
Abstract
Interleukin (IL)-4 is generally thought to promote tumor cell growth and inhibit apoptosis. However, its role in characteristics of monocytic leukemia cells was rarely reported. In this study, we assessed the role of IL-4 in tumor necrosis factor-related apoptosis-inducing ligand (TRAIL) sensitivity of human monocytes. After incubation with IL-4 for 24 h, death receptor 4 (DR4) was significantly increased without downregulation of TRAIL decoy receptors and antiapoptotic proteins in THP-1 monocytes, and human primary monocytes and U-937 cells also exhibited increased TRAIL-induced apoptosis compared with control. Enhancement of TRAIL-mediated apoptosis by IL-4 was blocked by anti-DR4-neutralizing antibodies. Both upregulation of DR4 and enhancement of TRAIL-mediated apoptosis by IL-4 could be blocked by inhibitors of Janus kinase (JAK)/signal transducer and activator of transcription (STAT), phosphoinositol 3-kinase (PI3K)/Akt, and extracellular signal-regulated kinase to varying degrees. Thus, our data demonstrated a novel effect on TRAIL sensitivity on monocytes and monocytic leukemia cells of IL-4 and suggested that it may be necessary to reconsider the impact of current therapies against IL-4, JAK/STAT, and PI3K/Akt pathways with regard to TRAIL sensitivity.
Introduction
T
In contrast to TNF-α, TRAIL specifically kills transformed cells (Ashkenazi and others 1999), suggesting its potential applications in cancer treatment. However, many tumor cells can resist TRAIL-mediated apoptosis, including myeloid leukemia cells (Jones and others 2003; Tazzari and others 2008; Trivedi and Mishra 2015).
Interferon gamma (IFN-γ) and interleukin-4 (IL-4) are multifunctional cytokines that are produced by Th1 and Th2 cells, respectively, and play important roles in the differentiation of monocytes and macrophages. In addition, IFN-γ and IL-4 are also involved in the pathogenesis of tumors. In general, IFN-γ is thought to be an antitumor cytokine because it was shown to be involved in antiproliferative, antiangiogenic, and proapoptotic effects against tumor cells (Zaidi and Merlino 2011), while IL-4 is considered an oncogenic cytokine owing to its effects on promoting proliferation and preventing apoptosis in various cell types (Ul-Haq and others 2016).
However, in some specific conditions and cell types, IL-4 can also induce or enhance apoptosis through various mechanisms (Li and others 2011), including mitochondrial damage (Bailey and others 2004), CC chemokine receptor type 3 upregulation (Jinquan and others 2003), and proapoptotic protein upregulation (Sarosiek and others 2009), while IFN-γ shows protumor functions through upregulation of regulatory T cells, suppression of cytotoxic T lymphocytes by inducing the expression of indoleamine 2,3-dioxygenase in melanoma cells (Katz and others 2008; Prendergast 2008; Brody and others 2009), and attenuation of neutrophil and myeloid cell infiltration into tissue microenvironments (Manoury-Schwartz and others 1997; Irmler and others 2007). In addition to the mechanisms described above, IFN-γ and IL-4 were also found to regulate tumor cell growth by influencing the sensitivity of tumor cells to TRAIL (Todaro and others 2008).
However, whether TRAIL sensitivity of monocytes, which is the important target of IFN-γ and IL-4, is affected by both cytokines has not been reported. In the present study, we found that IL-4 enhances TRAIL-mediated apoptosis not only in THP-1 and U937 human acute myeloid leukemia (AML) cells but also in primary human monocytes. Our findings provide insights into potential applications of IL-4 in the treatment of monocytic leukemia.
Materials and Methods
Cell culture
THP-1 human acute monocytic leukemia cells were purchased from American Type Culture Collection (Manassas, VA) and cultured in RPMI1640 medium supplemented with 10% fetal bovine serum (FBS) (Gibco) and 50 μM 2-mercaptoethanol (Gibco) at 37°C in an atmosphere containing 5% CO2. U-937 cells (purchased from the Type Culture Collection of the Chinese Academy of Sciences) and primary human monocytes are maintained in RPMI 1640 medium supplemented with 10% FBS (Gibco).
The concentrations of IL-4 and IFN-γ for induction of monocyte differentiation and gene expression studies were usually 20–80 ng/mL and 25–100 ng/mL for 24–48 h, respectively, and our pre-experiment found that 20 ng/mL IL-4 and 25 ng/mL IFN-γ for 24 h have been able to differentiate THP-1 with significant differences in gene expression profile (data not shown). Therefore, the concentrations of IL-4 and IFN-γ used in this study were 20 ng/mL and 25 ng/mL, respectively.
Isolation of human primary monocytes
Human peripheral blood mononuclear cells (PBMCs) were freshly isolated from healthy donors by Ficoll (Sigma) density gradient centrifugation. The collection and use of blood complied with relevant guidelines and institutional practices of the Ethics Committees of the First Affiliated Hospital of Chongqing Medical University. Then, CD14+ monocytes were isolated from PBMCs by the Miltenyi monocyte isolation kit according to the manufacturer's instructions and cultured in RPMI 1640 medium supplemented with 10% FBS.
MTT assay
Cells were seeded at 1 × 105 cells per well in 24-well plates. After incubation with IFN-γ and IL-4, cells were incubated for 4 h with addition of 50 μL of MTT (Sigma) reagent stock solution (5 mg/mL) in the CO2 incubator. Then, 0.5 mL of lysis buffer (0.04N HCL in isopropanol) was added directly per well. Plates were incubated at 37°C and gently shaken for 30 min. The absorbance was then measured at 570 nm.
Reagents and antibodies
Recombinant human IL-4 was obtained from PeproTech (Rocky Hill, NJ) and recombinant human IFN-γ was purchased from Gibco. Recombinant human soluble TRAIL was purchased from Enzo. Wortmannin, ruxolitinib, and FR180204 were purchased from Sigma Aldrich (St. Louis, MO), Apexbio, and EMD Millipore, respectively. The human TNF receptor superfamily member 11b (TNFRSF11B, OPG) enzyme-linked immunosorbent assay (ELISA) kit was from Thermo Scientific Pierce. Antibodies against cleaved caspase-8, Bid, DR4, DR5, DcR1, Bcl-2, Bcl-XL, and survivin were purchased from Cell Signaling Technology (Danvers, MA) and those against DcR2 and glyceraldehyde 3-phosphate dehydrogenase (GAPDH) were purchased from Santa Cruz Biotechnology (Santa Cruz, CA). Neutralizing antibodies against DR4, DR5, and goat IgG isotype control were from R&D Systems.
Terminal deoxynucleotidyl transferase dUTP nick end labeling and caspase-3/7 activity assays
Cells were fixed with 4% paraformaldehyde for 10 min at room temperature. After washing with phosphate-buffered saline (PBS), cells were stained using the DeadEnd fluorometric terminal deoxynucleotidyl transferase dUTP nick end labeling (TUNEL) system (Promega, Madison, WI) according to the manufacturer's instructions. Quantification of TUNEL staining was performed by examining 5 randomly selected fields by an investigator who was blinded to sample identity. Caspase-3/7 activity was determined using a Caspase-Glo-3/7 Assay System (Promega) according to the manufacturer's instructions.
Western blotting
Whole-cell lysates were prepared from cells that had been washed with PBS and harvested by centrifugation. Cell pellets were resuspended in RIPA lysis buffer containing 50 mM Tris, pH 7.5, 150 mM sodium chloride, 1% NP-40, 0.2% sodium dodecyl sulfate, 0.5% sodium deoxycholate, 0.1 mM ethylenediaminetetraacetic acid, and 1% protease and phosphatase inhibitors (Sigma-Aldrich) on ice for 30 min with occasional vortexing. The lysates were then centrifuged at 14,000 g for 15 min; supernatants were collected, and protein concentrations were measured using a Bio-Rad protein assay kit (Bio-Rad, Hercules, CA).
Cell lysates (40 μg) were separated by 10% sodium dodecyl sulfate–polyacrylamide gel electrophoresis and transferred to polyvinylidene difluoride membranes (Immobilon-P membranes; Millipore, Billerica, MA). Membranes were blocked with blocking buffer (5% nonfat milk, 0.1% Tween-20 in tris-buffered saline) for 1 h at room temperature. After incubation with appropriate primary antibodies overnight at 4°C, membranes were then incubated with horseradish peroxidase-conjugated secondary antibodies, and protein bands were detected using the ChemiDoc XRS+ imaging system (Bio-Rad).
Reverse transcription–real-time quantitative polymerase chain reaction
RNA was extracted using TRIzol reagent (Invitrogen, Carlsbad, CA). cDNA was generated using a High-Capacity cDNA Reverse Transcription Kit (Applied Biosystems, Foster City, CA). Reverse transcription–real-time quantitative polymerase chain reaction (RT-qPCR) was performed using a CFX96 Real-Time System (Bio-Rad) with specific sense and antisense primers in a 20-μL reaction volume containing 10 μL of SYBR Green PCR Master Mix (Bio-Rad), 10 μL of 1 μM primer stock, and 40 ng of cDNA. PCR was performed in the following conditions: 95°C for 10 min, followed by 40 cycles at 95°C for 10 s, 58°C for 20 s, and 72°C for 30 s. The relative amounts of mRNA were calculated by the 2−ΔΔCt method. The following primers were used: GAPDH forward: 5′-ACAACTTTGGTATCGTGGAAGG-3′ and reverse: 5′-GCAGATCGGACCGGATACTG-3′. DR4 forward 5′-GCGGGGAGGAT TGAACC AC-3′ and reverse: 5′-CGACGACAAACTTGAAGGTCTT-3′.
Statistics
Each experiment was repeated at least 3 times with 2 technical replicates each, unless indicated otherwise, as this was generally sufficient to achieve statistical significance for differences. Statistical significance between groups was calculated by using 1-way analysis of variance, followed by Tukey's test, and statistical significance between the 2 groups was calculated by 2-tailed unpaired Student's t-test using commercially available statistical software (SigmaPlot 11.0 for Windows; Systat Software, Inc., San Jose, CA). Data are presented as means ± standard deviations, and differences with P values lower than 0.05 were considered statistically significant.
Results
IL-4 enhanced TRAIL-mediated apoptosis in THP-1 cells
We first detected the impact of IFN-γ and IL-4 on cell viability of THP-1 cells using the MTT assay. As shown in Fig. 1A, after 48 h of incubation with IFN-γ and IL-4, no significant differences of optical density value among control, IFN-γ-treated, and IL-4-treated groups were observed (1-way ANOVA, P > 0.05). For TUNEL and caspase-3/7 activity, at 24 h after incubation of THP-1 cells with IL-4 (20 ng/mL) and IFN-γ (25 ng/mL), soluble TRAIL was directly added into the medium at a final concentration of 100 ng/mL, cells were then incubated for further 24 h and collected to perform tests. As shown in Fig. 1B and D, TRAIL induced a TUNEL-positive signal just in 4.89% ± 0.97% of cells in the untreated control group. However, the TUNEL-positive signal reached 21.83% ± 1.49% in the cell group pretreated with IL-4; this was significantly higher than that in the untreated control group and in the IFN-γ-pretreated group (5.4% ± 0.99%; Fig. 1B).
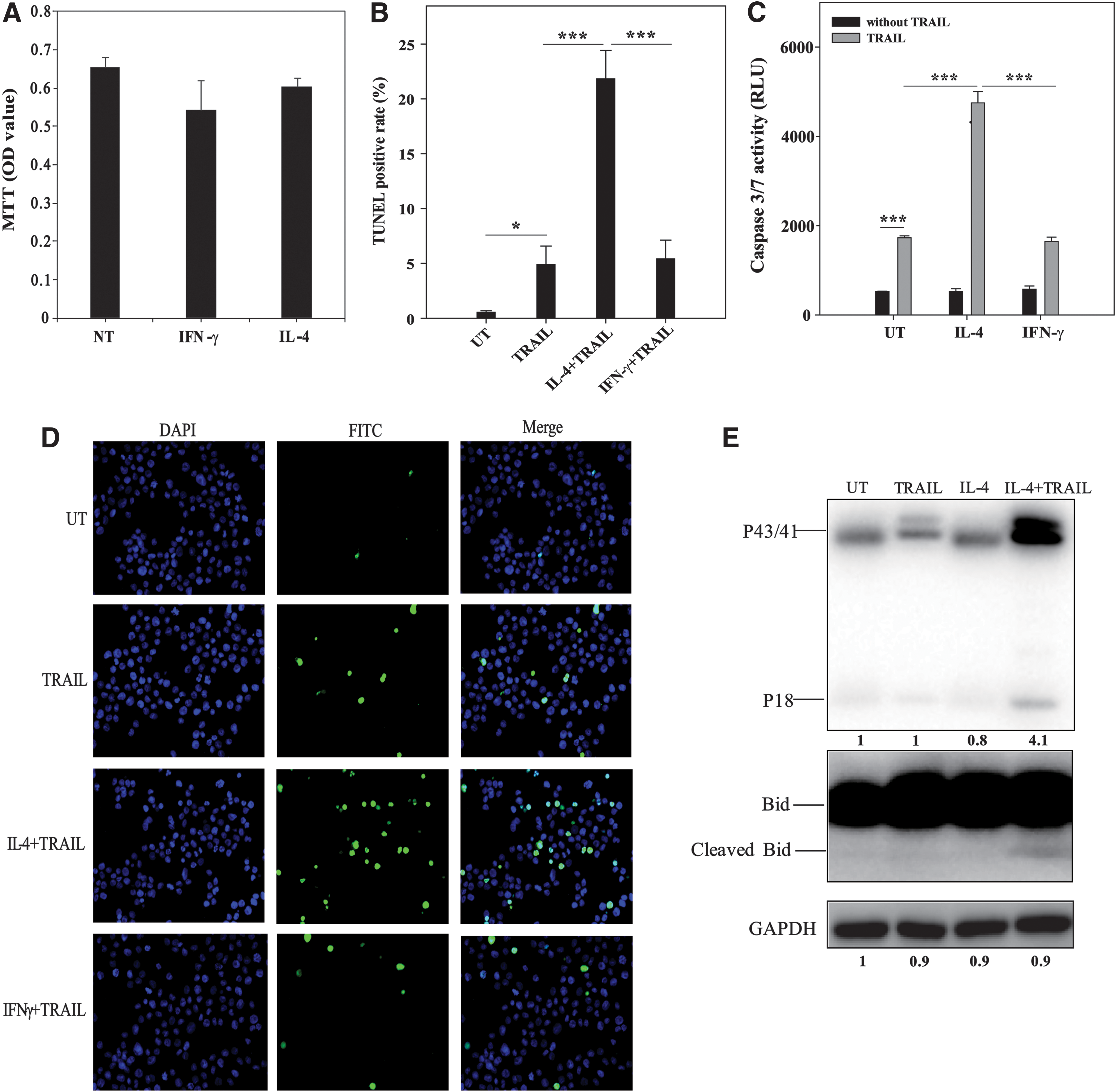
IL-4 enhanced TRAIL sensitivity in THP-1 cells.
In addition to apoptosis, DNA fragmentation presented here by TUNEL-positive signals can also occur in necrosis, pyroptosis, and some other types of cell death, which could be mediated by TRAIL. Therefore, caspase-3/7 activity was also performed to confirm if it is apoptosis. Similar to the TUNEL results, caspase-3/7 activity was significantly higher (4748 ± 254.34) in IL-4-pretreated cells than in untreated cells (1727.67 ± 43.66) and IFN-γ-pretreated cells (1650 ± 92.08) induced by TRAIL (Fig. 1C).
Previous studies have found that TRAIL activates caspase-8 through binding to DRs. Activated caspase-8 can cleave caspase-3 directly to induce apoptosis through the extrinsic apoptotic pathway and cleave Bid to damage mitochondria to induce apoptosis through the intrinsic apoptotic pathway (de Miguel and others 2016). In the present study, apoptosis of THP-1 cells mediated by IL-4 in combination with TRAIL was also involved in the activation of caspase-8 (P18 density compared with the untreated group, t-test, P < 0.05), and the mitochondrial pathway may be involved since cleavage of Bid was also observed in the IL-4 combined TRAIL-treated group (Fig. 1E).
Enhancement of TRAIL sensitivity by IL-4 in THP-1 cells involved in DR4 upregulation
The sensitivity of cells to TRAIL mainly depends on the balance between its receptors and decoy receptors, including DR4, DR5, DcR1, DcR2, and OPG. As shown in Fig. 2A, IL-4 significantly upregulated protein levels of DR4 in a time-dependent manner (t-test, P = 0.0019). Although IL-4 rapidly upregulated DR5 in THP-1 cells and peaked at 8 h post-treatment, it gradually decreased, showing no difference between pretreatment and 24 h post-treatment (t-test, P = 0.1041). However, IL-4 had no effect on both DcR1 (1-way ANOVA, P = 0.2443) and DcR2 (1-way ANOVA, P = 0.2012). For soluble OPG that is secreted into the culture medium, we used ELISA to detect its concentration in the supernatant after IL-4 stimulation. OPG was undetectable until 24 h post-IL-4 treatment (Fig. 2B).

IL-4 increased TRAIL-mediated apoptosis in THP-1 cells by upregulation of DR4.
Besides the expression level of DR and decoy receptors, the antiapoptotic proteins involved in mitochondrial apoptosis are also involved in regulation of TRAIL sensitivity. Therefore, Bcl-2, Bcl-xl, and survivin, which might be regulated by IL-4, were also detected. Consistent with previous reports (Conticello and others 2004; Di Stefano and others 2010), IL-4 could upregulate the expression of Bcl-xl (P = 0.011) and survivin (P < 0.01) at 8 h post-treatment, but then gradually decreased. Bcl-2 protein in THP-1 cells was not influenced by IL-4 (1-way ANOVA, P = 0.478). Since no decreases in decoy receptors or antiapoptotic proteins were observed at 24 h post-IL-4 treatment, we highly suspected that upregulation of DR4 is the main reason for enhancement of TRAIL sensitivity in THP-1 cells mediated by IL-4. As with its protein levels, DR4 mRNA levels also increased gradually with IL-4 treatment (Fig. 2C), suggesting that IL-4 upregulates DR4 expression at the pretranscriptional level.
To further confirm the role of DR4 in IL-4-mediated enhancement of TRAIL sensitivity of THP-1 monocytes, we used neutralizing antibodies against DR4 and DR5 to perform a rescue experiment. The increases in IL-4 plus TRAIL-mediated DNA fragmentation and caspase-3/7 activity were almost completely abolished by treatment with neutralizing antibodies against DR4 (Fig. 2D, E). Although IL-4 did not upregulate the expression of DR5 at 24 h post-treatment, the neutralizing antibody of DR5 still partially blocked TRAIL-induced apoptosis, which might be related to the baseline expression level of DR5, and DR5 delivers the death signal more effectively than DR4 (Phillips and others 1999; Chen and others 2015).
IL-4-mediated upregulation of DR4 and TRAIL sensitivity also occurred in the U-937 monocytic cell line and human primary monocytes
Since THP-1 is a monocyte-derived tumor cell line, some of the key experiments were repeated in human primary monocytes and other monocytic leukemia cells to evaluate whether upregulation of DR4 by IL-4-mediated enhancement of TRAIL sensitivity correlated with its tumor characteristics or monocyte characteristics. As shown in Fig. 3A, B, IL-4 also upregulated DR4 in both U-937 cells and human primary monocytes, and enhancement of TRAIL sensitivity mediated by IL-4 could be blocked by a neutralizing antibody against DR4 in both U-937 cells and human primary monocytes (Fig. 3C, D).

IL-4 upregulates DR4 to enhance TRAIL sensitivity of U-937 cells and human primary monocytes. IL-4 (20 ng/mL) for 24 h increased DR4 protein expression in U-937 cells
IL-4-mediated DR4 upregulation involved Janus kinase/signal transducer and activator of transcription, extracellular signal-regulated kinase, and phosphoinositol 3-kinase/Akt pathways
As described above, IL-4-enhanced TRAIL sensitivity might be caused by upregulation of DR4 mRNA and protein in a time-dependent manner, suggesting that the upregulation was likely to be a direct effect of IL-4 rather than a secondary effect at the transcriptional level. To date, IL-4 has been shown to regulate several signaling pathways, including phosphoinositol 3-kinase (PI3K)/Akt (Lee and others 2003), extracellular signal-regulated kinase (ERK) (Wery and others 1996), and Janus kinase (JAK)/signal transducer and activator of transcription (STAT) (Paul 2015). Therefore, inhibitors of these pathways were recruited in subsequent experiments to identify which pathways were involved in IL-4-mediated DR4 upregulation and increased TRAIL sensitivity.
First, we evaluated the effects of inhibitors of 3 pathways on IL-4-induced DR4 upregulation. As shown in Fig. 4A, all 3 inhibitors (ruxolitinib [JAK inhibitor], wortmannin [PI3K/Akt inhibitor], and FR180204 [ERK inhibitor]) could inhibit IL-4-induced upregulation of DR4 to varying degrees; among these inhibitors, ruxolitinib showed the most substantial inhibitory effects, reaching ∼90% inhibition. Consistent with the degree of inhibition of DR4 upregulation induced by IL-4, all 3 inhibitors could block IL-4-induced upregulation of TRAIL-mediated DNA fragmentation and caspase-3/7 activity to different degrees (Fig. 4B, C).

Signal pathways involved in IL-4-mediated upregulation of DR4 and TRAIL-induced DNA fragmentation and caspase-3/7 activity. After pretreatment with ruxolitinib (10 μM), wortmannin (1 μM), and FR180204 (20 μM) for 1 h, IL-4 (20 ng/mL) was added to the medium for 24 h.
Discussion
Acute leukemia can be divided into acute lymphoblastic leukemia (ALL) and AML. ALL involves both children and adults, whereas AML mainly affects adults. Many in vitro studies have found that TRAIL alone is insufficient to induce apoptosis in leukemia and lymphoma through cell type-specific mechanisms (Riccioni and others 2005; Secchiero and others 2007; Tazzari and others 2008; Sung and others 2010). In AML, Riccioni and others found that resistance to TRAIL could be explained by upregulation of DcR1 and DcR2 and downregulation of DR4 (Riccioni and others 2005).
The human THP-1 cell line has been developed from peripheral blood leukocytes of a patient with acute monocytic leukemia (Tsuchiya and others 1980) and it was used as an in vitro model of monocytes and monocytic leukemia. Although the impact of IFN-γ and IL-4 on differentiation of THP-1 cells has been researched in a number of studies focusing on monocyte differentiation, whether IFN-γ and IL-4 regulate its tumor-related properties has seldom been reported. In this study, we reported for the first time that IL-4 could enhance TRAIL-mediated apoptosis of THP-1 acute monocytic leukemia cells through upregulation of DR4.
Although we found that IL-4 also upregulated the expression of the antiapoptotic proteins, Bcl-xl and survivin, as previously reported in THP-1 cells (Todaro and others 2008; Di Stefano and others 2010; Roca and others 2012), the expression levels of these 2 proteins begin to decline after 8 h of IL-4 treatment, resulting in lower fold increases in survivin and Bcl-xl expression than DR4 expression after 24 h of IL-4 treatment compared with baseline. This might explain why IL-4 enhanced TRAIL sensitivity in THP-1 cells and even Bcl-xl and survivin were upregulated. Alternatively, THP-1 cells might be the type I cells that can undergo apoptosis without mitochondrial dysfunction (Chung and others 2016; Zhang and others 2016). Thus, upregulation of Bcl-xl and survivin may not block apoptosis in the context of enhanced TRAIL-mediated caspase-8 and caspase-3 activation in response to IL-4-mediated DR4 upregulation.
In addition to THP-1 monocytic leukemia cells, IL-4-mediated enhancement of TRAIL sensitivity by upregulation of DR4 was also found in another monocytic leukemia cell line U-937 and human primary monocytes in the present study, which means that IL-4-mediated upregulation of DR4 might be related to characteristics of monocytes rather than tumors. One of the most important features of TRAIL is the specific induction of tumor cell death without toxicity to normal cells, which may be involved in the regulation of other cell functions such as monocyte migration (Wei and others 2010). The present study also shows that TRAIL alone cannot induce cell death of human primary monocytes, but after IL-4 stimulation, even human primary monocytes are more sensitive to TRAIL than before IL-4 treatment, suggesting that IL-4 in combination with TRAIL might play roles in the balance between M1 and M2 subtype monocytes since M1 subtype monocytes express TRAIL upregulated by IFN-γ (Griffith and others 1999).
IL-4 binding to its receptors induces phosphorylation of JAK1 and JAK3 (Nelms and others 1999; Jiang and others 2000), which then mediate the phosphorylation and translocation of STAT6 into the nucleus and activate PI3K/Akt and ERK1/2 pathways through phosphorylation of insulin receptor substrate (IRS)-1 and IRS-2 (Paul 2015). In this study, we found that the JAK inhibitor ruxolitinib almost completely abolished IL-4-mediated upregulation of DR4 and TRAIL sensitivity, and inhibitors of PI3K/Akt and ERK also blocked this process to varying degrees. These results suggested that JAK/STAT, PI3K/Akt, and ERK1/2 may be involved in IL-4-mediated upregulation of DR4.
Interestingly, JAK/STAT (Furqan and others 2013), PI3K/Akt, and ERK (Steelman and others 2004) pathways are highly activated in AML, and inhibitors of JAK and PI3K/Akt developed for AML treatment, including ruxolitinib, are currently undergoing clinical trials (Moore and others 2013; Annesley and Brown 2015; Fransecky and others 2015). However, the present results suggested that inhibition of JAK/STAT and PI3K/Akt may downregulate IL-4-mediated DR4 expression and block TRAIL sensitivity in THP-1 human leukemia cells. Since TRAIL is an important factor in human tumor surveillance and defense, further studies are needed to determine whether these inhibitors affect DR4 expression in different cell types to avoid the decrease in TRAIL sensitivity related to the use of these inhibitors.
To date, researchers have developed a series of cytotoxic drugs that can upregulate DR4, and some signaling pathways have been shown to be involved in DR4 regulation. The ERK pathway and its downstream transcription factor AP-1 have been extensively studied (Guan and others 2002; Drosopoulos and others 2005; Yao and others 2016). Other pathways include the autophagy pathway (Kim and others 2016), p53 (Du and others 2016), Notch (Dhandapani and others 2011), and nuclear factor-κB (Mendoza and others 2008). In the IL-4-activated signal cascades described above, JAK acts upstream of PI3K/Akt and ERK, which could explain why the JAK inhibitor ruxolitinib almost completely blocked upregulation of DR4 and TRAIL sensitivity enhanced by IL-4. In addition to the ERK pathway, which was reported previously by other researchers, our results showed that the PI3K/Akt pathway was also involved in IL-4-mediated DR4 upregulation and played a more important role in this process than the ERK pathway.
In summary, our present results showed (for the first time) that IL-4 enhanced TRAIL sensitivity in THP-1, U-937, and human primary monocytes by upregulation of DR4. JAK/STAT, ERK, and PI3K/Akt signaling pathways may be involved in this process. Our findings identified a new antitumor role of IL-4 and highlighted the need for more careful and comprehensive assessments of the effects of JAK, ERK, and PI3K/Akt inhibitors on TRAIL sensitivity, particularly in monocytic leukemia.
Footnotes
Acknowledgments
This work was supported by grants from the National Natural Science Foundation of China (No. 81300317) and Chongqing Research Program of Basic Research and Frontier Technology (No. cstc2015jcyjnBX0079).
Author Disclosure Statement
No competing financial interests exist.