
Abstract
Virus-specific T cells are critical in mediating the pathogenesis of hepatitis B virus (HBV) infection. Interferon gamma (IFNγ)-producing T cells are associated with resolution; in contrast, interleukin-17 (IL-17)-producing T cells are linked to exacerbation of liver inflammation and injury. Checkpoint receptors stringently regulate T cell functions, with their expression profiles varying on different T cell subsets. Blockade of checkpoint receptors may be an effective therapeutic strategy for chronic hepatitis B (CHB); however, blockade may also inadvertently exacerbate proinflammatory responses. In this study, we sought to determine the balance of inflammatory and antiviral T cells and determine their inhibitory receptor profile. The frequency of total and HBV antigen-specific Th17 and Tc17 cells was higher in CHB patients compared with healthy controls (HCs). Th17 and Tc17 cells in CHB patients had significantly lower expression of T cell immunoglobulin and mucin domain protein-3 (TIM-3) compared with HCs, with no difference in programmed death-1 (PD-1) or CD244 expression. Conversely, Th1 and Tc1 cells in CHB patients hyperexpressed PD-1 and CD244, while TIM-3 expression was comparable in both cohorts. During CHB, antiviral IFNγ T cells hyperexpress multiple immune inhibitory receptors driving their functional impairment. In contrast, inflammatory Th17/Tc17 cells hypoexpress TIM-3, but not PD-1 or CD244. Checkpoint inhibitors for CHB should target PD-1 or CD244 to allow restoration of IFNγ responses without affecting inflammatory IL-17 production.
Introduction
C
It is well described that viral-specific T cells producing interferon gamma (IFNγ) are essential in limiting HBV infection (Chisari and Ferrari 1995) and the key role of IFNγ in mediating host control has been described in multiple studies (Guidotti and others 1999; Phillips and others 2010). This potent cytokine has been shown to be associated with resolution of acute hepatitis, whereas weak induction of this Th1 response results in the progression to persistence (de Waal Malefyt and others 1991; Hyodo and others 2003; Hyodo and others 2004; Rehermann and Nascimbeni 2005). We have also shown that induction of a favorable IFNγ-specific T cell response in CHB patients to antiviral treatment is associated not only with a beneficial treatment outcome but may also be a prerequisite to maintain control of replication (Cooksley and others 2008; Chokshi and others 2014).
In contrast, an interleukin-17 (IL-17)-rich environment in the liver during CHB is associated with exacerbation of inflammatory processes (Ye and others 2010), development of fibrosis (Du and others 2013), progression to cirrhosis (Shi and others 2015), and progression to hepatocellular carcinoma (Huang and others 2014). CD4+ and CD8+ IL-17-producing cells, termed Th17 and Tc17, respectively, are the main producers of IL-17 and are also known to orchestrate strong inflammatory responses downstream by producing an additional cocktail of proinflammatory cytokines, including IL-6, IL-22, and TNF-α (Harrington and others 2005; Wang and others 2013).
The mechanisms by which IL-17-producing cells are regulated and the factors that influence their function in vivo are not well understood and are of importance to develop immunomodulatory therapies for CHB. Moreover, there is a paucity of understanding regarding the role of HBV-specific Th17 and Tc17 cells during CHB as most studies focus on nonspecific IL-17-producing cells.
T cell functionality depends upon a dynamic balance of positive and negative costimulatory pathways (Riva and Chokshi 2018). The different functions of HBV-specific T cells are tightly regulated by immune homeostatic control measures to achieve a balance between efficient control of viral replication and unnecessary immunopathological injury. Several immunoregulatory signatures, also known as checkpoint receptors, have been demonstrated to play a role in dysregulation of the antiviral T cell responses in CHB infection (Callendret and Walker 2011).
There is much promise in the targeting of these receptors as a mechanism of restoring T cell functionality in CHB (Pham and others 2016) and also in hepatocellular carcinoma (Kudo 2017), where the aim is to restore antiviral and antitumor responses (Ye and others 2015) with particular focus on reinstating IFNγ T cell responses. However, their impact on other T cell subsets is not well described.
Programmed death-1 (PD-1) is one of the best characterized immune inhibitory receptors expressed on T cells during chronic infections and cancers (Sharpe and others 2007; Topalian and others 2012a). We have also shown a direct correlation between viral replication rates and PD-1 expression on virus-specific T cells in CHB patients (Evans and others 2008). Recent clinical data have also shown this to be a promising strategy for reversal of T cell impairment in cancer (Brahmer and others 2012; Topalian and others 2012b; El-Khoueiry and others 2017).
T cell immunoglobulin and mucin domain protein-3 (TIM-3), a member of TIMs, is another immunoinhibitory molecule that negatively regulates T helper type 1 immunity, and we have shown it to be involved in impairment of an appropriate immune response against pathogens (Markwick and others 2015). There is also a growing body of evidence to illustrate that TIM-3, with its ligand Galectin-9, has an influential role in antiviral immune evasion in a number of chronic viral infections, most recently HBV, where the upregulation of both TIM-3 and galectin-9 contributes to inhibition and depletion of T cells as they infiltrate the HBV-infected liver. Further evidence shows that hypoexpression can lead to overactivation of lymphocytes with bystander immunopathological damage and autoimmunity (Zhu and others 2005).
CD244 is a high-affinity ligand for CD48. The interactions between CD244:CD48 are complex, providing both stimulatory and inhibitory effects to regulate target cell lysis by cytotoxic T cells and natural killer cells. These processes are critical for viral clearance and generation of effector/memory T cells (Lee and others 2003; Clarkson and Brown 2009). CD244 acts as an inhibitory receptor on HBV-specific CD8+ T cells (Raziorrouh and others 2010). In chronically infected HBV patients, CD244 is highly coexpressed with PD-1 on both peripheral and liver-infiltrating T cells, demonstrating an inhibitory role for CD244 in the persistence of HBV infection.
The expression of these receptors on virus-specific IL-17-producing CD4+ and CD8+ T cells in CHB is unknown. If Th17 and Tc17 cells are regulated through these receptors, then checkpoint receptor inhibitors, which are currently being developed therapeutically, may exacerbate underlying inflammatory processes, possibly leading to the development of serious life-threatening conditions.
In this study, we have investigated the patterns of immunoinhibitory molecules on CD4+ and CD8+ T cells producing IL-17 or IFNγ in subjects infected with CHB compared with control subjects to determine the contribution of dysregulation of different immunoregulatory pathways to impaired T cell responses and their suitability for therapeutic targets in subjects with chronic HBV infection.
Patients and Methods
Study subjects
Ten patients with CHB were recruited from among patients attending the viral hepatitis clinic at Military Medical Academy, Sofia. None of the patients were receiving treatment at the time of enrollment into the study. The patients were negative for coinfection with hepatitis C, hepatitis delta, and human immunodeficiency virus. Ten HBV-negative subjects were recruited as healthy controls (HCs). The clinical characteristics of study participants are summarized in Table 1. The study conformed to ethical guidelines of the Declaration of Helsinki, as approved by the Ethics Committee of Military Medical Academy, Sofia. All patients gave written informed consent to participate in the study.
CHB, chronic hepatitis B; HBV, hepatitis B virus; N/A, not applicable.
Preparation of peripheral blood mononuclear cells and cell culture
Peripheral blood mononuclear cells (PBMCs) were isolated from heparinized blood using density centrifugation and cryopreserved as previously described (Cooksley and others 2008; Chokshi and others 2014). Before use, PBMCs were rapidly thawed and their viability determined by Trypan blue exclusion. The viability of all samples was found to be >90%.
To determine the frequency of IL-17-producing, IFNγ-producing, and noncytokine-producing HBV antigen-specific cells, 500,000 PBMCs were incubated in 200 μL of culture medium consisting of RPMI 1640 (Life Technologies, Paisley, UK) supplemented with 10% human AB serum (Sigma, Dorset, UK), 10 μg/mL recombinant hepatitis B core antigen (HBcAg) (American Research Products, Inc., MA), and 10 μg/mL recombinant hepatitis B surface antigen (HBsAg) (American Research Products, Inc.) for 72 h at 37°C with 5% CO2.
Control cultures were prepared where 500,000 PBMCs were cultured in 200 μL of RPMI supplemented with 10% human AB serum without antigens. After 72 h, the cell culture supernatants were removed and stored at −80°C for subsequent quantification of cytokine profiles. For HBV antigen-specific cultures, PBMCs were incubated for a final 6 h at 37°C with 5% CO2 in fresh culture medium supplemented with HBV antigens. Control cultures were incubated for a final 6 h in culture medium only. Brefeldin A was added to all cultures at a final concentration of 5 μg/mL for the last 5 h of incubation before immunophenotyping and intracellular cytokine detection by flow cytometry.
Immunophenotyping and intracellular cytokine staining
PBMCs were washed with phosphate-buffered saline (Life Technologies) supplemented with 1% fetal bovine serum (Life Technologies) (FACSwash) and incubated at 4°C for 20 min with a panel of antibodies to a variety of cell surface markers. The following fluorochrome-conjugated antibodies were used: α-CD3-APC-Cy7 and α-CD8-PE-Cy7 (BD Biosciences, Oxford, UK); α-CD244-FITC and α-PD-1-PerCP-efluor710 (eBiosciences, Hatfield, UK); and α-TIM-3-PE (R&D Systems, Abingdon, UK). After this time, the cells were washed with FACSwash and then fixed and permeabilized with CytoFix/CytoPerm buffer (BD Biosciences). PBMCs were washed with Permwash (BD Biosciences) and incubated at 4°C for 30 min with α-IFNγ-Horizon V500 (BD Biosciences) and α-IL-17-Alexa Fluor 647 (eBiosciences) antibodies. The fluorescently labeled PBMCs were analyzed on a BD FACSCanto II flow cytometer, and ∼200,000 lymphocytes were collected and analyzed using BD FACSDiva software (Becton Dickinson, Oxford, UK). The gating strategy we employed to identify cytokine producing T cells is presented in Supplementary Figure S1 (Supplementary Data available online at
Determination of cell culture supernatant cytokine profiles
The levels of a number of cytokines and chemokines were measured in cell culture supernatants. A Cytokine Bead Array® (BD Biosciences) was performed in accordance with the manufacturer's instructions to quantitate levels of IFNγ, IL-6, IL-8, IL-10, IL-12p70, IL-17A, and TNF-α. The levels of IL-23 and Galectin-9 were quantitated in the culture supernatants using commercially available ELISAs (R&D Systems, Abingdon, UK, and CUSABIO, Wuhan, PR China, respectively) in accordance with the manufacturer's instructions.
Statistical analyses
The independent samples t-test adjusted for variance and inequality was used for group comparisons. All statistical analyses were based on 2-tailed hypothesis tests with a significance level of P < 0.05.
Results
Frequencies of HBV-specific CD4+ and CD8+ cells producing IL-17 and IFNγ
The total number of CD4+ T cells producing IL-17 (Th17) was significantly higher in CHB patients than in control subjects (P = 0.03) and this difference was more pronounced in virus-specific Th17 cells (P = 0.001) (Fig. 1A). The number of virus-specific Tc17 cells was also significantly higher in CHB patients than in control subjects (P = 0.007), but not in the nonspecific Tc17 population (Fig. 1B). These data confirm the presence of HBV antigen-specific Th17 and Tc17 cells within the Th17 and Tc17 cell populations in CHB. As expected, the frequencies of HBV-specific CD4+ and CD8+ T cells producing IFNγ were significantly higher in CHB patients than control subjects (P = 0.008; P = 0.029, respectively) (Fig. 1C, D).

Frequency of total and HBV antigen-specific CD4+ and CD8+ T cells producing IL-17 or IFNγ in CHB patients compared with HC subjects. PBMCs from CHB and HC subjects were cultured with or without HBV antigens, and the frequency of T cells producing IL-17 or IFNγ was assessed by flow cytometry.
Expression of immune inhibitory markers on CD4+ and CD8+ T cells producing IL-17 or IFNγ
We analyzed the expression of PD-1, TIM-3, and CD244 on IFNγ-producing, IL-17-producing, and noncytokine-producing T cells in CHB patients and control subjects. Histograms from two representative subjects illustrating the expression of PD-1, TIM-3 and CD244 on differnt T-cell subsets are presented in Supplementary Figure S2. When we analyzed the expression of PD-1, TIM-3, and CD244 on noncytokine-producing cells, we saw that the magnitude of expression of all 3 of these molecules was lower than that observed on IFNγ and IL-17-producing CD4+ and CD8+ cells (Figs. 2 –4). We did not observe any differences in PD-1 expression on Th17 or Tc17 cells in CHB patients compared with control subjects (Fig. 2C, D).
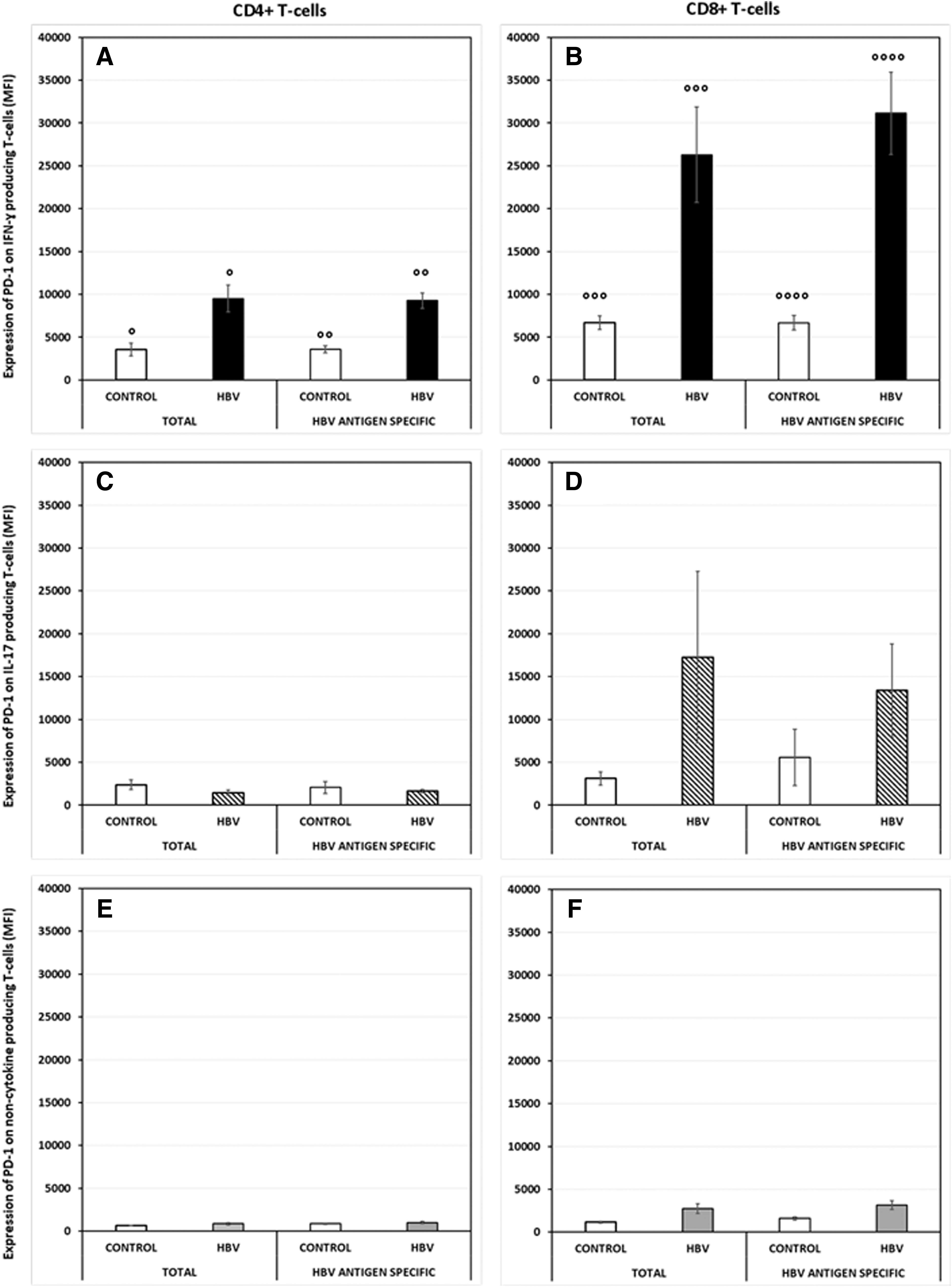
Expression of PD-1 on total and HBV antigen-specific CD4+ and CD8+ T cells producing IL-17 or IFNγ in CHB patients compared with HC subjects. PD-1 expression was assessed on PBMCs (from CHB and control subjects) that had been cultured for 72 h with or without HBV antigens.

Expression of TIM-3 on total and HBV antigen-specific CD4+ and CD8+ T cells producing IL-17 or IFNγ in CHB patients compared with HC subjects. PBMCs (from CHB and control subjects) that had been cultured for 72 h with or without HBV antigens and TIM-3 expression were assessed by flow cytometry.

Expression of CD244 on total and HBV antigen-specific CD4+ and CD8+ T cells producing IL-17 or IFNγ in CHB patients compared with HC subjects. CD244 expression was assessed on PBMCs (from CHB and control subjects) that had been cultured for 72 h with or without HBV antigens.
However, we saw significantly higher expression of PD-1 on both CD4+ and CD8+ IFNγ-producing cells (P = 0.004; P = 0.007) in CHB patients (Fig. 2A, B), supporting our previously reported findings. This difference was also observed on HBV-specific CD4+/CD8+ IFNγ+ cells (P = 0.0001; P = 0.001) (Fig. 2A, B). PD-1 expression on HBV-specific noncytokine-producing CD4+ (P = 0.0002) and CD8+ (P = 0.005) cells was significantly lower than that on IFNγ-producing cells (Fig. 2E, F).
Analysis of TIM-3 expression on CD4+/CD8+ T cells producing IFNγ in CHB patients revealed no difference (Fig. 3A, B). However, TIM-3 expression on Th17 cells from CHB patients was significantly lower compared with control subjects (P = 0.024) and this difference in expression was also observed on HBV-specific Th17 cells (P = 0.032) (Fig. 3C). TIM-3 expression was also significantly lower on Tc17 cells of CHB patients compared with control subjects (P = 0.023) (Fig. 3D); however, we did not observe any HBV-specific differences in these cells. As observed with PD-1, TIM-3 expression on HBV-specific noncytokine-producing CD4+ and CD8+ (both P = 0.0005) cells was significantly lower than that on IFNγ-producing cells (Fig. 3E, F).
Notably, CD244 expression was also different on HBV-specific CD4+ (P = 0.013) and CD8+ (P = 0.026) T cells producing IFNγ in CHB patients (Fig. 4A, B), but not in IL-17-producing T cells (Fig. 4C, D). CD244 expression on different T cell subsets in each individual subject are presented in Supplementary Figure S3. CD244 expression on noncytokine-producing CD4+ (P = 0.00003) and CD8+ (P = 0.012) cells was significantly lower than that on IFNγ-producing cells (Fig. 4E, F). These observations demonstrate that CD244 is coexpressed with PD-1 on HBV antigen-specific IFNγ-producing cells in CHB subjects, and like PD-1, this molecule does not appear to have a direct role in the regulation of IL-17-producing T cells in CHB infection.
Determination of cytokine profiles in cell culture supernatants
Secretion of IFNγ, IL-6, IL-8, IL-10, IL-12p70, IL-23, and TNF-α was significantly impaired in HBV antigen-stimulated PBMCs from CHB subjects compared with that of PBMCs from control subjects, reflecting the well-documented functional impairment observed in CHB infection. We observed elevated levels of galectin-9 secretion by HBV antigen-stimulated PBMCs compared with PBMCs cultured in medium alone; however, the difference observed was not significant. In HBV infection, increased galectin-9 expression is associated with viral persistence due to the expansion of T regulatory cells and dysfunction of T effector cells. We did not detect any IL-17A in the culture supernatants we tested, and we suggest that this may be because the total amount of IL-17A that would be produced by the relatively low frequency of IL-17-producing cells we detected in our cohorts would fall below the lower level of detection for the cytokine bead array assay (LLD 0.3 pg/mL).
Discussion
In this study, we reveal for the first time that inflammatory virus-specific T cells producing IL-17 express a different immune inhibitory checkpoint receptor profile from antiviral T cells producing IFNγ in CHB infection. Our results also demonstrate that in CHB, there is an increased frequency of HBV-specific CD4+ and CD8+ IL-17-producing T cells. High levels of viral antigens and ongoing replication drive the production of this inflammatory subset of T cells, which are known to augment disease progression (Yang and others 2013).
Currently, there is little data available on the mechanisms by which these proinflammatory Th17 and Tc17 cell functions are regulated. We have previously shown that HBV-specific IFNγ-producing T cells, which are critically important for mediating antiviral T cell responses, are functionally exhausted due to overexpression of PD-1 in CHB and in other liver diseases (Evans and others 2008; Markwick and others 2015).
The current study confirms this and also shows that PD-1 is not involved in the regulation of IL-17-producing T cells. We did, however, observe hypoexpression of the checkpoint receptor TIM-3 on total and HBV-specific Th17 cells and total Tc17 cells in CHB patients. Low TIM-3 expression on IL-17-producing T cells in CHB can induce exacerbated production of IL-17, leading to excessive inflammatory processes.
It has been previously reported that CD244 is highly coexpressed with PD-1 on both peripheral and liver-infiltrating T cells in patients with chronic HBV infection, demonstrating an inhibitory role for CD244 in the persistence of HBV infection (Raziorrouh and others 2010). In this study, we did not observe any differences in CD244 expression on Th17 or Tc17 cells. There is, however, a significantly higher expression of CD244 on HBV antigen-specific CD4+ and CD8+ cells producing IFNγ, demonstrating a pattern of coexpression with PD-1.
However, despite this observed parallel, the magnitude of PD-1 expression is much greater than that observed for CD244. The discrepancy could be explained by the findings of a study that investigated the relationship between PD-1, TIM-3, and CD244 expression and exhausted T cell functionality on cultured and ex vivo CD8+ T cells from HIV+ and HIV– donors (Pacheco and others 2013). They observed that despite displaying functional exhaustion, CD244 was downregulated on CD8+ T cells from HIV+, but not HIV–, subjects upon stimulation with cognate peptide. This observed downregulation only occurred upon simultaneous TCR signaling with CD244 crosslinking and internalization into an intracellular compartment since CD244 downregulation was not observed on PMA-ionomycin-stimulated CD8+ cells.
In antigen-stimulated cells, CD244 internalization occurred within hours of TCR stimulation and at lower peptide concentrations than that required to induce IFNγ production. The authors conclude that this rapid CD244 internalization 2-step mechanism plays a role in modulation of antiviral CD8 T cell responses. A similar phenomenon could play an important role in CD244 expression on the exhausted phenotype of HBV antigen-specific T cells. HBV antigen stimulation may lead to internalization of CD244 as observed in the HIV setting and thus account for the lower levels of membrane CD244 expression that we have observed in our study.
Additionally, it has been reported from a study investigating the role of CD244 in antiviral T and NK cell responses that in vitro CD244 blockade can result in either an inhibitory or activating effect on the antiviral T cell responses in PBMCs of subjects with different viral infections (Waggoner and Kumar 2012). In light of these observations, it is clear that further in vitro work to determine whether blockade of CD244 will restore or further inhibit functional anti-HBV immune responses is required to determine the applicability of CD244 blockade as a therapeutic strategy for chronic HBV infection.
It has been reported by several authors that checkpoint receptors, including PD-1, TIM-3, and CD244, are expressed at higher levels on intrahepatic antiviral HBV antigen-specific CD8+ T cells compared with those found in the peripheral compartment. These findings demonstrate that cells in locations of viral tropism possess a more exhausted phenotype and also that this phenotype is mirrored in the peripheral compartment (Peng and others 2008; Fisicaro and others 2010; Raziorrouh and others 2010). While findings of our current study are restricted to the periphery, there is precedence to suggest that our observations from the peripheral compartment would be a representative, if not subdued, reflection of the hepatic compartment.
Immune checkpoint receptors regulate the magnitude of immune responses to protect against collateral tissue damage during immune responses to infection and to maintain peripheral self-tolerance. They act as homeostatic regulators of T cell functionality where hyperexpression of the signature leads to impairment of T cell function and hypo- or underexpression leads to an activation of T cell immunity. Restoration of effective immune homeostatic regulation has been harnessed therapeutically in several tumors, including primary liver cancer, where the application of antibody to PD-1 can unmask antitumor T cell responses with much success (Topalian and others 2012a).
At the time of publication there are 5 anti-PD-1 antibodies and 3 anti-PDL-1 antibodies are being investigated in a number of clinical trials as potential new treatments for a variety of solid and hematological malignancies. (Brahmer and others 2012; Topalian and others 2012b). The use of an anti-PD-1 monoclonal antibody, Nivolumab, for treatment of hepatocellular carcinoma in uninfected patients and patients chronically infected with HBV or HCV has been investigated in the CheckMate 040 study. The results of this open label, noncomparative, dose escalation, and expansion Phase 1/2 study have shown encouraging objective response rates and overall survival with a manageable safety profile (El-Khoueiry and others 2017).
Since T cell exhaustion is common to both cancer progression and persistence of chronic viral infections, the use of immune checkpoint inhibitors could potentially be a useful therapeutic strategy to treat chronic viral infections such as HBV. Evaluation of Nivolumab in a cohort of chronic HCV patients has been described (Gardiner and others 2013). In this study, 15% of patients had a significant reduction in HCV RNA after receiving just a single dose of Nivolumab.
More recently, the data from a Phase 1 study evaluating anti-PD-1 therapy with or without a therapeutic vaccine (GS-4774) in HBeAg-negative
It is also worth noting that in this study, patients received 0.3 mg/kg Nivolumab, while in the CheckMate 040 study, hepatocellular carcinoma subjects chronically infected with HBV had manageable safety profiles when receiving 3 mg/kg Nivolumab. Together, the findings from these studies suggest that checkpoint receptor inhibition as a therapeutic strategy for
When utilizing immunotherapy to treat CHB infection, it will be important to ensure that employment of checkpoint inhibitors does not inadvertently activate IL-17-producing cells, or other inflammatory immune cells indirectly, in addition to restoration of vital antiviral IFNγ immune responses. The expression profiles of immune inhibitory molecules and their effect on the function of IL-17-producing cells will need to be evaluated to identify the most suitable candidates.
The results of our study suggest that blockade of the PD-1/PDL-1 inhibitory pathway would be an ideal therapeutic target for CHB infection. The use of a PD-1 antibody would restore effective antiviral IFNγ production without augmenting IL-17 production as our results demonstrate that PD-1 is only upregulated on IFNγ-producing and not IL-17-producing CD4+ and CD8+ T cells. However, targeting the TIM-3/galectin-9 pathway should be approached with caution as a treatment strategy for CHB infection as blockade of this pathway may potentially lead to increased IL-17 production and exacerbation of the inflammatory processes and severity of disease in CHB infection.
Footnotes
Acknowledgment
This work is supported by a grant from the Foundation for Liver Research.
Authors' Contributions
S.C. and H.C. conceived and designed the experiments and wrote the manuscript. H.C. and T.H-L. performed the experiments. H.C. and S.C. analyzed the data with statistical analysis from A.R. M.S. and K.K. contributed the clinical material. R.W. funded the research and edited the manuscript.
Author Disclosure Statement
No competing financial interests exist.