
Abstract
Abnormal alveolar formation and excessive disordered elastin accumulation are key pathological features in bronchopulmonary dysplasia. Transforming growth factor (TGF)-β is an important regulator of the extracellular matrix in the developing lung. To determine if increased TGF-β would injure alveolar development by activating TGF-β signaling and by influencing the expression of elastogenesis-related protein, we performed intraperitoneal injection of newborn mice with the TGF-β-neutralizing antibody 1D11 and observed whether 1D11 had a protective role in the oxygen (O2)-exposed newborn mouse lung. The newborn mice were exposed to 85% O2 for 14 and 21 days. 1D11 was administered by intraperitoneal injection every day from postnatal days 3 to 20. Alveolar morphology was assessed by hematoxylin and eosin staining. The expression and distribution of elastin were evaluated by immunohistochemistry. The level of TGF-β signaling-related proteins were measured by immunohistochemistry, enzyme-linked immunosorbent assay, and Western blot. The expression levels of elastogenesis-related proteins, including tropoelastin, fibulin-5, and neutrophil elastase (NE), which participate in the synthesis, assembly, and degradation of elastin, were detected by real-time PCR and Western blot. In this research, impaired alveolar development and elastin deposition as well as the excessive activation of TGF-β signaling were observed in the newborn mouse lung exposed to hyperoxia. 1D11 improved alveolarization as well as the distribution of elastin in the newborn lung with hyperoxia exposure. The expression levels of tropoelastin, fibulin-5, and NE, which are important components of elastogenesis, were decreased by treatment with 1D11 in the injured newborn lung. These data demonstrate that 1D11 improved alveolarization by blocking the TGF-β signaling pathway and by reducing the abnormal expression of elastogenesis-related proteins in the O2-exposed newborn mouse lung. 1D11 may become a new therapeutic method to prevent the development of bronchopulmonary dysplasia.
Introduction
Bronchopulmonary dysplasia (BPD) is a chronic lung disease that most often occurs in premature infants exposed to high oxygen (O2) concentrations and mechanical ventilation (Vicencio and others 2004; Northway and others 1967). BPD is currently defined as the need for O2 supplementation for 28 days of life, with high rates of severe sequelae and even death (Bhandari 2014).
Transforming growth factor (TGF)-β is a member of a family of multifunctional growth factors involved in normal alveolar development (Lee and others 2006). The concentration of TGF-β in bronchoalveolar lavage fluid of patients with BPD is increased and correlates with the severity of BPD (Kotecha and others 1996). In our previous study, we found increased expression of TGF-β and activation of the TGF-β signaling pathway in the mouse lung exposed to hyperoxia (Han and others 2015). We speculate that inhibition of TGF-β expression may have a protective role in the O2-exposed newborn mouse lung and may become a new therapy for treating BPD. 1D11 is a TGF-β-neutralizing antibody that effectively controls the elevated expression level of TGF-β protein. The current research on 1D11 has focused more on its protective function against BPD (Alejandre-Alcázar and others 2007), cancer (Xin and others 2014), and radiation-induced lung injury (Anscher and others 2006). The effect of 1D11 in pup lungs with chronic exposure to hyperoxia has rarely been reported (Alejandre-Alcázar and others 2007), and the mechanisms of its action are different.
Some studies have indicated that the protective mechanism of 1D11 in the injured developing lung is through enhancing the expression of nitric oxide signaling enzymes (Bachiller and others 2010). Elastogenesis is a multistep process that involves the formation, assembly, and degradation of elastin (Yanagisawa and others 2009). Excessive disordered accumulation of elastin leads to air space enlargement and BPD (Kuang and others 2003). Disharmonious expression of elastogenesis-related protein can lead to the abnormal distribution of elastin (Camoretti-Mercado and Solway 2005; Bland and others 2007). TGF-β signaling participates in the regulation of the extracellular matrix by enhancing its synthesis and inhibiting the degradation of its components (Takemasa and others 2012).
In our research, we explored the role of 1D11 in a BPD model caused by hyperoxia and tested whether the mechanism of action of 1D11 is connected with the disordered expression of elastogenesis-related protein.
Materials and Methods
Animal experimental design
All the animal studies were conducted in accordance with protocols approved by the institutional review board and the animal use and care committee of Chongqing Medical University. Postnatal 24 h C57BL/6 mice weighing 1.32 ± 0.10 g were obtained from the experimental animal center of Chongqing Medical University. Within 24 h of delivery, littermates were randomly divided into four groups: the air-exposed control group (air group), the hyperoxia-exposed group (O2 group), the hyperoxia-exposed phosphate-buffered saline (PBS)-treated group (O2 + PBS group), and the hyperoxia-exposed 1D11-treated group (O2 + 1D11 group). The mice in the O2 group, the O2 + PBS group, and the O2 + 1D11 group were exposed to 85% O2 and were fed by the same mother, whereas the air control group mice were fed under normal air in the same room. The maternal mice were rotated daily between the O2 groups and the air groups to avoid the influence of maternal mouse O2 toxicity on the nursing pups.
The TGF-β-neutralizing antibody 1D11 (R&D Systems, Minneapolis, MN), at a total of 0.2 μg/g body weight in 5 μL/g body weight, was administered by intraperitoneal injection in the O2 + 1D11 group mice every day from postnatal (P) days 3 to 20. The O2 + PBS group mice were treated with the same amount of PBS at the same times.
Tissue collection and lung morphometry
At postnatal days 14 and 21, selected animals from each group were euthanized using carbon dioxide, and their lung tissues were obtained. The right lobe was stored at −80°C for enzyme-linked immunosorbent assay (ELISA), quantitative real-time PCR (qPCR), and Western blotting. The left lobe was fixed in 4% paraformaldehyde for hematoxylin and eosin (H&E) staining and immunohistochemistry. The fixed tissues were dehydrated and embedded in paraffin. The paraffin blocks were serially sliced in the coronal plane at 4 μm septum thickness. Serial sections were then stained with H&E to quantify alveolar structures on a motorized microscope stage. Mean linear intercept (MLI) and radial alveolar counts (RACs) were measured (Han and others 2015). MLI was detected by a cross curve drawn in the center of the field and was used to calculate the ratio of the cross curve length to the intersection number of the cross curve and alveolar septum. RAC was used to count the alveolar number by a straight line drawn from the center of a respiratory bronchiole to the nearest interlobular septum. The images were randomly acquired from 10 nonoverlapping fields per slide, with three slides per animal and four animals per group.
Immunohistochemistry of elastin and TGF-β1
Serial 4-μm-thick sections were deparaffinized by xylene and rehydrated by successive incubations in 100%, 95%, 80%, and 70% ethyl alcohol. Deparaffinized and rehydrated sections were microwave antigen retrieved by citric acid buffer (pH 6.0) and endogenous peroxidase quenched with 3% hydrogen peroxide for 15 min. After blocking with 5% bovine serum albumin (BSA) (Sigma, USA), the sections were incubated with primary antibodies anti-elastin (1:100; Abcam, UK) and TGF-β1 (1:200; GeneTex, USA) diluted in PBS overnight at 4°C. Immune complexes were detected with the appropriate secondary antibody conjugated to horseradish peroxidase (HRP), and the HRP activity was visualized using the Liquid DAB Plus Substrate Kit (Life Technologies, Carlsbad, CA). The slides were analyzed using a Nikon 55I microscope with a DS-Filc camera and NIS-Elements F software. The images were randomly acquired in 10 nonoverlapping fields per slide, with three slides per animal and six animals per group.
ELISA for active TGF-β1
At 14 and 21 days after delivery, the lung tissues were cut into pieces and homogenized with a protease inhibitor (Roche) using the method as described previously (Han and others 2015). The protein concentrations in the supernatants were measured by a BCA Assay Kit (Thermo Scientific, Rockford, IL). Equivalent protein concentrations of active TGF-β1 were quantified using a commercial ELISA Kit (MB100B; R&D Systems). The ELISA was performed according to the manufacturer's instructions. All the samples were analyzed in duplicate as a single batch and were run with control standards. The experiment was repeated three times.
Quantitative real-time PCR
qPCR was performed in the core laboratory of the Children's Hospital of Chongqing Medical University. The total RNA was isolated from the dissected lung tissues using the TRIzol Reagent (Invitrogen, USA) according to the manufacturer's instructions. The RNA was reverse transcribed into complementary DNA using the commercial reverse transcription reagent kit (TaKaRa, Japan) and amplified on a 7300 Real-Time PCR system (Applied Biosystems, Foste, CA) with TaqMan primers (Applied Biosystems). qPCR, using proprietary primers and probes (Taqman Gene Expression Assays; Applied Biosystems), was used to measure the messenger RNA (mRNA) expression of tropoelastin, fibulin-5, and neutrophil elastase (NE) with the primers shown in Table 1 (Life Technologies). β-actin was used as an internal control gene. In our experiments, we discovered that β-actin mRNA was not significantly differently expressed in various fetal and newborn mouse lung samples. Negative controls included reactions without templates and/or enzymes. The relative quantification method was used to determine the Ct values for the PCR products of the target genes and β-actin. The relative quantification formula was as follows: 2−ΔCt × 100%, where ΔCt = Ct(target gene) − Ct(β-actin).
Mouse Primer Sequences for Quantitative Real-Time PCR
Western blotting
The frozen right lobes of the mice were minced and homogenized in RIPA buffer (Thermo Scientific) containing a protease inhibitor cocktail (Thermo Scientific). The tissue lysates were centrifuged at 14,000g at 4°C for 5 min, and the supernatants were collected. The protein concentration was determined using a BCA Assay Kit (Thermo Scientific). Equivalent amounts of protein were separated by sodium dodecyl sulfate polyacrylamide gel electrophoresis and electrotransferred to a polyvinylidene fluoride (PVDF) Membrane (Millipore, USA). The membrane was blocked for 1 h at room temperature with 5% BSA (Sigma). The membranes were incubated overnight at 4°C with specific diluted primary antibodies, including anti-TGF-β1 (1:600; GeneTex), anti-Smad2/3 (1:1000; Cell Signaling Technology, USA), anti-P-Smad2/3 (1:500; Cell Signaling Technology), anti-tropoelastin (1:500; Abcam, USA), anti-fibulin-5 (1:500; Proteintech Group, USA), and anti-NE (1:500; Abcam). After three washes with TBST, the membrane was incubated with an appropriate HRP-conjugated secondary antibody (goat anti-rabbit IgG; sc-2301, Santa Cruz, CA) diluted in TBST for 1 h at room temperature.
After washing, the subsequent analysis of the protein was performed with chemiluminescence using an electro-chemi-luminescence (ECL) Western Blotting Kit (Millipore) following the manufacturer's recommendations. The relative intensities of the bands were evaluated using Kodak 1D software, version 3.5.4 (Kodak Scientific Imaging system, Rockville, MD). All the experiments were repeated at least three times.
Statistical analysis
A two-way analysis of variance multiple comparison tests was used to identify differences in both the mRNA expression and protein levels, as well as the quantitative histological measurements. The statistical analyses were performed using the Prism software package, version 5.01 (GraphPad, San Diego, CA). P value <0.05 was considered statistically significant.
Results
The TGF-β signaling pathway was successfully blocked by 1D11 treatment in the newborn mouse lung exposed to hyperoxia
TGF-β is the basic factor in the TGF-β signaling pathway (Tang and others 2014). Increased TGF-β can lead to activation of the TGF-β signaling pathway (Bartram and Speer 2004). We, therefore, assessed the expression of TGF-β1 protein by immunohistochemistry at days P14 and P21 (Fig. 1a–h). There was a difference in alveolar size and alveolar septum thicknesses among each group of mouse lungs; thus, we determined the expression of TGF-β1 by measuring the mean optical density (immunohistochemical image gray-scale analysis of the results of the integral optical density [IOD]/area).
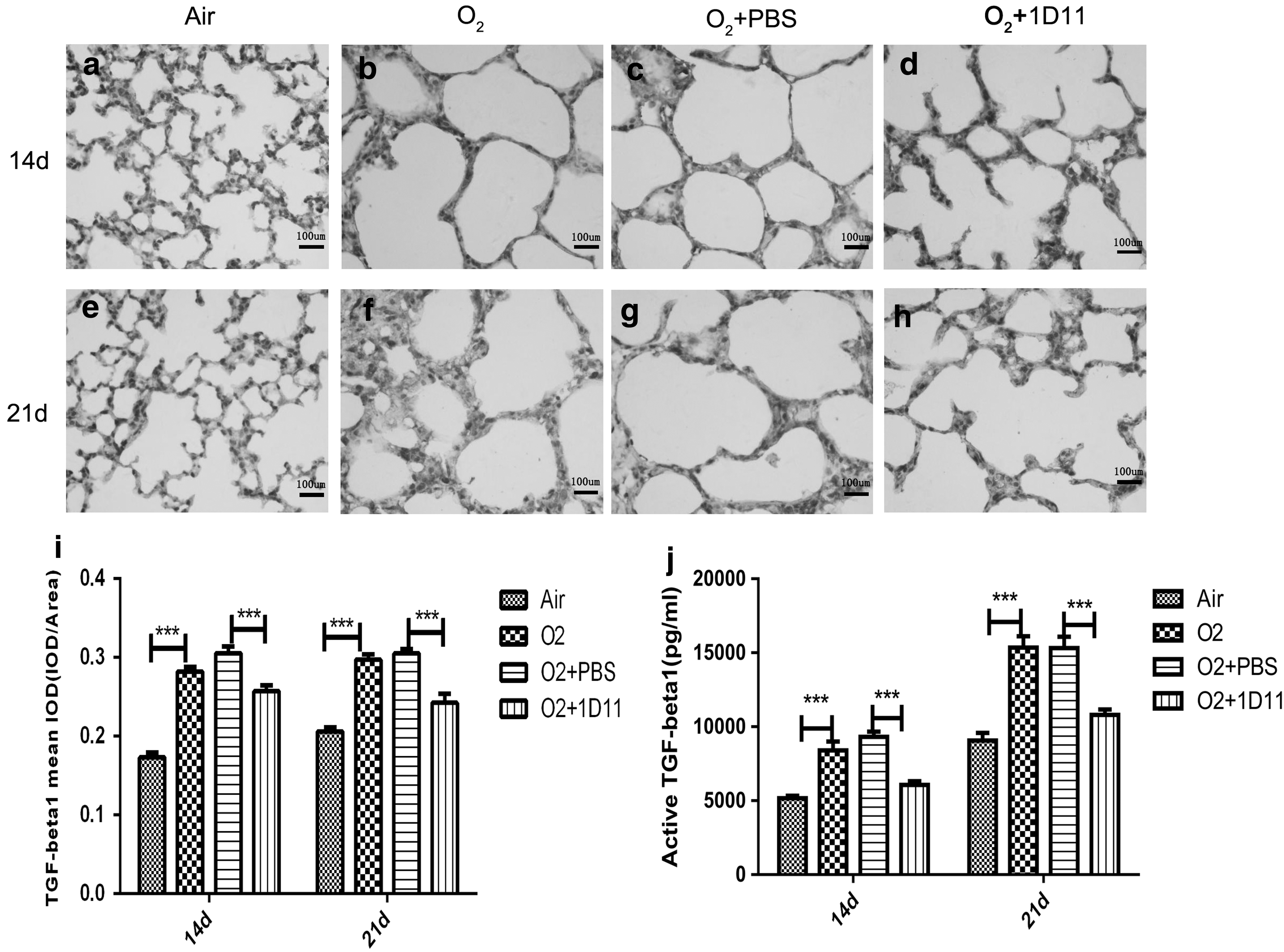
1D11 inhibited the overexpression of TGF-β1 protein in the newborn mouse lung exposed to hyperoxia. The amount and location of TGF-β1 on days 14 and 21 in injured mouse pup lungs were detected by immunohistochemistry (n = 6 animals, 12 fields/animal). One representative slide per group is shown
The results showed that TGF-β1 was located in the alveolar epithelium in the air control mouse lungs and that the expression of TGF-β1 was increased and distributed all over the alveolar wall in the newborn mouse lung exposed to hyperoxia (Fig. 1i). Treatment with 1D11 significantly decreased the expression and distribution of TGF-β1 (Fig. 1i). Our results showed that 1D11 succeeded in inhibiting the expression of TGF-β1 protein in the hyperoxia-exposed newborn mouse lung.
TGF-β binds to the constitutively active type II receptor, which then recruits and phosphorylates the type I receptor (Wang and others 2015). To evaluate active TGF-β1 expression in the lung tissue, we detected the expression of active TGF-β1 at 14 and 21 days by ELISA. Compared with the air control group, the expression of active TGF-β1 was significantly increased in the O2 group mouse lung (Fig. 1j). After intervention with 1D11, we observed that the expression of active TGF-β1 was decreased (Fig. 1j). 1D11 decreased the expression of active TGF-β1 in the newborn mouse lung after hyperoxia exposure.
The expression level of phosphorylated Smad2,3 (P-Smad2,3) protein can be used as an index to judge whether TGF-β signaling is activated (Camoretti-Mercado and Solway 2005). To assess whether the activity of the TGF-β signaling pathway was influenced by hyperoxia and 1D11, we investigated the protein expression of TGF-β1 and Smad2,3, as well as the phosphorylation of Smad2,3, at 14 and 21 days by Western blot. As shown in Fig. 2, the protein level of TGF-β1 in the O2 group newborn lungs was higher than that in the air control group (Fig. 2a). However, compared with the mice in the O2 + PBS group, the protein level was significantly decreased in the mouse lungs that had received treatment with 1D11 (Fig. 2b). The level of Smad2,3 protein was markedly decreased in response to hyperoxia (Fig. 2c). After intervention with 1D11, the expression of Smad2,3 was increased (Fig. 2c). The P-Smad2,3 expression was observed at a higher degree in the mouse lung exposed to hyperoxia than that exposed to air (Fig. 2d). 1D11 treatment significantly decreased the protein level of P-Smad2,3 in the hyperoxia-exposed newborn lung (Fig. 2d). These findings indicate that the TGF-β signaling pathway can be activated by hyperoxia and that 1D11 may succeed in blocking the TGF-β signaling by suppressing the expression of the TGF-β1 protein.
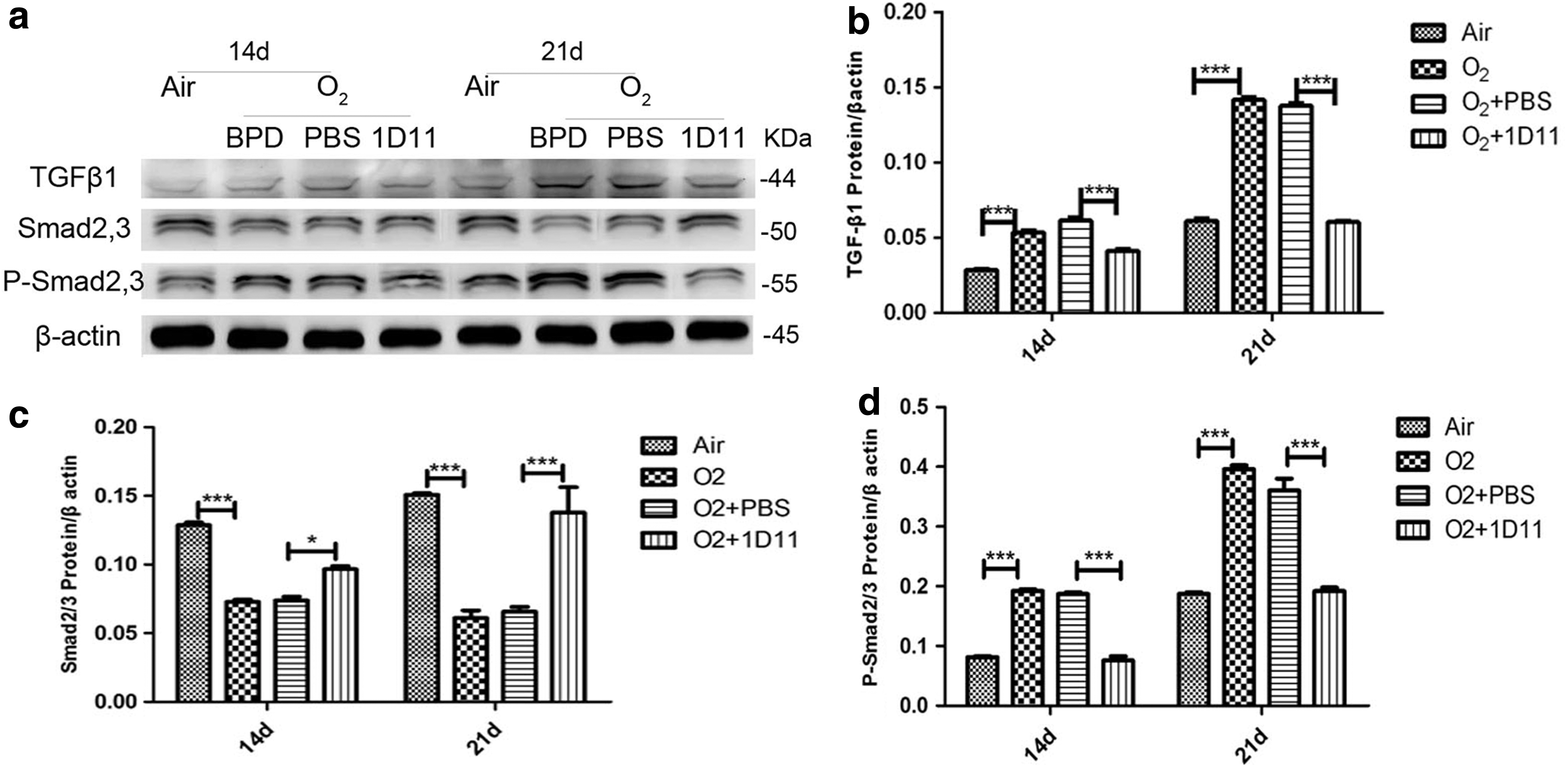
1D11 was successful in blocking the TGF-β signaling pathway in the hyperoxia-exposed newborn mouse pups lung. To assess the activity of TGF-β signaling, the expression levels of TGF-β signaling-related proteins, including TGF-β1, Smad2/3, and phosphorylated Smad2/3, were measured by Western blot at 14 and 21 days. Representative examples from a minimum of three independent experiments are shown
Hyperoxia-induced impairment of lung development was improved by ID11 treatment
The P14- and P21-day lung histopathology of the air control group lungs showed that the alveolar structure was normal. However, marked impairment of alveolarization characterized by fewer and larger alveoli was observed in the O2 group newborn lungs (Fig. 3a, b, e, f). Compared with the O2 + PBS group, the O2 + 1D11 group improved the development of alveoli (Fig. 3c, d, g, h). To evaluate the effect of hyperoxia and ID11 on lung development, assessment of RAC and MLI was performed. RAC evaluated the number of alveoli, and the quantification of alveolar size was calculated by MLI measurement. The RAC value of the O2 group was significantly decreased compared with the air control group (Fig. 3i), but the value of MLI was significantly increased in the O2 group compared with the air control group (Fig. 3j).
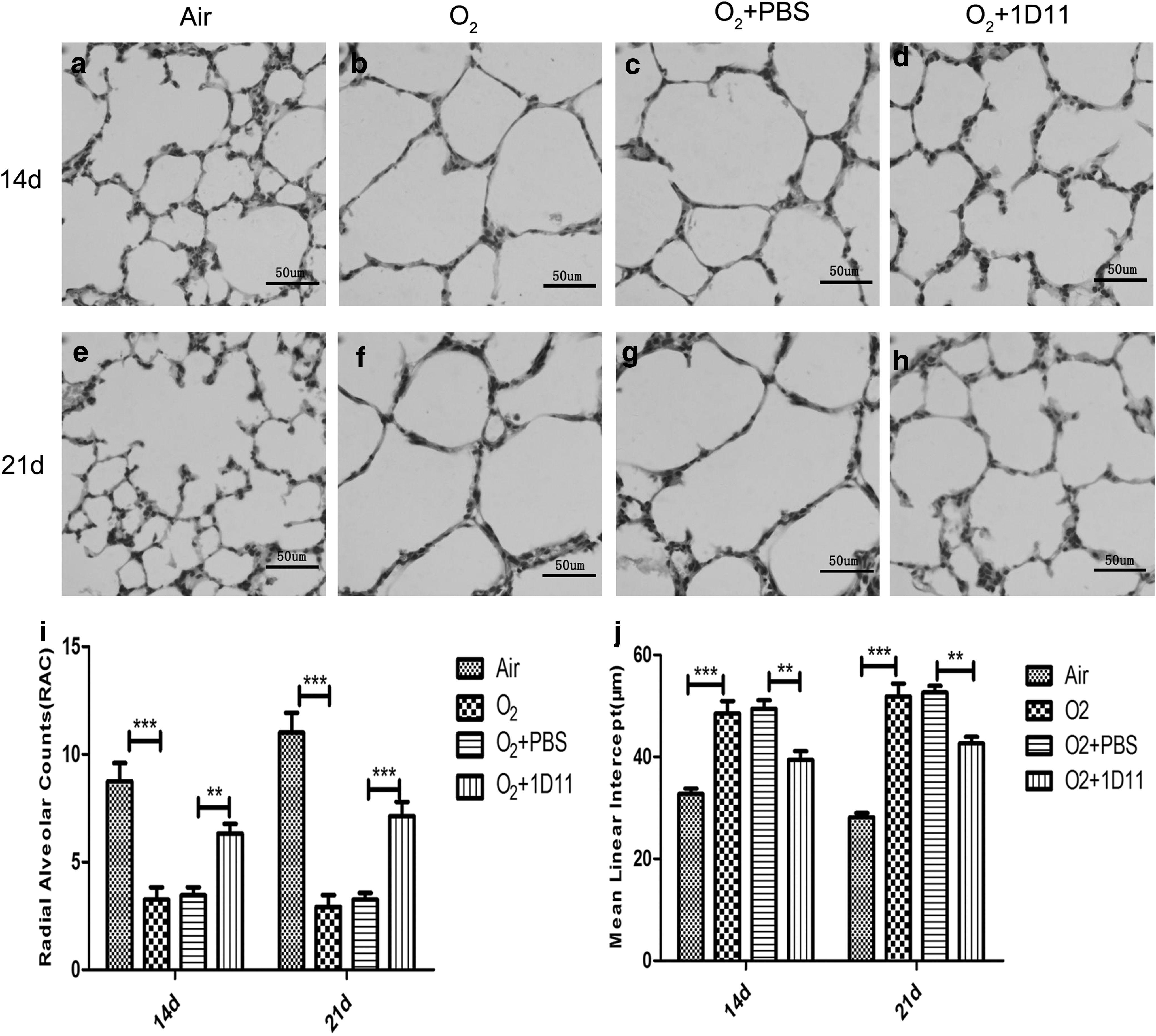
1D11 ameliorated alveolar development in the impaired newborn mouse lung. Lung morphology was observed by hematoxylin–eosin staining (n = 4 animals, 12 fields/animal). One representative slide per group is shown. Bar = 50 μm. Hyperoxia markedly inhibited alveolarization at 14 days
The pathological characteristics of the O2 group were consistent with alveolar simplification, which was the typical feature of BPD (Witsch and others 2014a). In comparison with the O2 + PBS group, the RAC value of the O2 + 1D11 group was significantly increased (Fig. 3i), and the MLI value of the O2 + 1D11 group was significantly decreased (Fig. 3j). These results indicate that 1D11 can improve hyperoxia-induced impairment of alveolar development.
The body weight of newborn mice exposed to hyperoxia was improved by ID11 treatment
Because alveolarization in the damaged developing lung was improved by ID11 treatment, we further determined whether 1D11 could ameliorate the reduced body weight in hyperoxic-injured newborn mice. The body weight data of each group of mice at postnatal days 1, 3, 7, 14, and 21 were collected and analyzed using statistical software. The mice were all from the same batch, and the experiment was repeated at least three times. Compared with the air control group mice, body weight was significantly decreased in mice exposed to O2 (Fig. 4). After intraperitoneal injection with 1D11, the body weight of hyperoxia-exposed mice was notably increased. However, 1D11 was not sufficient to return the body weight to normal (Fig. 4). The index could in part reflect the growth and development of newborn mice. Hyperoxia inhibited growth development in pups, and 1D11 improved neonatal mouse development.
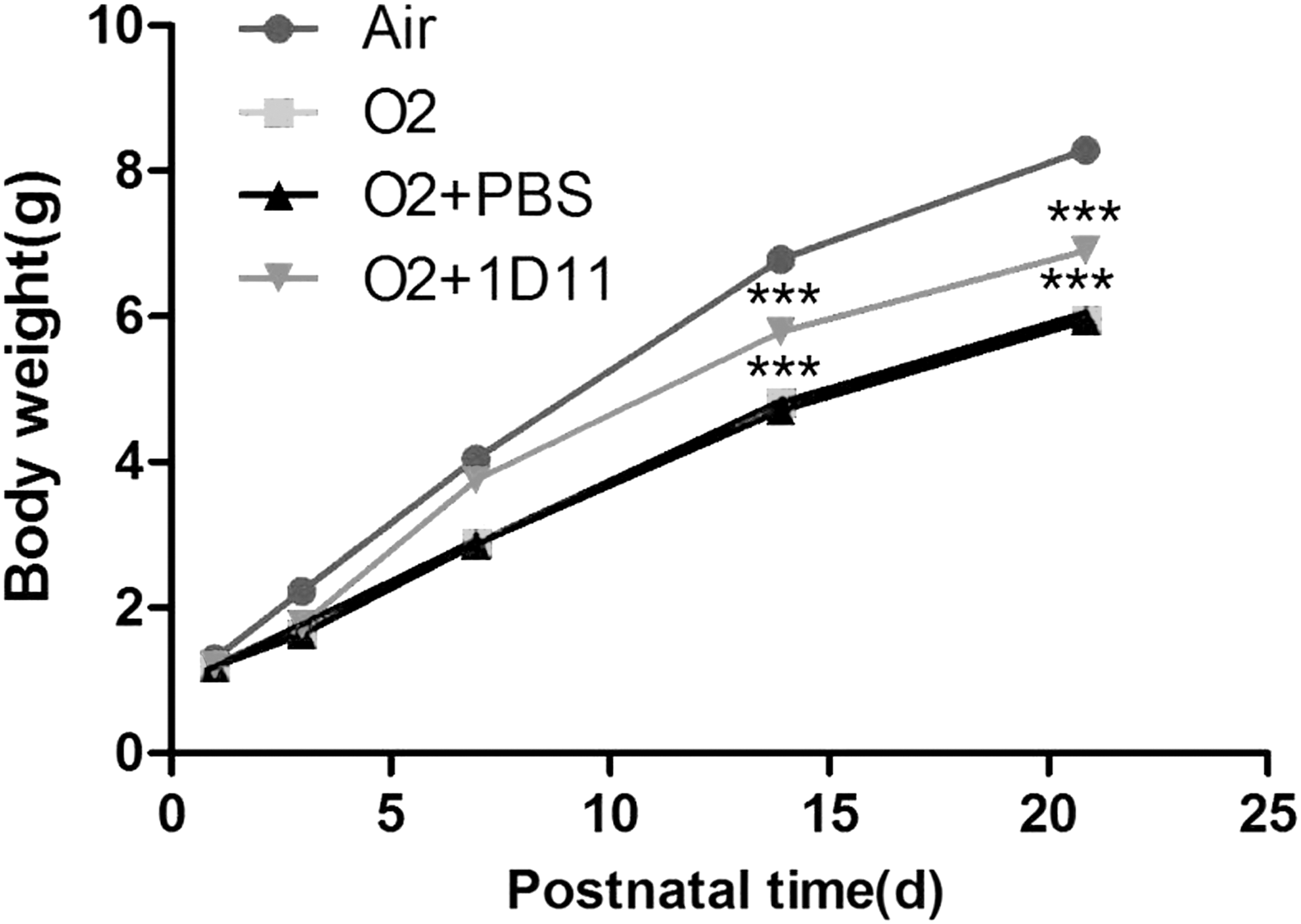
1D11 contributes to improvements in the postnatal growth and development of mice. Time course of body weight for the pups treated as indicated. The data represent 10–15 mice per group at each time point. Compared with the air-exposed control, the body weight of O2-exposed mice was reduced. Compared with the 85% O2-exposed mice treated with PBS, the body weight of mice that were treated with 1D11 was increased. ***P < 0.001.
Aberrant expression and distribution of elastin in the hyperoxia-exposed mouse lung was improved by 1D11 treatment
We evaluated the localization and expression of elastin by immunohistochemistry. There was a difference in alveolar size and alveolar septum thicknesses among each group; thus, we determined the expression of elastin by measuring the mean optical density (IOD/area). As shown in Fig. 5, elastin was localized in the top of the distal airway in the air control pup lungs at days P14 and P21. However, the staining of elastin was observed in O2 group pup lungs throughout the walls of the alveoli and were significantly increased compared with the air control pup lung (Fig. 5b, f). After injection with 1D11, the lung sections obtained from the mice displayed reduced expression of elastin (Fig. 5c, d, g, h), and the distributed expression of elastin was improved. These consequences suggested that 1D11 improved the disordered accumulation of elastin.
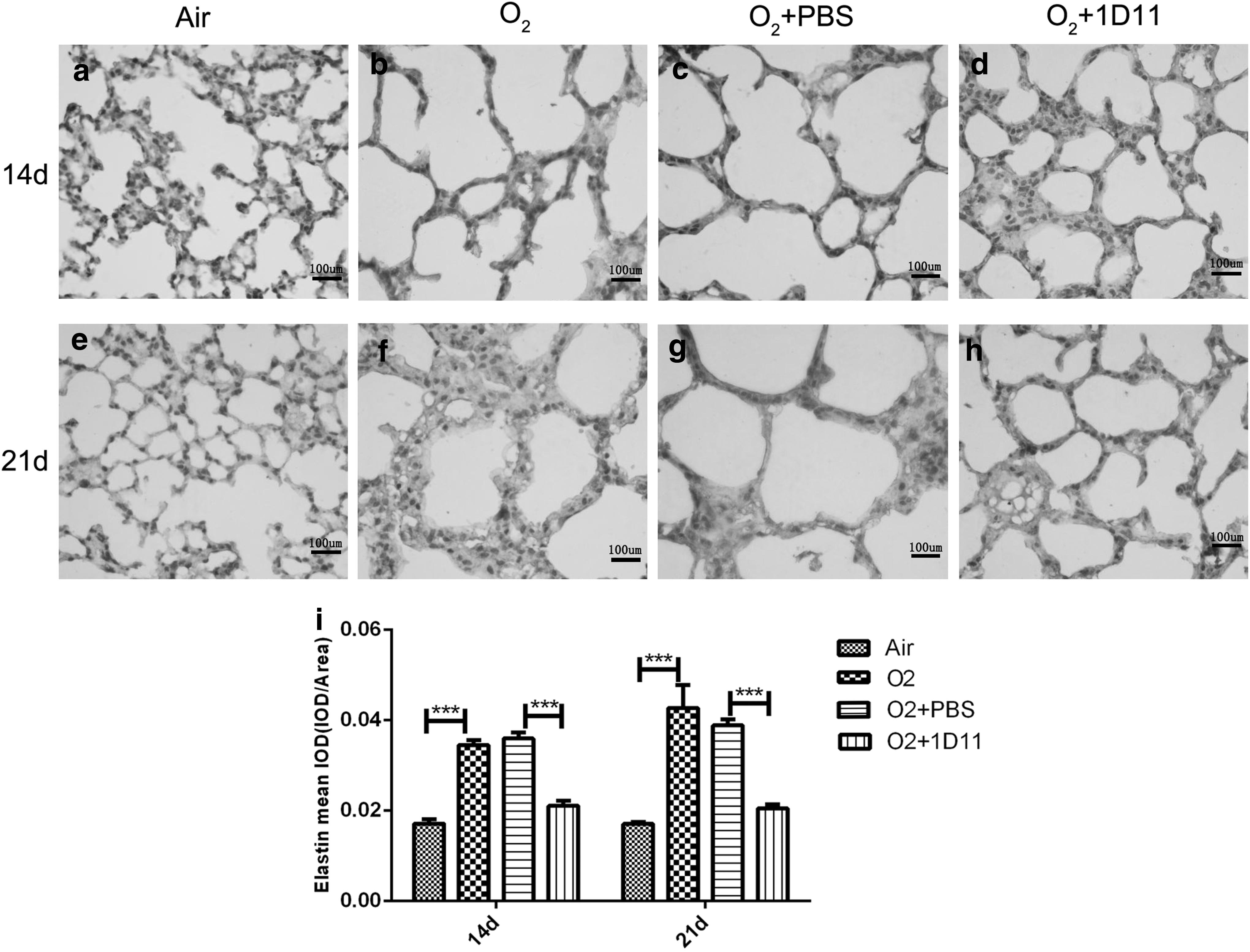
Disordered expression and distribution of elastin was improved by 1D11 treatment in the lung after 85% O2 exposure. Immunohistochemistry was used to investigate the position and amount of elastin (n = 6 animals, 12 fields/animal). One representative figure per group is shown. Bar = 100 μm. Positive staining of the tips of the alveolar septa is observed in the air control newborn lung. However, elastin distribution was spread over the airway wall in the lung obtained from 14 days
The expression of elastogenesis-related genes and proteins in the mice exposed to hyperoxia were decreased by 1D11 treatment
Elastogenesis-related proteins include tropoelastin, fibulin-5, and NE, which contribute to the synthesis, assembly, and degradation of elastin (Yanagisawa and others 2009). To evaluate whether 1D11 improved lung development and elastin distribution by regulating the expression of elastogenesis-related proteins, we measured the mRNA and protein expression of tropoelastin, fibulin-5, and NE in 14- and 21-day newborn O2-exposed lungs treated with 1D11. Tropoelastin is secreted from lung myofibroblasts and is then converted to elastin (Bax and others 2009).
As shown in Fig. 6, the expression of the tropoelastin gene was increased in the mouse lung when exposed to hyperoxia (Fig. 6a). After treatment with 1D11, the expression of tropoelastin was significantly decreased (Fig. 6a). Consistent with the mRNA expression, the protein expression of tropoelastin was increased in the O2 group (Fig. 7b). Its expression in the 1D11-treated mouse lung was significantly decreased (Fig. 7b). Fibulin-5 can help in the assembly of elastic fibers by binding to elastin (Hu and others 2006). The overexpression of fibulin-5 can inhibit the normal formation of elastic fibers (Albig and Schiemann 2005). In comparison with the air control group, the mRNA expression of fibulin-5 was significantly increased in the O2 group (Fig. 6b). Administration of 1D11 to the hyperoxia-exposed mouse lung resulted in a significant decrease in the expression of fibulin-5 (Fig. 6b). Consistent with the previous results of gene expression, the protein level of fibulin-5 was increased in the mouse lungs exposed to hyperoxia (Fig. 7c). After 1D11 treatment, the level of fibulin-5 protein was decreased (Fig. 7c). NE is a hydrolase that can degrade the extracellular matrix and can provide feedback to induce the accumulation of elastin (Takemasa and others 2012).
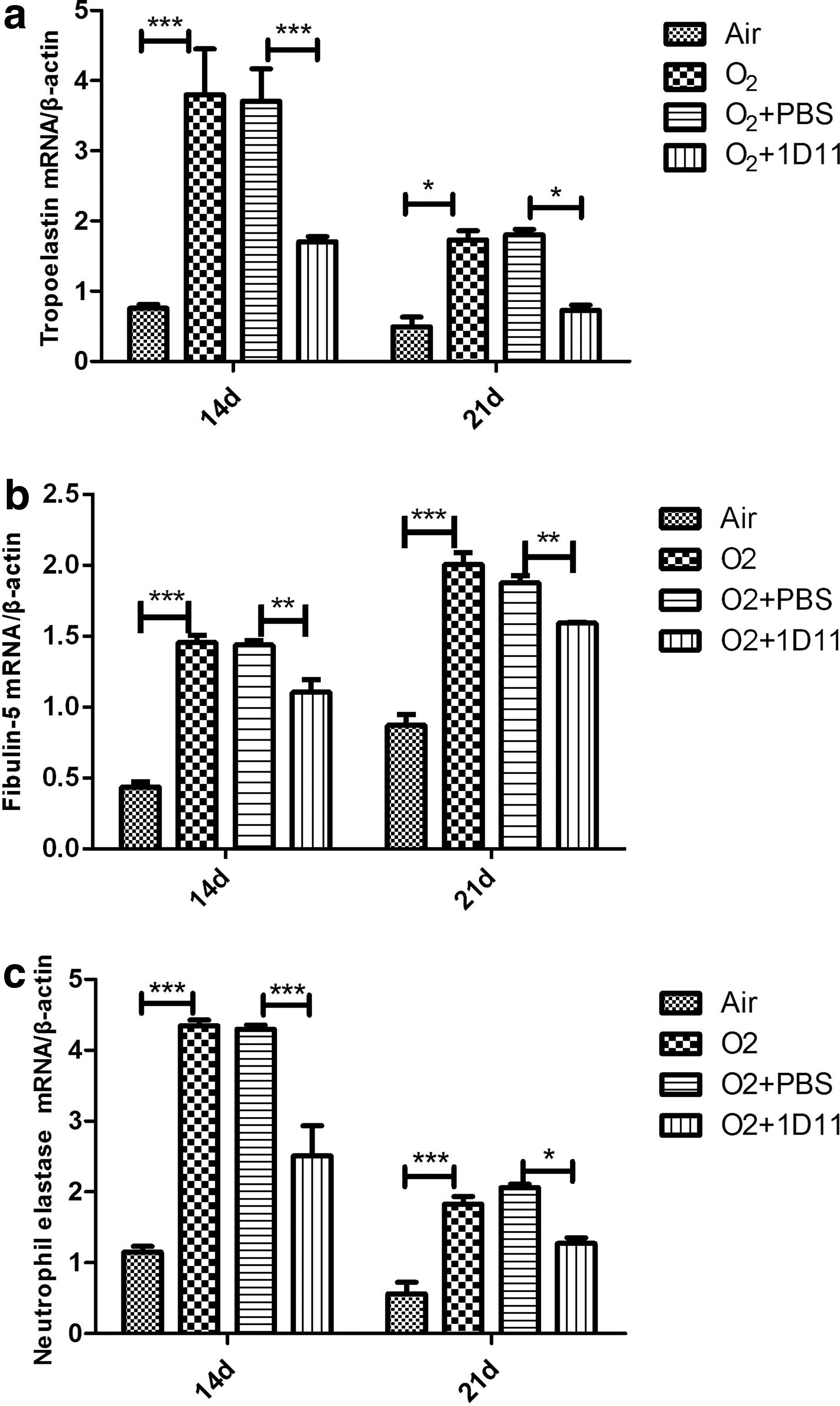
1D11 prevents the overexpression of genes related to the synthesis, assembly, and degradation of elastin after 14 and 21 days in mouse lungs after exposure to 85% O2. These genes are tropoelastin
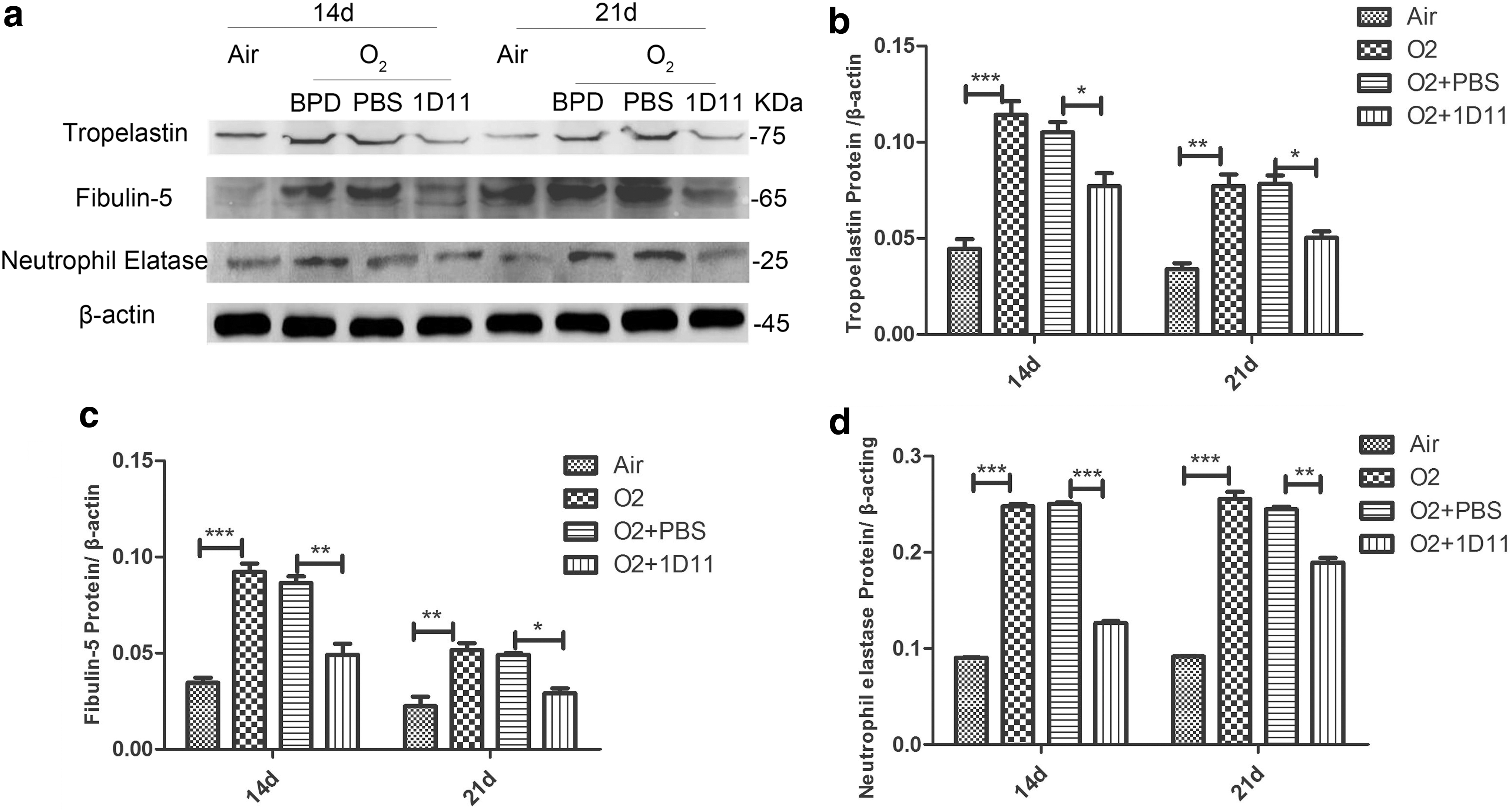
1D11 decreases the expression of elastogenesis-related proteins in the impaired newborn lung. Western blot detected the expression of tropoelastin, fibulin-5, and NE in total protein extract from the lung tissue. A representative example from a minimum of three independent experiments is shown
As shown in Fig. 6c, the gene expression of NE in the newborn lung after hyperoxia exposure exhibited a significant increase compared with air exposure (Fig. 6c). 1D11 treatment reduced the mRNA expression of NE compared with PBS treatment in lungs with hyperoxia exposure (Fig. 6c). Consistent with the mRNA data, the expression of NE protein was increased in the hyperoxia group lungs (Fig. 7d). A decreased protein level of NE was found in the O2-exposed mouse lungs that were treated with 1D11 (Fig. 7d). Based on the earlier data, we believe that hyperoxia may disturb the normal expression of elastogenesis-related proteins, causing accumulation of elastin. 1D11 may prevent this process, improving alveolar development. We speculate that proper blockade of the TGF-β signaling pathway may promote alveolarization by accelerating the harmonious expression of elastogenesis-related proteins.
Discussion
In our research, we observed that TGF-β signaling is activated and that alveolar structures are abnormal in the O2-exposed newborn mouse lung. 1D11 improved the development of pulmonary alveoli and the distribution of elastin in the newborn mouse lung exposed to O2 by blocking TGF-β signaling and by inhibiting the expression of its downstream target genes and proteins. The high expression level of elastogenesis-related proteins was inhibited by 1D11 in the hyperoxia-exposed mouse lung. These observations suggest that TGF-β activity participates in the upregulation of elastogenesis-related protein, which may contribute to serious disruption of elastin fiber and alveolar simplification. The TGF-β neutralizing antibody 1D11 may become a novel therapeutic approach to BPD.
TGF-β signaling activity is essential in lung development (Bartram and Speer 2004; Morty 2011; Wang and others 2015). Smad3 deficiency causes abnormal mouse lung alveolarization and centrilobular emphysema (Chen and others 2005; Jablonska and others 2010). Nevertheless, our previous study demonstrated that excessive TGF-β activity participated in the processes that result in abnormal expression and distribution of elastin (Han and others 2015). Inhibition of TGF-β/Smad signaling protects the injured mouse lung caused by hyperoxia through upregulation of the expression of peroxisome proliferator-activated receptor-γ (Kumarasamy and others 2009; Gong and others 2011).
In our research, activation of the TGF-β pathway and alveolar simplification were observed in the mouse lung exposed to hyperoxia. After treatment with 1D11, the activity of TGF-β signaling was significantly reduced, and alveolarization was improved. 1D11 attenuated vascular remodeling and protected alveolar development in the newborn lung by inhibiting TGF-β signaling and regulating the abnormal expression of ET-1 (Olave and others 2012) and plod2 (Witsch and others 2014a). Nakanishi found that 1D11 could attenuate TGF-β signaling and could improve extracellular matrix assembly, microvascular development, and alveolarization in the injured developing lung (Nakanishi and others 2007; Witsch and others 2014b).
Our research focused on the role of 1D11 in improving the process of the abnormal formation of elastin fibers. Elastin is the predominant component of elastic fiber, accounting for 90% of the total mass (Yanagisawa and others 2009). Excessive and disordered deposition of elastin is a prominent pathological feature in BPD (Bland and others 2007). Elastin is a basic material in lung development and plays an important role in normal alveolar septation (Kuang and others 2003). Deletion of the elastin gene in mice results in abnormal lung development and congenital emphysema (Shifren and others 2007). Some research has suggested that treatment with exogenous TGF-β induced rapid and transient increases in the levels of elastin heterogeneous nuclear RNA, followed by increases in elastin mRNA and protein expression (Morty 2011).
In our study, excessive elastin was distributed throughout the walls of the distal air spaces, and increased TGF-β was observed in the mice exposed to hyperoxia. After TGF-β signaling was blocked by 1D11 in the hyperoxia-exposed mouse lung, the expression of abnormally located elastin was decreased, and the elastin that was localized in the tips of terminal airspaces was increased. Thy-1-/-mice given 1D11 had improved alveolar development and decreased collagen and elastin (Shifren and others 2007). Study of the protective mechanism of 1D11 during the process of elastogenesis may help to further understand the pathogenesis of BPD and to find better targets for therapy.
Elastin fiber is very important in the integrity and distensibility of the airway structure (Bland and others 2008). The formation of elastin fibers involves a large number of proteins (Han and others 2015). Discordant expression of elastogenesis-related proteins may lead to disordered formation of elastin fibers (Shifren and others 2007). Tropoelastin is secreted from lung myofibroblasts and is a soluble monomer of elastin (Bland and others 2008). Our study found that hyperoxia promoted the expression of tropoelastin and activated TGF-β. Previous research found a significant upregulation of smooth muscle cell tropoelastin production induced by TGF-β (Takacs and others 2012). This result is in agreement with our findings. The increased expression of tropoelastin can promote elastin synthesis (Choi and others 2010). The C-terminal domain of tropoelastin plays an important role in the assembly of elastin (Akhtar and others 2011). Hyperoxia damages the assembly of elastic fibers by impairing the C-terminal domain of tropoelastin, and it decreases the molecular interaction between tropoelastin and fibulin-5 (Akhta and others 2010).
Fibulin-5 is an extracellular matrix protein and is a critical determinant in elastic fiber organization (Mokres and others 2010). Fibulin-5 contributes to the process of elastogenesis by binding to structural components, including tropoelastin and fibrillin-1, helping in elastic fiber assembly (Kozel and others 2004). Knockout mice (fibulin-5-/-) have a tortuous aorta with loss of compliance, severe emphysema, and loose skin (Nakamura and others 2002).
We found that fibulin-5 was increased in the mouse lung exposed to hyperoxia. Overexpression of fibulin-5 led to elevated expression and accumulation of elastin as well as the production of desmosine, indicating that fibulin-5 facilitates cross-linking of tropoelastin and destroys the formation of normal elastic fibers (Nonaka and others 2009). TGF-β stimulates murine fibroblasts to synthesize fibulin-5 transcripts and protein (Bartram and others 2004). This finding supports our results, showing that 1D11 inhibited the expression of fibulin-5 by blocking the TGF-β signaling pathway.
NE is a proteinase that can hydrolyze components of the extracellular matrix such as elastin (Lee and others 2006). The degradation of elastin is very important in the process of elastogenesis (Akhta and others 2010). Hydrolyzation of elastin can lead to activation of the TGF-β signaling pathway, which causes increased myofibroblasts (Takemasa and others 2012). NE enhances TGF-β gene expression and protein production in human airway smooth muscle cells (Lee and others 2006). The NE inhibitor sivelestat attenuates subsequent ventilator-induced lung injury in mice by suppressing apoptotic responses (Sakashita and others 2007). Our previous study showed that the elastase inhibitor elafin could ameliorate alveolarization by preventing neutrophil influx and by the activation of TGF-β (Han and others 2017).
In our research, the expression level of NE was inhibited by 1D11, indicating that NE not only influenced TGF-β activation but also was driven by TGF-β signaling. We speculate that hyperoxia may stimulate the expression of NE and that it may promote the proliferation of myofibroblasts that secrete tropoelastin and activate TGF-β signaling. Activation of TGF-β signaling upregulates the expression of tropoelastin, fibulin-5, and NE. Then, tropoelastin is increased, whereas hyperoxia impairs the C-terminal domain of tropoelastin. Increased fibulin-5 connects the abnormal tropoelastins, and this process may cause disturbed assembly of elastic fibers and may lead to disordered accumulation of elastin. 1D11 blocked TGF-β signaling and decreased the overexpression of these genes and proteins and, subsequently, the disordered accumulation of elastin; thus, the structure of elastin fibers was improved.
In summary, our research demonstrated that 1D11 has a protective effect on the newborn mouse lung exposed to hyperoxia. The possible mechanisms are that 1D11 alleviated the disordered abundance and distribution of elastin in distal airways by blocking TGF-β signaling and by downregulating elastogenesis-related proteins. However, 1D11 could not complete the healing of the abnormal alveolarization of BPD, suggesting that other signaling pathways may participate in the pathogenesis of BPD and may need to be explored in future studies.
Footnotes
Acknowledgments
We thank Prof. Xianqing Jin for providing technical assistance and insightful discussions during the preparation of the article. We thank Dr. Xiaoyong Zhang of the Wistar Institute (USA), who provided medical writing services. This research was supported by the National Natural Science Foundation of China (Grant Nos. 81270058 and 30770950), the Chongqing Natural Science Foundation (CSTC; Grant No. 2009BB6072), and the Science and Technology Research Project of Chongqing Education Committee (Grant No. KJ1500229).
Authors' Contributions
S.D., C.G., and W.H. designed and performed the experiments, analyzed the data, and prepared the article. Chun Deng bred the mice. Chun Deng and Han Zhang analyzed the data, and evaluated and wrote the article.
Author Disclosure Statement
The authors declare that they have no competing interests.