzyme by TAPI-0 increases tmTNFα expression on IFN-DCs of high-grade glioma patients and enhances DC cytotoxic activity against tumor cells
To determine the role of transmembrane and soluble forms of TNFα in IFN-DC cytotoxicity, DCs were pretreated with the synthetic inhibitor TAPI-0, which blocks TNFα-converting enzyme (ТАСЕ), transforming tmTNFα into sTNFα (Black and others 1997). Adding TAPI-0 simultaneously with LPS into patient DC cultures led to the increase of tmTNFα+IFN-DCs (Fig. 4A) and to the decrease of sTNF production (Fig. 4В). The increase of tmTNFα expression, caused by TAPI-0, was associated with an almost 3-fold enhancement of IFN-DC cytotoxicity (P = 0.068; Fig. 4С). Thus, shifting the tmTNFα/sTNFα balance toward a tmTNFα form due to inhibiting ТАСЕ and, consequently, the decrease of tmTNFα shedding, significantly increases tumoricidal activity of IFN-DCs in high-grade glioma patients.

TACE inhibition enhances cytotoxic activity of high-grade glioma patient IFN-DCs. Day 4 unstimulated IFN-DCs of high-grade glioma patients were treated with LPS and without or with metalloproteinase inhibitor TAPI-0 for additional 24 h.
rIL-2 and exogenous dsDNA upregulate tmTNFα expression on glioma patient IFN-DCs but do not influence sTNFα production
Recent data demonstrated that lymphocyte cytotoxicity can be increased in the presence of some molecules, including IL-2 and exogenous dsDNA (Sarina and others 1997; Raz and others 2000). We also showed that rIL-2 and dsDNA are capable of upregulating the cytotoxic activity of glioma patient IFN-DCs against tumor HЕp-2 cells (Tyrinova and others 2013). However, it remained unclear whether upregulation of tmTNFα contributes to increased DC cytotoxicity. Therefore, as the next step the effect of rIL-2 and dsDNA on tmTNFα expression on patient IFN-DCs was investigated. The enhancement of IFN-DC cytotoxicity on rIL-2 or dsDNA treatment (Fig. 5A) was associated with increased tmTNFα expression (Fig. 5В). The median number of tmTNFα+IFN-DCs in the presence of rIL-2 increased up to 6.0% (1.7 times), and in the presence of dsDNA up to 5.5% (1.6 times), reaching donor median value (6.0%).
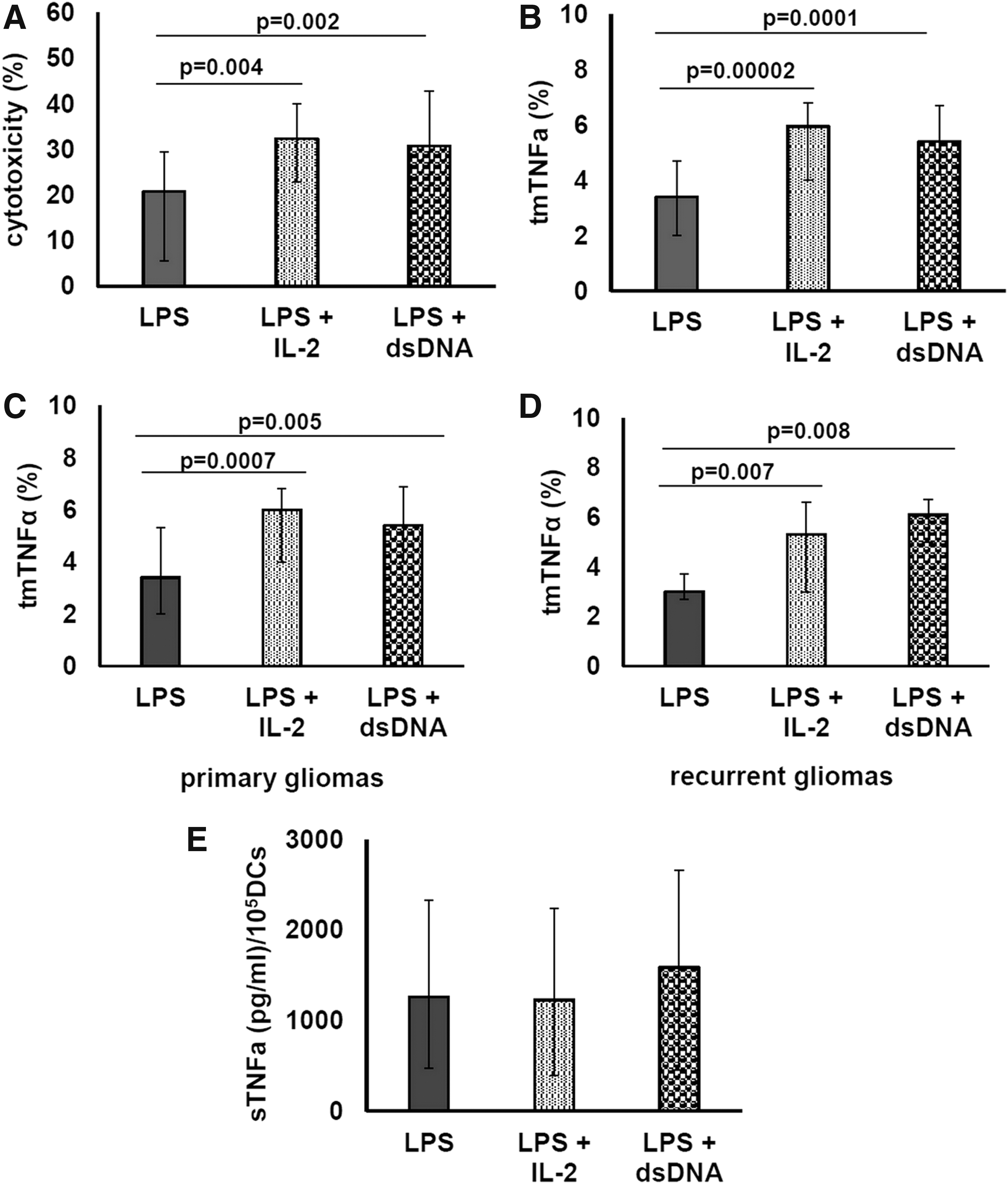
The effect of rIL-2 and dsDNA on tmTNFα expression and sTNFα production by IFN-DCs of high-grade glioma patients. Day 4 unstimulated patient IFN-DCs were treated with LPS and without/with rIL-2 or dsDNA for additional 24 h.
The stimulating effect of rIL-2 on tmTNFα expression by IFN-DCs was observed in the majority of patients (in 83%) and had similar frequency in newly diagnosed tumor patients and recurrent tumor patients (83% and 82%, accordingly). The restoration of the impaired tmTNFα expression by IFN-DCs to normal levels (>5.3%) was observed in half of the patients (57%, 20/35), and was noted both in the newly diagnosed (58%, 14/24) and in the recurrent glioma (55%, 6/11; Fig. 5C, D). The expression of tmTNFα tended to correlate with the cytotoxic activity of rIL-2-treated IFN-DCs (R = 0.475, P = 0.07).
The stimulating effect of dsDNA on the expression of tmTNFα by IFN-DCs was revealed in 77% of total group, including 65% in newly diagnosed tumor and in 100% of recurrent glioma. Normalization of decreased tmTNFα expression by IFN-DCs on stimulation with dsDNA was observed in 60% of glioma patients (15/25). In the newly diagnosed glioma, tmTNFα expression was restored in 9 of 17 (53%) patients and in the recurrent glioma—in 6 of 9 (67%) patients (Fig. 5C, D). Of note, tmTNFα expression by dsDNS-treated DCs strongly correlated with cytotoxic activity of DCs (R = 0.746, P = 0.0014).
Adding rIL-2 or dsDNA to DC cultures did not significantly affect the production of sTNFα by IFN-DCs in the total group, as well as in the newly diagnosed and the recurrent glioma patients (Fig. 5E). Changes in sTNFα production by rIL-2-treated DCs, more than 20%, were registered in 6 of 14 patients and were manifested by an increased TNFα production in 30% (4/14) of patients and by a decrease in sTNFα production in 2 of 14 (14%) cases. Concerning the individual effects of dsDNA, its addition increased sTNFα production in 35%, decreased sTNFα levels in 40% and did not influence secretion of sTNFα in 25% of cases.
Soluble rhTNFR1 reduces dsDNA-mediated enhancement of IFN-DC cytotoxic activity but upregulates rIL-2-stimulated IFN-DC cytotoxicity
To evaluate TNFα involvement in the rIL-2- and dsDNA-induced enhancement of IFN-DC cytotoxic activity of glioma patients against НЕр-2 cells, soluble receptor rhTNFR1, blocking TNFα/TNF-R1-signaling pathway, was utilized (Fig. 6). We did not reveal the effect of rhTNFR1 on LPS-stimulated DCs of glioma patients since this mechanism was believed not to work in coculture of HEp-2 cells and LPS-stimulated IFN-DCs of glioma patients because of low expression of tmTNFα.

The effect of rhTNFR1 on cytotoxic activity of high-grade glioma patient IFN-DCs. Day 4 unstimulated patient IFN-DCs were treated with LPS and without/with rIL-2 or dsDNA for additional 24 h. Harvested DCs were preincubated for 1 h with rhTNFR1 and then cocultured with HEp-2 cells at a ratio of HEp-2:DC 1:1. Evaluation of cytotoxic activity of DCs against HEp-2 cells was performed using the MTT test within 24 h. Data are presented as median (Me) and interquartile range (LQ-UQ) of cytotoxic activity of IFN-DCs versus HEp-2 cells from 5 independent experiments.
Pretreating dsDNA-stimulated DCs with rhTNFR1 molecule reduced cytotoxic activity of IFN-DCs in all the cases by 34% on average (P = 0.04). rhTNFR1 pretreatment of rIL-2-stimulated IFN-DCs did not result in blocking the cytotoxic activity of DCs but in contrast had a stimulating effect. The obtained data indicate that the dsDNA-induced enhancement of DC cytotoxicity in glioma patients is related to an increase of tmTNFα expression, while rIL-2 may activate the TNFα-dependent pathway as well as other signaling pathways of antitumor cytotoxicity.
rIL-2 and exogenous dsDNA enhance tmTNFα expression on IFN-DCs of high-grade glioma patients not via NFκB activation
Transcription factor NFκB is a key participant of multiple signaling pathways, including those connected with TNFα synthesis. To ascertain if the differences in tmTNFα expression on donor and patient IFN-DCs are associated with phosphorylated NFκB expression, and IL-2 and dsDNA upregulate tmTNFα expression and DC cytotoxic activity via NFκB-dependent mechanisms, we analyzed the expression of pNFκB p65 in intact and rIL-2/dsDNA-treated DCs. To find the most optimal time point for detection of phosphorylated NFκB, we studied the kinetics of pNFκB p65 in LPS-stimulated IFN-DCs of healthy donors. As shown in Fig. 7A, the expression of pNFκB was being increased and had reached the maximum values after 30 min on LPS stimulation. A subsequent analysis of pNFκB+cells in patient DCs was conducted after 30 min on stimulation. As shown in Fig. 7B, LPS increased the expression of pNFκB in donor (P = 0.028) and patient (P = 0.02) DCs. Furthermore, donor and patient DCs (both unstimulated and LPS stimulated) did not differ in the number of pNFκB+ cells. The addition of rIL-2 or dsDNA in patient DC cultures did not significantly affect the expression of pNFκB compared with nontreated LPS-stimulated patient DCs (Fig. 7C).
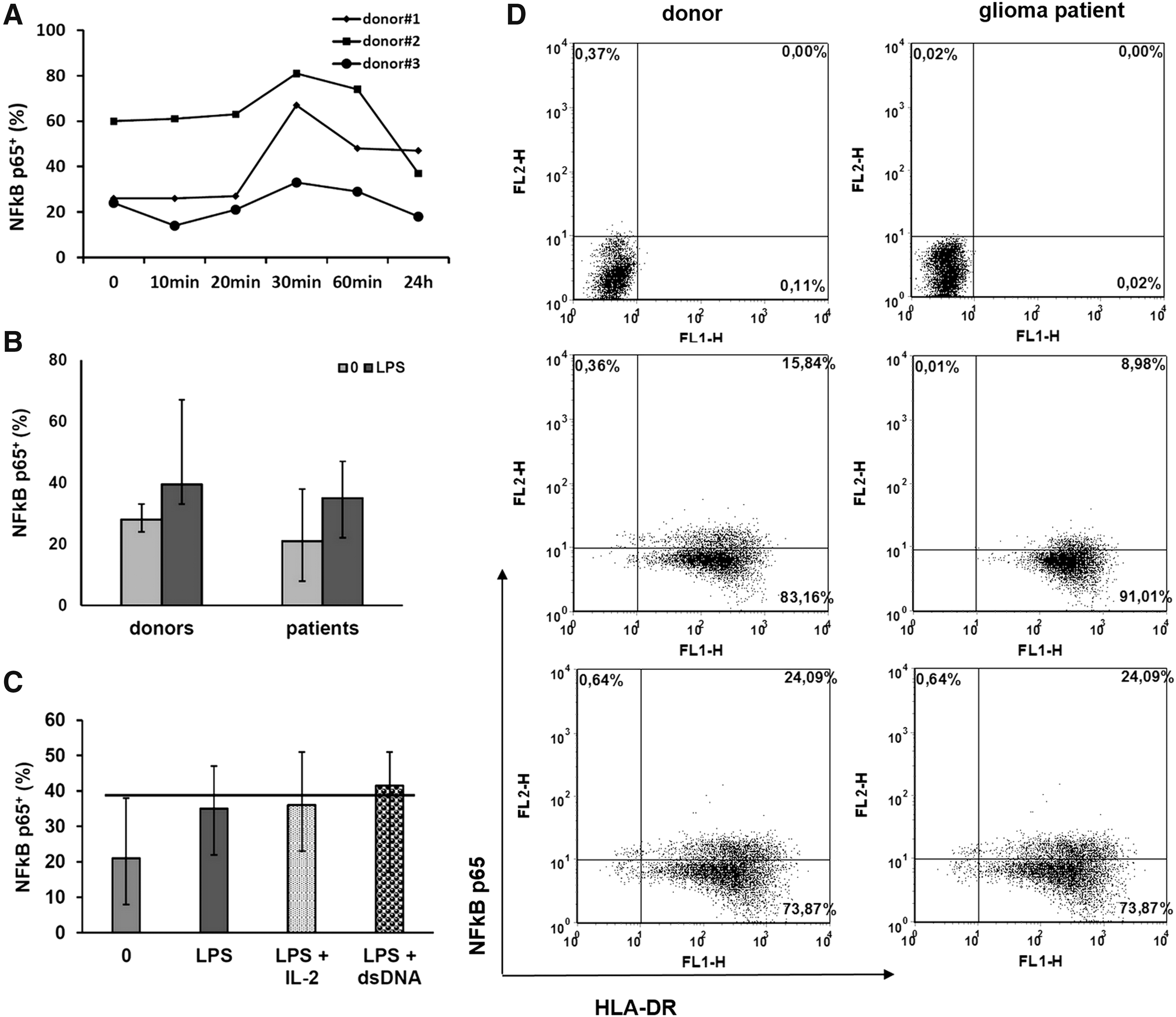
Phosphorylated NFκB level in IFN-DCs of donors and high-grade glioma patients. Day 4 unstimulated donor and patient IFN-DCs were treated with LPS and without/with rIL-2 or dsDNA.
Discussion
IFNα is an early mediator of the innate immunity and is largely produced in response to infection antigens and proinflammatory cytokines. IFNα is a potent upregulator of differentiation and maturation of monocytes (Gessani and others 2014). Monocyte-derived DCs generated in the presence of GM-CSF and IFNα are the unique cell population that can possess properties of myeloid DCs, plasmacytoid DCs, and NK-cells. IFN-DCs are characterized by higher migratory activity and stability compared with IL4-DCs (Paquette and others 1998; Santini and others 2000; Della Bella and others 2004). Moreover, I IFN enhances the cytotoxic potential of effector cells, including DCs (Fanger and others 1999; Liu and others 2001). According to previous studies, IFN-DCs display greater direct killer activity toward tumor cells compared with IL4-DCs (Leplina and others 2015).
Phenotypic maturation is attained when DCs upregulate surface maturation ligands such as CD80, CD83, and CD86 along with the MHC class II molecule (Reis e Sousa 2006). According to published reports, IFN-DCs generated ex vivo from adherent PBMCs display a semimature phenotype with reduced CD14 expression and suboptimal expression of CD83, HLA class II, and costimulatory molecules (Tosi and others 2004; Leplina and others 2015). The current study revealed that LPS-stimulated IFN-DCs of high-grade glioma patients exhibited high expression levels of costimulatory molecule CD86 and class II MHC antigens (HLA-DR), similar to donor IFN-DCs. These data can indirectly evidence that patient IFN-DCs are able to present antigens and stimulate T cells. Nevertheless, we found higher expression of CD14 molecule and immature DC molecule CD1a and lower expression of mature DC marker CD83 on patient IFN-DCs than on donor IFN-DCs. This implies that glioma patient IFN-DCs display signs of delay in maturation and do not reach a developmental stage of donor IFN-DCs.
The defective capacity of DCs to induce potent antitumor immune response in cancer patients is a well-known phenomenon. However, effector functions of DCs in antitumor resistance remain unclear. Recently, we demonstrated that high-grade glioma patients have impaired cytotoxic activity of monocyte-derived IFN-DCs against HEp-2 tumor cells (Tyrinova and others 2013). As lysis of these cells was shown to be mediated via the TNFα/TNFα-R1-signaling pathway and to require a cell-to-cell contact (Leplina and others 2011; Tyrinova and others 2013), we suggested that tumoricidal activity of IFN-DCs is mediated by tmTNFα and the cytotoxicity defect of IFN-DCs is caused by the reduced expression of tmTNFα.
It comes to our attention that donor IFN-DCs exhibited a relatively low tmTNFα expressing population in flow cytometric analysis but possessed high cytotoxic function toward HEp-2 cells. Nevertheless, our data are in line with NK cell cytotoxicity study. Indeed, the MNC population consists of small percentages of NK cells; however, the MNC NK activity displays a high level against NK-sensitive tumor cell lines (Choi and others 2004). Single NK cells can make serial contacts with multiple targets and majority of contacts lead to lysis of target cells (Bhat and Watzl 2007). So, we suggest that, similar to NK cells, a small tmTNFα+ subpopulation of IFN-DCs could effectively kill TNFα-sensitive tumor cells.
The present study demonstrated that IFN-DCs of high-grade glioma patients, including newly diagnosed and recurrent glioma patients, are characterized by a decreased tmTNFα expression. Low tmTNFα+ cell frequency in DC cultures of recurrent glioma patients is associated with enhancement of sTNFα production, whereas the production of sTNFα by IFN-DCs of newly diagnosed glioma patients does not change. Furthermore, the regression analysis shows the cytotoxic activity of IFN-DCs depends on tmTNFα expression, and not on sTNFα production. There are a few reports demonstrating the defective cytotoxic activity of peripheral blood DCs caused by low expression of TRAIL and TNFα molecules in chronic myeloid leukemia patients (Hira and others 2014, 2015). Our findings first show the defect of TNFα-mediated tumoricidal activity of DCs in high-grade glioma patients.
We have previously demonstrated that HEp-2 cells were not sensitive to TRAIL- and FasL/CD95-mediated lysis and blocking of TNFα-dependent signaling pathway led to reduced cytotoxic activity of donor IFN-DCs (Tyrinova and others 2013). Nevertheless, glioma patient IFN-DCs expressing a low level of tmTNFα molecule exhibited slight cytotoxic activity against the HEp-2 cells. This may be due to the fact that DCs are able to mediate the death of tumor cells via classical death-inducing ligands (TRAIL, FasL, TNFα). DC-induced apoptosis in target cells can be dependent on caspase-8 activation, as reported by Vanderheyde and others (2001). DC-induced apoptotic signal is tightly controlled at the mitochondrial level by a balance between antiapoptotic or proapoptotic molecules of the Bcl-2 family (Vanderheyde and others 2001). The cytotoxicity of patient DCs with low expression of tmTNFα, which we observed, could be an effect of intracellular signaling pathways of apoptosis triggered by DC toward HEp-2 cells. Anyway, despite the possible involvement of TNF-independent mechanisms, the defect of tmTNFα/TNF-R-signaling pathway determines significant decrease of glioma patient IFN-DC cytotoxicity toward tumor cells.
Low tmTNFα expression can be caused by an impairment of the TNFα gene expression in DCs of high-grade glioma patients. Besides, a decrease of tmTNFα expression can be a result of its impaired expression at the posttranslation level, in particular, due to high activity of ТАСЕ that cleaves TNFα transmembrane form into soluble form (Black and others 1997). According to our data, decreased tmTNFα expression on IFN-DCs of recurrent glioma patients was associated with enhanced sTNFα production. ТАСЕ is known as a primary sheddase for multiple epidermal growth factor receptor proligands (TNFα, transforming growth factor-alpha, amphiregulin, IL-6, L-selectin), supporting tumor growth and metastasis (Borrell-Pagès and others 2003; Katakowski and others 2009; Scheller and others 2011; Zheng and others 2012). High levels of ТАСЕ expression were found in various solid tumors (nonsmall-cell lung cancer, breast cancer) and were associated with the degree of malignancy (Borrell-Pagès and others 2003; McGowan and others 2007; Kornfeld and others 2011). Enhanced TACE expression has also been shown in high-grade glioma cells and it was indicative of the tumor progression and poor prognosis (Wu and others 2014).
Similar to glioma cells, IFN-DCs of high-grade glioma patients can also have upregulated TACE activity (data from an unpublished study). High ТАCE activity in DCs may contribute to tumor growth as it inhibits tumoricidal activity of DCs by reducing tmTNFα expression. The current study revealed that blocking ТАСЕ using metalloproteinase inhibitor TAPI-0 led to an increase of tmTNFα+DC frequency and reduced sTNFα production by IFN-DCs of high-grade glioma patients, associated with an almost 3-fold increase of IFN-DC cytotoxicity toward HEp-2 cells. These results strongly support the role of tmTNFα in DC-mediated cytotoxicity against НЕр-2 tumor cells.
НЕр-2 tumor cell line is often used as a model to study TNFα-mediated signaling pathways, since it expresses TNFα receptor type 1 TNF-R1 (р55) (Brockhaus and others 1990). However, our previous research has shown that killing of НЕр-2 cells by IFN-DCs required cell-to-cell contact, and supernatants of IFN-DCs, containing sTNFα, failed to lyse НЕр-2 cells (Tyrinova and others 2013). Similarly, Fang and others reported that НЕр-2 cells are resistant to hTNFα-mediated lysis but become susceptible to hTNFα in the presence of cycloheximide (Fang and others 2001), inhibiting the antiapoptotic molecule c-FLIP (Pajak and others 2005). On the other contrary, several previous reports evidence the higher ability of tmTNFα to induce tumor cell apoptosis (Luettig and others 1989; Gruss 1996; Ardestani and others 2013). It can be explained by the fact that tmTNFα forms a more stable complex when binding with receptors compared with sTNFα. As for DCs, Lu and others showed that a decrease of tmTNFα expression and an increase of sTNFα production during maturation of IL-4-induced DCs result in reduction of DC tumoricidal activity (Lu and others 2002). Our results are yet another argument supporting the leading role of tmTNFα in DC-mediated cytotoxicity against НEp-2 tumor cells. Also, decreased expression of tmTNFα is supposed to have pathogenetic importance and to reduce the cytotoxic potential of DCs against TNFα-sensitive tumor cells.
A major point of the present study is an assessment of rIL-2 and dsDNA impact on tmTNFα expression on IFN-DCs. Our previous studies have demonstrated that rIL-2 and dsDNA enhance cytotoxic activity of glioma patient DCs against HЕp-2 cells (Tyrinova and others 2013). However, it remained unclear if these effects were mediated via tmTNFα expression. Herein, we showed that rIL-2 and dsDNA upregulated tmTNFα expression (both in newly diagnosed and recurrent glioma patients) without affecting sTNFα production. Since half the patients displayed normalization of tmTNFα expression on stimulation with rIL-2 and dsDNA, we suggest their possible application to correct the failure of DC cytotoxic activity in high-grade glioma patient DCs. IL-2 is known to upregulate TNFα mRNA, tmTNFα molecule expression, and sTNFα production by NK cells (Caron and others 1999). Myeloid DCs also express a high-affinity IL-2 receptor, CD25, and may be sensitive to modulating effect of IL-2 (Sanarico and others 2006). Herein, we are the first who show the capability of IL-2 to stimulate TNFα-mediated cytotoxicity of DCs.
The mechanisms by which dsDNA activates DCs are not completely understood. However, it was demonstrated that dsDNA can induce a strong adaptive antitumor immune response through TLR-independent signaling pathways (Coban and others 2008), leading to activation of TNFα gene, including protein synthesis and secretion (Ishii and Akira 2006; Coban and others 2008). We observed the strong correlation between tmTNFα expression and DC cytotoxicity in cultures of dsDNA-treated DCs and a lack of significant correlation in cultures of rIL-2-treated DCs. These findings suggest that the stimulation effect of dsDNA is mediated through an increase of tmTNFα expression involving TNFα/TNF-R1 pathways, whereas rIL-2 activates, coupled with enhancement of tmTNFα expression, other signaling pathways. Indeed, blocking tmTNFα with soluble rhTNFR1 showed a decrease of DC cytotoxicity in dsDNA-stimulated DCs, but enhanced cytotoxicity in rIL-2-stimulated DCs. An increase of cytotoxic activity of rIL-2-treated DCs in the presence of rhTNFR1 is an unexpected result that may be explained by the capacity of tmTNFα to work as a reverse signaling receptor (Eissner and others 2004). This mechanism was described for IL-2-activated NK-cells, which expressed tmTNFα and on pretreatment with a soluble TNFR1 increased cytotoxicity due to augmented expression of TRAIL, FasL, perforin, and granzyme В (Yu and others 2009). As НЕр-2 cells are resistant to TRAIL and FasL (Morton and Blaho 2007; Jiang and others 2011), an enhancement of their lysis by rIL-2-treated DCs can be mediated via perforin/granzyme B-dependent mechanism. However, this hypothesis needs to be investigated.
TNFα synthesis is regulated with the transcription factor pNFκB (Zuckerman and Evans 1992), which is upregulated by LPS stimulation (Chiao and others 1994; Muroi and others 1995). Given this, we tried to ascertain if low tmTNFα+DC frequency in cases of high-grade gliomas is a result of pNFκB inhibition. By analyzing the pNFκB, we revealed an equal expression of pNFκB in donor and patient IFN-DCs. Thus, a decreased number of tmTNFα+DCs in glioma patients are not caused by downregulation of pNFκB.
Similar to LPS, IL-2 and dsDNA can activate pNFκB (Arima and others 1992; Ligtenberg and others 2013). Our studies demonstrated that neither rIL-2 nor dsDNA increased pNFκB in patient DCs evidencing that stimulatory effects of rIL-2 or dsDNA on DC cytotoxicity are pNFκB-independent processes and can be mediated through other signaling molecules. For example, IL-2 can stimulate р38 and ERK МАРK-kinases pathways of TNFα synthesis activation (Karnitz and others 1995; Crawley and others 1997). We also cannot exclude that rIL-2 and dsDNA can increase tmTNFα expression on DCs through TACE regulation.
Our work has several limitations. One of the limitations of this study is that we perform monocyte purification by adherence of MNCs for generation of IFN-DCs. In this protocol small lymphocyte population presents in obtained IFN-DC culture, though in less than 10% of all cells. It is therefore to be feared that increased cytotoxic activity of patient IFN-DCs might be associated with NK cell contamination expanded under rIL-2 or dsDNA treatment. However, we can appeal to these concerns because HEp-2 cells used as targets for DCs in the present study are NK-resistant tumor cells (Jewett and others 2003). So, we can exclude the cytotoxic effect of NK cells or other cell types, which could be presented and activated under cytokine treatment as an admixture in DC cultures. Second, we used LPS to trigger DC maturation through TLR4 expressed on DCs. Mature DCs potently stimulate adaptive immunity better than immature DCs, so many vaccine adjuvants currently under development are designed to efficiently induce functional maturation and activation of DCs. There are a few synthetic TLR4 agonists that serve as an effective adjuvant and enhance the capacity of DCs in vivo to immunize against protein antigens. These are monophosphoryl lipid A, glucopyranosyl lipid A, and so on, that is, chemical modifications to lipid A portion of LPS. In our experiments, we stimulated DC maturation with LPS as it was an in vitro model. As known, the glioma microenvironment is strongly immunosuppressive. Glioma cells and glioma-associated cells produce proinflammatory cytokines, but Th2/3 (immunosuppressive) profile is dominant and thus incapable of activating DCs and other effector immune cells. Therefore, IFN-DCs stimulated with TLR4 agonists could be of great interest in terms of clinical use as vaccines for treating glioma.
Further researches are needed to determine the exact mechanisms underlying the impairment of DC tumoricidal activity in glioma patients and to determine molecular targets for the regulation of DC killing activity. Of special interest is the sensitivity of glioma cells to the tmTNFα-mediated lysis and the significance of defective DC cytotoxicity in high-grade glioma progression.
Footnotes
Acknowledgment
This work was supported by the Russian Foundation for basic research “Initiative scientific projects” [grant number 14-04-00446].
Author Disclosure Statement
No competing financial interests exist.