
Abstract
In this study we tried to explore whether calcitonin gene-related peptide (CGRP) regulates the potential antigen uptaking ability of human bronchial epithelial cells (HBECs) and promoting the differentiation of Th1/Th2. We found that CGRP increased the uptake of fluorescein isothiocyanate labeled ovalbumin (FITC-OVA) by HBECs using fluorescence microscopy and flow cytometry analysis. MTT assay showed that T cells proliferated in a dose-dependent manner in the presence of OVA-pretreated HBECs and CGRP inhibited the proliferation of T cells. CGRP decreased secretion of IFN-γ, while it had no influence on secretion of IL-4 by ELISA. Our data suggest that CGRP enhanced HBECs antigen uptake ability and inhibits HBECs induced T cells proliferation.
Introduction
B
Materials and Methods
Cells culture
16HBE14o- cell, a generous gift from Dr. Dieter C Gruenert, University of California San Francisco, is an immortalized HBEC line. The cells were cultured in DMEM/F-12 medium (Hyclone) supplemented with 10% newborn bovine serum (Hyclone), 100 U/mL penicillin, and 100 U/mL streptomycin (Hyclone). Cells were cultured at 37°C with 5% CO2. They were used when cells reached 80% confluence. The viability of HBECs was consistently >95% of the cells harvested.
Isolation of T cells
Peripheral blood was isolated from healthy donors by density gradient centrifugation using Ficoll–Hypaque lymphocyte separation media. The isolated lymphocytes were washed twice with PBS. The puried T lymphocytes obtained by CD3 separation column (P&D System). Cells were suspended in RPMI 1640 medium (Gibco) containing 10% fetal calf serum (FCS), then incubated in the culture flasks at 37°C a humidified incubator with 5% CO2.
Antigen uptake of HBECs by using fluorescence microscopy and FACS
After the cells were grown in 96-well round-bottom plates with 80% confluence, HBECs or HBECs pretreated with CGRP (final concentration 10−8 M) for 30 min were pulsed with fluorescein isothiocyanate labeled ovalbumin (FITC-OVA) (Molecular Probes) (0.1 mg/mL) at 37°C, harvested at various time points, and analyzed with fluorescence microscopy (OLYMPUS IX51, Japan). The cells were then washed twice with cold PBS containing 1% FCS and 0.01% NaN3 and analyzed by flow cytometry (FACS) for quantitative analysis (BD FACS Calibur). The fluorescence background (cells incubation with FITC-OVA at 0°C) was always subtracted. The results obtained are expressed as mean fluorescence intensity (MFI) values.
The proliferation of T cells by stimulation HBECs with antigen
HBECs (1.5 × 105) were cultured in 6-well round-bottom plates with 80% confluence and then pretreated with or without CGRP (final concentration 10−8 M) for 30 min to verify the cells were used as accessory cells for antigen presenting. OVA (Sigma) were added at the final concentrations of 0, 0.01, 0.1, and 1 mg/mL, respectively, and incubated at 37°C with 5% CO2 for 4 h and eluted. About 106 of fresh-isolated T cells were then co-cultured with HBECs for 24 h. The cells were centrifuged, resuspended, and removed to 96-well round-bottom plates. MTT method was carried out to determine the proliferation activities of T cells.
Cytokine ELISA
The contents of cytokines in the supernatants were determined by specific ELISA. IFN-γ and IL-4 were assayed according to the protocols (R&D Systems).
Statistical analysis
All data were expressed as mean ± SE. The numerical data were analyzed using unpaired Student's t-test. The differences between groups were analyzed using the 1-way ANOVA with Student-Newman-Keuls multiple comparison tests.*P < 0.05 or **P < 0.01 was considered as statistically significant.
Results
CGRP increased antigen uptaken ability of HBECs
HBECs were pulsed with FITC-labeled OVA. The dynamic antigen uptake function of HBECs, punctuation and green signals could be observed in 30 min with a maximal effect at 60 min (Fig. 1). In order to quantitatively analyze the effects of CGRP on HBECs antigen uptake, MFI of HBECs was measured by FACS. We observed that the MFI of HBECs treated with CGRP was higher than untreated cells (P < 0.01) (Fig. 2A, B).

Uptake of antigen by HBECs. HBECs were grown on 96-well flat-bottomed plates, pulsed with FITC-labled OVA, harvested at different time points, and analyzed by fluorescence microscope.
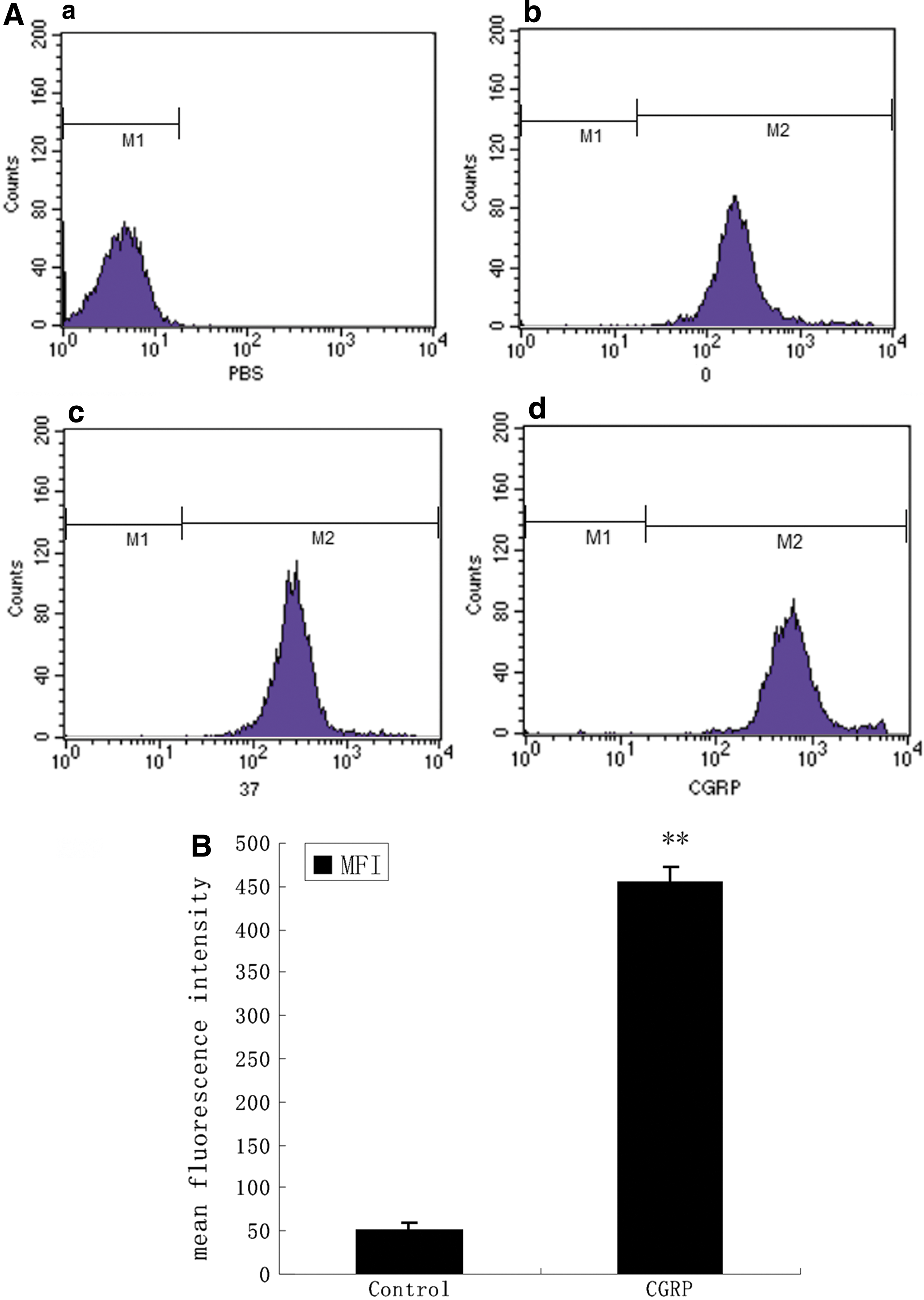
Effects of CGRP on uptake of FITC-OVA by HBECs.
CGRP decreased the ability of HBECs of promoting T cells proliferation
We have found that HBECs could not activate T cells proliferation in the absence of OVA. HBECs induced the proliferation of T cells by pretreatment with OVA in a dose-dependent manner (Figs. 3 and 4). However, HBECs pretreated with CGRP decreased T cells proliferation in the presence of OVA (Fig. 5).

The capacity of HBECs to serve as APCs for presenting antigen to stimulate T cell proliferation. Bars represent proliferation index. The proliferation of T cells was not enhanced when they were stimulated by HBECs without OVA used as antigen, and it was almost the same with background values (1). T cells did not proliferate in the presence of OVA as antigen only, either. When HBECs were used as antigen presenting cells to stimulate T cells in the presence of OVA, T cells showed enhanced proliferation. **P < 0.01 versus 1; ## P < 0.01 versus 2; ++ P < 0.01 versus 3. APC, antigen presenting cells.

Effects of different doses of OVA on T cells proliferation response. HBECs pretreated with different concentration of OVA on T cells proliferation. Results are expressed as OD value, bars represent mean ± SEM, are representative of 1 experiment using 3 different donors.

Effects on the capability of T cells proliferation by CGRP on HBECs as APCs. After stimulation with CGRP for 30 min, HBECs incubated With FITC-OVA and T cells, Bars represent mean ± SEM, are representative of at least 3 different experiments (n = 6, **P < 0.01 versus CGRP).
Effects of CGRP on the cytokines secreted by HBECs
After activation by antigen, naive CD4+ T cells differentiate into Th1 or Th2 effector cells. To identify the differentiation pathway that is predominantly affected by CGRP, we analyzed the cytokines secreted by HBECs that may act on Th1 and Th2 differentiation. In the supernatants of HBECs pretreated with CGRP. We detected concentrations of both cytokine IFN-γ and IL-4 were increased in the presence of OVA and T cells. CGRP treatment of HBECs decreased the amount of IFN-γ while it had no influence on secretion of cytokine IL-4 (Fig. 6).

IFN-γ and IL-4 secretions were determined by ELISA. Results are the mean ± SEM of 3 independent experiments performed in duplicate. HBECs pretreated with CGRP decreased the secretion of IFN-γ
Discussion
Asthma is a common respiratory disease characterized by airway inflammation and airway hyperreactivity. The balance of Th1/Th2 appears to play an important role in the immune regulation of respiratory system. Many studies showed that Thl/Th2 immune imbalance and the activation of Th2 play a very important role in asthma pathogenesis. “Neurogenic theory” was also put forward, and many researchers began to pay attention to CGRP, which was shown to participate in this pathogenesis. Preliminary study confirmed that CGRP can regulate the expression of HBECs antigen presenting molecules (Peng and others 2008). Its impact on APC has attracted attention, too.
Uptaking antigen is the precondition of antigen presenting of APC
Garry examined the ability of human small airway epithelial cells (SAECs) to recognize and ingest apoptotic human eosinophils. Cultured SAEC ingested apoptotic eosinophils but not freshly isolated eosinophils (Walsh and others 1999).
The ability of SAEC to ingest apoptotic eosinophils was enhanced by IL-1a or TNF-r in a time- and concentration-dependent fashion (Sexton and others 2004). Erez demonstrated that a physiologically relevant Ag (RW) is endocytosed by airway epithelial cells (AECs) and follows a class II processing pathway (Oei and others 2004). Salik and others (1999) had investigated accessory cell function, antigen trafficking, and uptake of immune complexes in isolated nasal epithelialcells (NEC), AECs, and in the 2 respiratory epithelial cell lines A549 and BEAS-2B. The kinetics of antigen uptake by the respiratory epithelial cells was dramatically increased by IFN-γ and GM-CSF.
We also confirmed HBECs can uptake FITC-OVA (Liu and others 2012). The effects of regulatory peptides including CGRP on the uptake of antigen by the AECs were rarely studied. By using flow cytometry as a quantitative analysis, we discovered that CGRP enhanced HBECs uptaking ability of FITC-OVA. It was shown that CGRP can improve the function of accessory cells and help to clear the pathogens and induce the inflammation effect, which has a protective effect on the body when the AECs encounter external antigen stimulation.
Bronchial epithelium lines the mucosal surface of airways, forming a mechanical barrier that separates the external environment from the internal milieu (Kurosawa and others 2003; Kim and others 2005). Previous studies showed that respiratory epithelial cells can stimulate T cell proliferation in a mixed lymphocyte reaction and can induce receptor-associated tyrosine kinase activation in cocultured T cells (Kalb and others 1997; Liao and others 2012). The results of our experiments confirmed that the proliferation of T cells is achieved through the HBECs uptaking and processing antigen, and the proliferation of T cells is closely related to the concentration of OVA. Taken together, HBECs can indeed act as APCs.
Little is known about the effects of CGRP on antigen presenting action of HBECs. CGRP-pretreated dendritic cell (DC) reduced the activation and proliferation of antigen-specific T cells and increased the numbers of T regulatory cells. The transfer of CGRP-pretreated DC diminished allergic airway inflammation in vivo, CGRP inhibits DC maturation and allergen-specific T cell responses, which affects the outcome of the allergic airway inflammation in vivo (Rchlitzer and others 2011). The predominant Th2 reaction plays an important role in the pathogenic machanism of asthma (Penaloza-MacMaster and others 2014). IFN-γ and IL-4 are a couple of cytokines that can not only agonist each other but also promote each other, respectively (Chen and others 2007). IL-4 is the characteristic cytokine of Th2 while IFN-γ is the characteristic cytokine of Th1 (Tang and others 2007). Tokoyoda and others (2004) showed that CGRP downregulates IL-2 and IFN-γ production and upregulates IL-4 production to promote Th2 differentiation by moderate activation of the cAMP/PKA pathway via the CGRP receptor in the presence of a CD3/CD28 costimulation signal. We observed that CGRP upregulated the expression of HLA-DR, downregulated CD80 on HBECs, enhanced antigen uptake of HBECs, inhibited the proliferation of T cells, and decreased the secretion of Th-1 type cytokines IFN-γ from HBECs. Although CGRP upregulated the expression of HLA-DR and enhanced antigen uptake of HBECs, it downregulated CD80 expression. These data illustrate an important role of the second signal in costimulatory molecules. According to the classical immunology theory, CD80 can induce Th0 cells to differentiate to Th1, and CD86 induce cells Th0 to differentiate to Th2 (Zhou and others 2009). CGRP decreased IFN-γ, which may be related to the downregulated expression of CD80.
In conclusion, our results unequivocally verify that HBECs express full antigen-presentation machinery. CGRP can modulate the differentiation of Th1/Th2 cells after antigens stimulation. In view of the complexity of the process of antigen presentation, this study in which cultured HBECs were used can only reflect the effects of CGRP in vitro. Further study needs to be carried out to fully understand the regulatory effect of CGRP.
Footnotes
Acknowledgments
The authors thank the Center of Medical Functional Experiment of Central South University for kind assistance. This work was supported by the Scientific Research Foundation of Hunan Provincial Science Technology Department (2014 FJ3009) and Scientific Research Fund of Hunan Provincial Education Department (14C1140).
Author Disclosure Statement
No competing financial interests exist.