
Abstract
Long noncoding RNAs (lncRNAs) exhibit highly lineage-specific expression and act through diverse mechanisms to exert control over a wide range of cellular processes. lncRNAs can function as potent modulators of innate immune responses through control of transcriptional and posttranscriptional regulation of mRNA expression and processing. Recent studies have demonstrated that lncRNAs participate in the regulation of antiviral responses and autoimmune disease. Despite their emerging role as immune mediators, the mechanisms that govern lncRNA expression and function have only begun to be characterized. In this study, we explore the role of lncRNAs in human plasmacytoid dendritic cells (pDCs), which are critical sentinel sensors of viral infection and contribute to the development of autoimmune disease. Using genome-wide sequencing approaches, we dissect the contributions of Toll-like receptor 7 (TLR7) and type I interferon (IFN-I) in the regulation of coding and noncoding RNA expression in CAL-1 pDCs treated with R848 or IFNβ. Functional enrichment analysis reveals both the unique and synergistic roles of TLR7 and IFN-I signaling in the orchestration of pDC function. These observations were consistent with primary cell immune responses elicited by detection of viral infection. We identified and characterized the conditional TLR7- and IFN-I-dependent regulation of 588 lncRNAs. Dysregulation of these lncRNAs could significantly alter pDC maturation, IFN-I and inflammatory cytokine production, antigen presentation, costimulation or tolerance cues, turnover, or localization, all consequential events during viral infection or IFN-I-driven autoimmune diseases such as systemic lupus erythematosus. These findings demonstrate the differential responsiveness of lncRNAs to unique immune stimuli, uncover regulatory mechanisms of lncRNA expression, and reveal a novel and tractable platform for the study of lncRNA expression and function.
Introduction
D
Plasmacytoid dendritic cells (pDCs) have been extensively implicated in the pathogenesis of SLE (Rowland and others 2014; Sisirak and others 2014) as a major source of IFN-I, generating up to 1,000 times more than any other cell type (Cella and others 1999; Siegal and others 1999). Acting as phagocytic surveillance sentinels, pDCs are specialized in the detection of nucleic acids and production of IFN-I through high constitutive expression of the endosomal PRRs, TLR7 and TLR9, which engage single-stranded RNA or unmethylated CpG DNA, respectively (Reizis and others 2011). IFN-I produced by pDC is key in initiating protective resistance to infection in peripheral tissues and modulating the activity of other immune cells, as studies in murine models of pDC depletion have shown (Swiecki and Colonna 2010). As such, pDCs bridge the intersection of innate and adaptive immunity, and therefore are strategic targets for the manipulation of autoimmune responses (Swiecki and Colonna 2015). IFN-I is also essential to the function of pDCs, as autocrine/paracrine IFN-I signaling in activated pDCs results in amplification of subsequent IFN-I, inflammatory cytokine production, and modulation of pDC effector functions such as expression of costimulatory molecules and chemokine receptors (Asselin-Paturel and others 2005; Gautier and others 2005; Prakash and others 2005; Siren and others 2005; Liao and others 2010; Lorenzi and others 2011; Oh and others 2011; Bao and Liu 2013; Kim and others 2014; Pantel and others 2014).
Long noncoding RNAs (lncRNAs) (noncoding RNAs >200 nucleotides in length) have emerged as potent regulators of varied cellular processes through diverse mechanisms (Cech and Steitz 2014). lncRNAs are derived from pseudogenes, alternatively transcribed or spliced coding genes, antisense transcription, introns, or standalone intergenic regions (Griffiths-Jones 2007; ENCODE Project Consortium 2012; Derrien and others 2012; Djebali and others 2012). Many lncRNAs share features of coding genes such as promoter-controlled expression, exon splicing, and polyadenylation (Cabili and others 2011; Hung and others 2011). The functional activities of lncRNAs are largely linked to their subcellular localization. In the nucleus, lncRNAs direct epigenetic and transcriptional regulation (Khalil and others 2009; Bonasio and Shiekhattar 2014). In the cytosol, lncRNAs are key participants in posttranscriptional mRNA regulation (Yoon and others 2013), where lncRNA sequence complementarity, conserved RNA binding protein (RBP) motifs, and labile secondary and tertiary structures can influence their selective dynamic associations (Cook and others 2015). In the absence of a requirement to maintain codon integrity, lncRNAs display a high degree of evolutionary permissiveness to mutation, resulting in highly species-specific signatures, with <10% of lncRNA loci conserved between human and mouse (Johnsson and others 2014). While sequence conservation among lncRNA loci across species is low, an increasing appreciation for conservation in lncRNA structural elements is emerging (Nitsche and Stadler 2017). Mutations that disrupt sequence specificity and RBP motifs, or perturb secondary/tertiary structure may alter lncRNA function, and thereby dysregulate processes that define disease outcomes. Indeed, genome-wide association studies demonstrate that the majority of disease-associated variants fall outside protein-coding regions (Ricano-Ponce and Wijmenga 2013; Welter and others 2014), suggesting the functional importance of lncRNAs at the host–pathogen interface and in disease pathogenesis (Kapusta and Feschotte 2014).
lncRNAs are highly lineage specific (Kim and others 2015; Gloss and Dinger 2016), even more so than coding or small noncoding RNA signatures. Such features position them as critical regulators of cell subset differentiation and cell type-specific activities; however, the exact mechanisms through which lncRNAs modulate pDC development and function are unknown. While gene expression profiling in human pDCs has defined cell type-specific coding genes (Waddell and others 2010; Miller and others 2012), a comprehensive description of the long noncoding transcriptional landscape in pDCs has been challenging due to limited numbers of cells and low levels of lncRNA expression. To gain a better understanding of the noncoding transcriptional landscape in activated pDCs, we undertook comprehensive transcriptome profiling in a tractable human pDC line following TLR7 engagement or IFN-I signaling. We identified novel subsets of lncRNA species that are differentially regulated by these innate immune stimuli, neighboring genes that are co-regulated with lncRNAs, and transcriptional regulators likely to coordinate lncRNA expression. We compared these transcriptome profiles with those of activated primary pDCs, demonstrating that these data provide a solid platform for the study of lncRNA expression and their roles in pDC development and effector functions. Further characterization of lncRNAs described in this study will identify new biomarkers and therapeutic targets for disease.
Materials and Methods
Cell culture and reagents
CAL-1 pDCs (a kind gift from Takahiro Maeda) were cultured as previously reported (Steinhagen and others 2013). Cells were rested for 6 h in 0.1% fetal bovine serum (FBS) CAL-1 media before stimulations with 1,000 U/mL recombinant human IFN-β (PBL Interferon Source), 1 μg/mL R848 (InvivoGen) with or without 1 μg/mL recombinant vaccinia virus B18R protein (Thermo Fisher Scientific), or CpG-B (InvivoGen). Stimulated cells were incubated at 37°C for the indicated times; 20 × 106 frozen MUTZ3 progenitor cells were resuspended in 40 mL 20% minimum essential media (MEM)-alpha containing ribonucleosides/deoxyribonucleosides, 20% FBS, and 10% conditioned medium from bladder carcinoma cell line 5638 and cultured at 37°C for 4 days. Differentiation to immature conventional dendritic cell (cDC) was induced by culture at 0.25 × 106/mL MEM-alpha 20% FBS without 10% bladder line media, with the addition of 100 ng/mL granulocyte-macrophage colony-stimulating factor (GM-CSF) (PeproTech), 10 ng/mL interleukin (IL)-4 (Shenandoah Biotechnology), and 2.5 ng/mL tumor necrosis factor-α for 7 days, replacing half medium after 3 days. Cells were then stimulated with 100 ng/mL lipopolysaccharide (LPS; Invivogen) for 12 h. THP-1 monocyte-derived dendritic cells (moDC) were derived and stimulated as previously described (Lim and others 2016).
Primary cell analyses
Primary monocytes and monocyte-derived DCs were obtained, cultured, and stimulated with LPS or Pam2CSK4 as previously described (Lim and others 2016).
Subcellular fractionation
CAL-1 cells were cultured and stimulated as described above for 0, 3, and 12 h. RNA from nuclear and cytoplasmic cellular fractions was isolated from cell lysates using the Ambion PARIS™ system (Thermo Fisher Scientific) as per the manufacturer's instructions.
siRNA knockdowns
CAL-1 pDC were nucleofected with 1 μM siRNAs at 1.2 × 106 cells/well in supplemented SF media from the SF Cell Line 4D-Nucleofector X Kit (Lonza) using an Amaxa nucleofector with 96-well plate strips (Lonza) on program DN-100 as per the manufacturer's instructions. siRNAs were obtained from Integrated DNA Technologies (IDT) targeting lncRNAs using the following sequences: lnc-DC 5′-GAGTTATCTTAAGGATCAT-3′, lnc-1133 5′-CUUGCAGGAAGGAUGGAUUCUCC (CA)-3′, and lnc-ROR 5′-GGAGAGGAAGCCTGAGAGT-3′. Proprietary triplicate siRNA mixes were obtained from Dharmacon targeting lncRNAs: lnc-515 SMARTpool: Lincode LINC00515 siRNA R-189581-00-0005 and lnc-SIPA1L1-2 SMARTpool: Lincode Loc145474 siRNA R189240-00-0005. Nucleofected cells were rested and stimulated with R848 as described above for the indicated times, with RNA collection at 18 h postnucleofection.
RNA isolation, reverse transcription, and quantification of gene expression
Total RNA was extracted by the NucleoSpin RNA extraction kit (Macherey-Nagel) and cDNA was reverse transcribed with the QuantiTect RT kit (Qiagen) according to the manufacturer's instructions. TaqMan qPCR was conducted using the ViiA7 qPCR system (Life Technologies) with the following probe assays obtained from IDT: lnc-DC Hs.PT.58.4925540, lnc-SIP1AL1-2 Hs.PT.58.24273269.g, lnc-ROR Hs.PT.58.40897944, lnc-515 Hs.PT.56a.24878664.g, lnc-578 Hs.PT.58.23294839, and lnc-1133 Hs.PT.58.40059900. Gene expression levels were normalized to HPRT or GAPDH as indicated. Data were analyzed using the QuantStudio real-time PCR software (Applied Biosystems).
RNA sequencing and bioinformatic analysis
CAL-1 cells (1 million cells/mL) were cultured in 0.1% FBS CAL-1 media in triplicate in a 12-well tissue culture plate for 6 h before stimulation with 1,000 U/mL recombinant human IFN-β (PBL Interferon Source) or 1 μg/mL R848 (Invitrogen), or 1 μg/mL R848 (Invitrogen) +1 μg/mL recombinant vaccinia virus B18R protein (Thermo Fisher Scientific) or mock stimulation. Stimulated cells were incubated for 12 h at 37°C and RNA was isolated as described above. RNA integrity was determined using the RNA 6000 Nano Kit with a 2100 bioanalyzer (Agilent Genomics) and quantified using the fluorometric Qubit™ RNA BR assay kit (Invitrogen). cDNA libraries were prepared with the TruSeq Stranded mRNA Library Prep Kit and sequenced on an Ilumina NextSeq 500 sequencer. Library preparation, QC, and sequencing were carried out by Seattle Genomics (
Both the genome sequence (fasta) and gene transfer files (gtf) for human were obtained using igenomes (
Transcript annotation
Annotation of lncRNAs was done by filtering transcripts through “transcript biotype” designations and universal nomenclatures were derived from LNCipedia database (Volders and others 2015). Chromosome location, start and end positions, and strand orientation of the lncRNAs were retrieved using “biomaRt” (version 2.34.2) in R. Circos plots were generated using the RCircos (version 1.2.0) package in R.
Nearest neighbor analysis
Nearest neighbor genes for DE lncRNAs were identified with the seq2pathway package in R statistical programming language using RStudio. Functional analysis of nearest neighbor coding genes by gene ontology term enrichment was performed using Enrichr (Chen and others 2013).
Transcription factor enrichment analysis
Transcription factor (TF) enrichment was performed using the ENCODE Chip-seq dataset
Statistical analysis
GraphPad Prism 7.0 (GraphPad Software, Inc.) was used for statistical analysis and plotting of reverse transcription–polymerase chain reaction (RT-PCR) data. Significance in gene expression differences is plotted as mean ± standard error of the mean and was determined using 2-tailed unpaired Student's t-test for stimulations versus mock or by one-way analysis of variance across multiple stimulations (P < 0.05, indicated by asterisk *). Pearson correlation analyses and plots were performed using the ggplot package in R statistical programming language using RStudio.
Data dissemination
The data generated in this study are available through the following accession identifiers on the NCBI GEO database (GSE117127). RNA sequencing data from primary monocytes treated with IFN-α for 6 h were retrieved from the NCBI-GEO database under accession number GSE72502 (Hung and others 2015). RNA sequencing data from influenza-infected primary pDC1/2/3 subset were retrieved from NCBI-GEO database under accession number GSE84204 (Alculumbre and others 2018).
Results
TLR7 and IFN-I signals drive broad transcriptional change in human pDCs
We utilized human CAL-1 cells, which phenotypically and functionally resemble primary peripheral pDCs (Maeda and others 2005; Steinhagen and others 2013), to understand the differences in the response to TLR7 and/or IFNAR ligation. CAL-1 cells were treated for 12 h with the TLR7 agonist R848 in the presence or absence of the pan-IFN-I inhibitory protein B18R (Colamonici and others 1995; Symons and others 1995), or treated with IFN-β, enabling dissection of the relative contributions of TLR and IFN-I signaling to gene expression (Fig. 1A). Treatment with R848 led to a significant induction of IFNB1 expression, which was partially decreased upon blockade of the IFNAR-mediated TLR amplification loop (R848 + B18R) (Fig. 1B left and Supplementary Fig. S1A; Supplementary Data are available online at

Transcriptome sequencing in IFN-I- and TLR7-stimulated human pDCs.
Functional analysis of coding DEGs across these stimulatory conditions revealed enrichment of gene sets pertaining to IFN-I signaling, PRR activation, and dendritic cell (DC) functions (Fig. 1G). Accordingly, we observed enrichment of IFN signaling modules and strong induction of classical ISGs (Supplementary Table S1) by TLR7 and IFN-I stimulations, which were markedly diminished by IFN-I blockade during TLR7 activation. In contrast, enrichment of DEGs related to DC functions such as antigen presentation and lymphocyte engagement was modulated primarily by TLR engagement versus IFN-I (Fig. 1G). However, control of DEG expression regulated by interferon regulatory factors (IRFs) was similar downstream of both IFN-I and TLR stimulations, indicating redundancy in the pathways that control the activity of these factors (Fig. 1G). To better understand the complexity of TLR7-mediated responses, we implemented a regulatory network analysis, which demonstrated robust production of IFN-I (IFN-α and IFN-β), the activation of the signal transducer, and activator of transcription (STAT; STAT1; and STAT3), IRFs, NF-κB (RELA, NFKB1, and NFKBIA), and activator protein 1 (AP-1) (Fig. 1H, left). The strength of these activities was diminished in the absence of IFN-I signaling following TLR7 activation, but the diversity of mediators is preserved (Fig. 1H, middle). In contrast, responses to IFN-I were predicted to be primarily driven by STATs and IRFs, and there was a notable loss of NF-κB and AP1 activity (Fig. 1H, right). These data demonstrate that the stimulus-specific transcriptional response to TLR7 agonists and IFN-I is driven by unique TFs. In addition, the induction of IFN-I further promotes cellular activation, and cytokine production and maturation. Given the consistency of the observed gene expression patterns in CAL-1 cells with those observed in primary human and murine pDCs (Merad and others 2013; Heidkamp and others 2016), this cell line provides a viable platform for the investigation of immune-responsive noncoding RNAs.
TLR7 and IFN-I stimulations induce distinct lncRNA signatures
Further characterization of the DEGs modulated in these conditions revealed that roughly one-fourth are noncoding transcripts (Fig. 2A and Supplementary Fig. S2A). Notably, in contrast to nearly equivalent numbers of upregulated and downregulated DEGs among coding loci, noncoding loci display a marked trend toward more genes downregulated than upregulated following TLR7 activation, and IFN-I blockade enhances this phenomenon (Fig. 2A). Among the noncoding transcripts, the majority (n = 588) were identified as lncRNAs, with their distribution across lncRNA biotypes biased toward antisense transcripts (33%), standalone intergenic lincRNAs (30%), and pseudogenes (29%) (Fig. 2B). We did not capture a bias among lncRNA biotypes in the directionality of expression (Fig. 2C and Supplementary Fig. S2B).
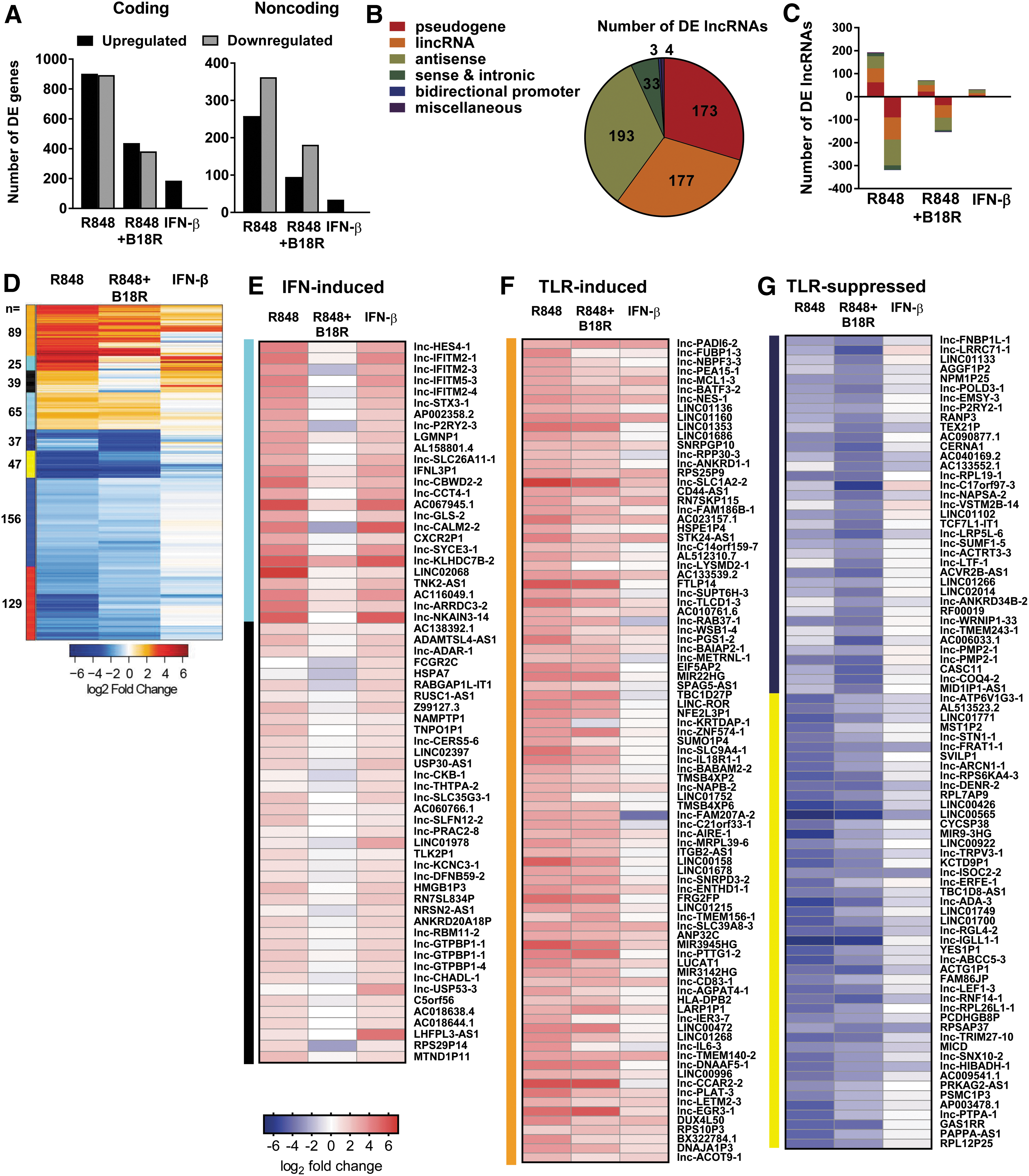
TLR- and IFN-I-modulated lncRNAs in human pDCs.
Unsupervised clustering of lncRNA relative expression yielded unique response modules, identifying lncRNAs under the regulation of either TLR7 or IFN-I (Fig. 2D and Supplementary Table S2). Consistent with the induction of coding ISGs by IFN-I, we identified 64 lncRNAs induced by exogenous IFN-β and whose induction is specifically dependent upon auto/paracrine IFN-I signaling in the context of TLR7 activation, as evidenced by the impact of IFN-I blockade (Fig. 2E). Conversely, we identified 89 lncRNAs which are generally indifferent to IFN-I signaling but are induced strictly by TLR engagement through mechanisms independent of IFN-I feedback (Fig. 2F). We identified a large set of lncRNAs (n = 369) whose expression is inhibited following TLR engagement (Fig. 2D and Supplementary Table S3). Of these downregulated lncRNAs, 84 were markedly downregulated by R848 treatment despite IFN-I exposure (Fig. 2G). In sum, these 588 DE lncRNAs comprise a novel definition of lncRNA sensitivity to TLR and IFN-I stimuli.
Immune-regulatory pathways govern TLR- and IFN-I-modulated lncRNAs and co-expressed neighboring coding genes
Global analyses of lncRNA and neighboring coding gene expression patterns have revealed modest associations in expression levels (Kutter and others 2012). Patterns of co-expression in IFN-I-induced lncRNAs and nearby coding genes have been identified in murine (Josset and others 2014) and human (Barriocanal and others 2014; Carnero and others 2014; Kambara and others 2014a, 2014b) epithelia and demonstrated for several lncRNA-neighboring ISG pairs (Carnero and others 2014). To better understand the distinct co-regulation of lncRNAs and neighboring genes by TLR7 and IFN-I signaling cascades in human pDCs, we undertook a global co-expression analysis. First, we identified broad genomic distribution across the DE coding genes and lncRNAs (Fig. 3A). Then, we identified the nearest coding neighbor across the previously identified TLR7- and IFN-I-regulated modules. Noticeable concordance in the directionality of gene expression was noted between the IFN-I-induced lncRNAs and their neighboring genes (Fig. 3B). Similar co-expression was observed among the TLR-induced (Fig. 3C) and TLR-suppressed (Fig. 3D) lncRNAs and their neighboring genes, with some examples of inverse directionality, suggesting it is likely that lncRNA expression is transcriptionally regulated by similar factors regulating coding gene expression. To explore the signaling and transcriptional influences responsible for lncRNA-neighbor gene co-expression, we performed functional analysis of the nearest neighbor coding genes. Functional enrichment analysis demonstrated that the coding genes neighboring IFN-induced lncRNAs are involved in the response to IFN-I and STAT activation (IL-35, IL-23, and IL-21 signaling pathways) (Fig. 4A; top). TLR-inducible lncRNAs were in close proximity with not only ISGs but also genes involved in antigen presentation (Fig. 4A; middle). Interestingly, TLR-suppressed lncRNAs were found near genes involved in hormonal regulation and membrane integrity pathways (Fig. 4A; bottom). Given the parallels in functional enrichment between these nearest neighbor coding genes (Fig. 4A) and those observed from the global signatures (Fig. 1G), these IFN-responsive and TLR-responsive lncRNAs are likely under the regulation of the previously identified master regulators of TLR7- and IFNAR-mediated signaling (Fig. 1H).

TLR-/IFN-modulated lncRNAs are co-expressed with neighboring coding genes.

Immune modulatory control of lncRNA expression changes.
We leveraged the ENCODE ChIP-seq database to identify TF binding sites upstream of the transcriptional start site of the IFN- and TLR-responsive lncRNA loci. We found an appreciable enrichment of STAT, IRF, NF-κB, and AP-1 binding at sites proximal to a subset of lncRNAs (Fig. 4B). Overall, IFN-induced lncRNAs display enhanced representation of IRF binding sites and AP-1 binding sites (Fig. 4C), while TLR-induced lncRNAs display enhanced representation of NF-κB and AP-1 binding sites (Fig. 4D). In contrast, TLR-suppressed lncRNAs displayed less enrichment of binding sites for these TFs (Fig. 4E), suggesting that TLR activation likely induces additional negative regulators of gene expression. Collectively, these analyses demonstrate the role of TLR- and IFN-I-dependent transcriptional mediators in control of lncRNAs and their neighboring coding gene expression patterns in pDCs, and specifically implicate NF-κB and IRF proteins in their respective transcriptional control. These data suggest that many lncRNAs and their neighboring genes are coordinately regulated by similar canonical TLR- and IFN-I-signaling pathway mediators.
IFN-I- and TLR-modulated lncRNAs have diverse cytoplasmic functions
The limited abundance of primary pDCs is prohibitive for investigations of lncRNA function. To test the validity of CAL-1 as a model for exploration of functionally relevant lncRNAs, we screened these transcriptome profiles in conjunction with reports of known lncRNA functions to identify 6 lncRNAs representative of each TLR/IFN modulation group for further study: lnc-DC (WFDC21P, LOC645638), lnc-SIPA1L1-2 (LOC145474), lnc-ROR (LINC-ROR), lnc-515 (LINC00515), lnc-578 (LINC00578), and lnc-1133 (LINC01133) (Table 1). Validation of expression changes by RT-PCR in IFN-β and R848 +/− B18R treated CAL-1 cells revealed lnc-DC and lnc-SIPA1L1-2 displaying IFN-I and TLR inducibility, lnc-ROR and lnc-515 induced only by TLR stimulation, and lnc-1133 and lnc-578 showing significant downregulation following TLR engagement (Fig. 5A). Analysis of transcript modulation kinetics across 18 h following stimulation revealed variable patterns in the timing of inducibility/suppressibility in response to these stimuli, with evident effects of waves of autocrine/paracrine cytokine signaling as well as the influence of translated ISGs in boosting lncRNA levels (Supplementary Fig. S3A). We also tested the response of these lncRNAs to TLR9 agonism with CpG DNA, resulting in expression changes comparable to those observed in TLR7 stimulation (Supplementary Fig. S3B). Modulation in response to TLR2 and TLR4 agonists in myeloid cDC lineages showed similar patterns of change in abundance of these lncRNAs to that of TLR7 stimulation in pDCs, with the exceptions in lnc-ROR and lnc-578 (Supplementary Fig. S3C–E). We found similar patterns of expression in primary human myeloid lineages following TLR or IFN-I stimulation (Supplementary Fig. S3F, G).
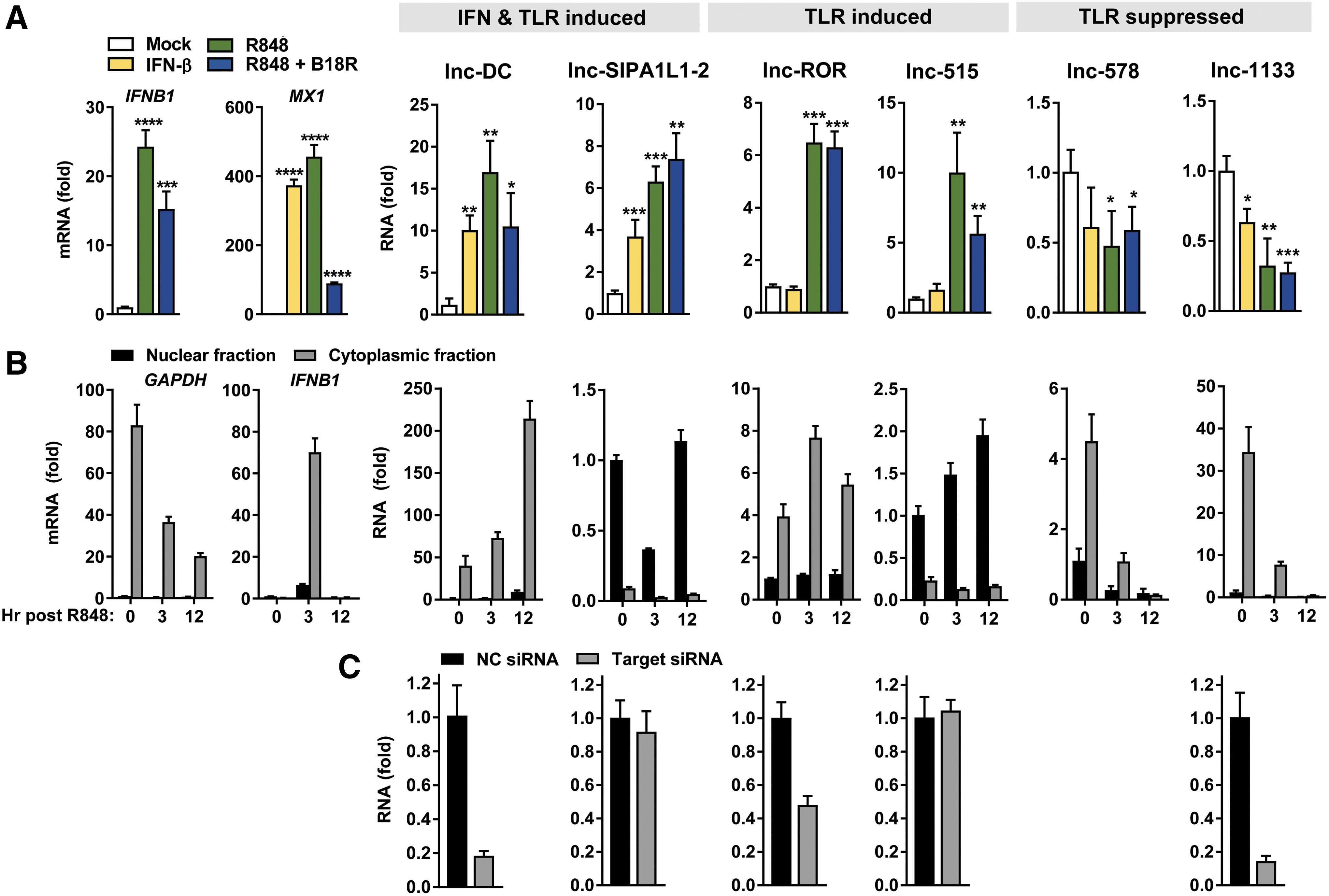
Analysis of select nuclear and cytoplasmic TLR-/IFN-modulated lncRNAs.
lncRNA, long noncoding RNA.
lncRNAs termed competing endogenous RNA (ceRNA) function as decoys, competing with mRNAs for occupation of RBPs and microRNAs (miRNAs), thereby relieving gene repression or stabilization by these factors (Tay and others 2014). These activities occur within the cytosol, where lncRNAs can also modulate mRNA decay by masking RBP or miRNA binding sites, structurally support mRNA stability, disrupt translational machinery, and control the activities of signaling mediators through occupation of protein interaction or modification sites (Jalali and others 2013; Forero and others 2017). Through their ability to regulate the repressive activities of miRNAs, RBPs, or other enzymes, cytoplasmic lncRNAs are critical determinants of posttranscriptional gene regulation and cellular signaling. Thus, we focused our analysis on lncRNAs within this cytoplasmic niche. We determined the subcellular localization of the 6 validated lncRNAs in CAL-1 pDC by fractionation of nuclear and cytoplasmic compartments, identifying 3 of 6 lncRNAs enriched within each compartment (Fig. 5B). Consistent with prior reports, lnc-DC (Wang and others 2014), lnc-ROR (Wang and others 2013), and lnc-1133 (Wu and others 2017) localize within the cytosol, a finding also evidenced by their sensitivity to RNAi (Fig. 5C). Our demonstration of the amenability of this culture system to RNAi manipulations and the consistency of lncRNA localization and modulation patterns in CAL-1 with reports of their behavior in other tissues and primary cells supports the validity of this model for subsequent lncRNA characterizations.
TLR7-mediated gene expression changes reflect those observed in primary pDCs
We have identified TLR7-responsive coding and noncoding RNAs, their regulators, and subcellular localization in in vitro pDC cultures. To evaluate whether these observations faithfully recapitulate the transcriptional changes captured in primary human pDCs, we compared CAL-1 gene signatures with those of ex vivo virus-infected pDCs (Alculumbre and others 2018). In vitro infection of pDCs with influenza virus is reported to activate TLR7 to induce IFN-I and proinflammatory cytokines (Cella and others 2000), and was shown to induce differentiation of ex vivo primary human pDCs into subsets that primarily produce IFN-I (“pDC1” which are PD-L1hi CD80lo), those with reduced IFN-I production, but elevated antigen presentation capacity (“pDC3,” which are PD-L1lo CD80hi), or an intermediate phenotype (“pDC2,” which are which are PD-L1hi CD80hi), as measured by flow cytometry (Alculumbre and others 2018). We compared the transcriptome of R848-stimulated CAL-1 with those of in vitro influenza-infected sort-purified primary pDC1, pDC2, and pDC3 cells. We observed a moderate correlation in coding DEG expression between TLR7-stimulated CAL-1 and virus-infected primary pDC subsets (Fig. 6A). More importantly, we found a high to moderate degree of correlation in the expression of DE lncRNAs between CAL-1 and pDC1, pDC2, and pDC3 subsets (Fig. 6B). Correlation of these DEG patterns between mRNAs and lncRNAs reflects concordance between TLR7 ligands and culture systems (Fig. 6C). Notably, we observe higher correlations between DE lncRNAs than mRNAs common to each dataset (Fig. 6C), consistent with the phenomenon of lncRNA signatures more tightly reflecting cellular lineage and activation states than coding gene signatures. Functional enrichment analysis of coding DEGs common to TLR7-stimulated CAL-1 and virus-infected primary pDCs revealed genes involved in inflammatory cytokine and IFN-I signaling, and antiviral responses (Fig. 6D). This analysis demonstrates that the CAL-1 cell line displays similarities with primary human pDCs in their responses to TLR7 agonists.
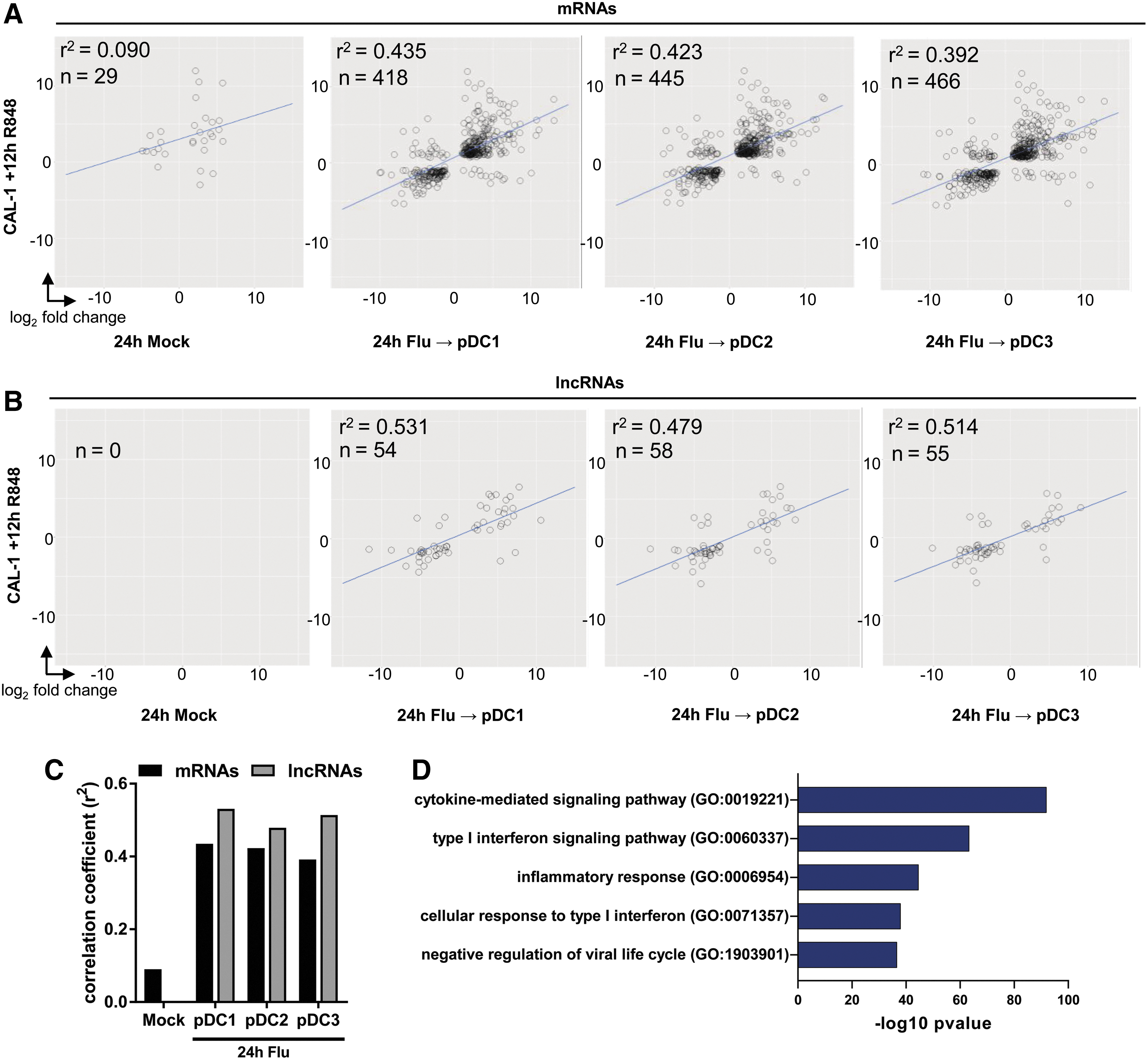
Transcriptomes of CAL-1 pDC correlate with in vitro infected primary pDCs.
Discussion
lncRNAs have been implicated in essential processes in immune responses such as control over cytokine and antiviral gene expression, regulation of signal-mediating TFs, and control of PRR activation (Willingham and others 2005; Carpenter and others 2013; Gomez and others 2013; Rapicavoli and others 2013; Bidet and others 2014; Cui and others 2014; Heward and Lindsay 2014; Li and others 2014; Ouyang and others 2014; Turner and others 2014; Wang and others 2014; Chan and others 2015; Liu and others 2015; Atianand and others 2016, 2017; Castellanos-Rubio and others 2016; Hu and others 2016; Tong and others 2016; Jiang and others 2018). Studies of gene expression changes following viral infection have profiled marked changes in lncRNA signatures (Peng and others 2010; Yin and others 2013; Zhang and others 2013; Josset and others 2014; Ouyang and others 2014, 2016; Winterling and others 2014). Other studies have examined the contributions of antiviral cytokines or PRR stimuli as upstream regulators of lncRNA expression (Carpenter and others 2013; Dave and others 2013; Barriocanal and others 2014; Carnero and others 2014; Cui and others 2014; Kambara and others 2014a, 2014b; Ilott and others 2014). However, the mechanisms through which viral infection and inflammatory signals contribute to changes in lncRNA expression are poorly understood.
Given the vast potential for discovery of novel mechanisms of immune response control by lncRNAs, we sought to describe the noncoding transcriptome of human pDCs and develop a model system, whereby functional analyses of lncRNA regulatory processes can be achieved. Collectively, these in vitro studies define the coding and noncoding transcriptome signatures of TLR7- and IFN-I-stimulated human pDCs. We isolated the impact of IFN-I feedback following TLR activation and identify coding and noncoding gene sets sensitive to this influence. Our analysis describes the biotype and genomic distribution of TLR- and IFN-I-modulated lncRNAs, and identifies regional co-expression patterns among DE lncRNAs and nearby coding genes, and their potential transcriptional regulators. This investigation also delineates the concordance in DEG patterns of the CAL-1 line with primary human pDCs. Our characterization of the lncRNA landscape in human pDCs adds a critical element to the growing body of annotation of tissue- and stimulation-specific lncRNA expression profiles and lays a foundation for further study of their functions in immune responses.
Poor conservation of lncRNAs across species and accelerated evolution at these loci warrant comprehensive documentation of expression profiles in human cells (Nitsche and Stadler 2017). High lineage specificity and stimulation-specific expression changes add a further layer of analysis required to fully catalogue human noncoding genomic output. lncRNA expression profiles and individual functional roles have yet to be fully elucidated for the breadth of innate immune stimuli and human cellular subsets. Although coding ISGs have been extensively annotated, noncoding loci were omitted from such studies and very few descriptions of IFN-stimulated lncRNAs exist. Functional examples of lncRNAs induced by TLR activation or IFN-I illustrate the capacity of these molecular players to potently regulate this critical pathway in innate antiviral immunity (Atianand and others 2017). In this study, we present the transcriptome signatures of TLR7- and IFN-I-stimulated human pDCs and identify lncRNAs whose expression is modulated by these stimuli.
In this analysis, we have defined patterns of lncRNA regulation by innate immune TLR activation and IFN-I signaling, subsetting cohorts of transcripts differentially induced or suppressed by TLR or IFN-I signaling. We characterized the genomic distribution of TLR- and IFN-I-modulated lncRNAs and performed an analysis of parallel and inverse expression patterns in their neighboring genes. This analysis revealed co-expression between lncRNAs and neighboring TLR- or IFN-inducible coding genes. This phenomenon may be a product of regulation of these neighboring loci by similar TFs, and our analysis of enrichment of several factors known to control coding gene expression following these stimuli supports this hypothesis. Specifically, we found that IFN- and TLR-induced lncRNA groups are notably enriched for IRF and NF-κB associations, respectively. These analyses suggest that canonical pathways dictating IFN-I- and TLR-driven coding transcriptional changes extend to noncoding loci as well. Co-expression patterns between lncRNAs and coding neighbors are a naturally intuitive phenomenon for lncRNAs derived from alternative splice forms, antisense or intronic transcription of coding genes, as well as those derived from pseudogenes with preserved promoters that parallel their neighboring ancestral ortholog. However, for many of these transcripts, co-expression with a neighboring coding gene may be not only coincidental but also causal, as in the case of antisense lncRNA cis transcriptional control. In addition, co-expressed pseudogenes may contain sequence similarity to their coding orthologues, thereby conferring ceRNA functions that enable relief of repressive elements targeting the coding gene. Thus, the roles of these lncRNA-coding gene neighbor pairs are intriguing points of study and are of particular relevance within contexts of robust transcriptional induction and protein production, such as those of TLR and IFN-I signals. Further study of lncRNAs identified in this analysis, whose expression positively or inversely correlates with that of their coding neighbor, is warranted.
Nuclear lncRNAs within this gene set may act in cis to regulate transcription of their neighboring coding genes (Villegas and Zaphiropoulos 2015). Cytoplasmic lncRNAs co-expressed with a neighboring coding gene may function as ceRNAs for the negative regulatory factors targeting the neighbor gene, thereby facilitating enhanced protein production (Salmena and others 2011). This is especially relevant in the case of pseudogene lncRNAs, which contain high sequence similarity to their coding orthologs (An and others 2017). We validated expression changes of 6 lncRNAs representative of TLR- and IFN-I-modulated signatures and determined their subcellular localization, identifying 3 cytoplasmic regulators whose activities may modulate pDC function. We discovered that expression of lnc-DC, a cytoplasmic noncoding pseudogene specific to DCs and reported to promote STAT3 activity through antagonism of SHP1–STAT3 interaction (Wang and others 2014), is induced following TLR7 and IFN-I stimulation in pDCs and cDCs. Our findings suggest that DCs experience increased sensitivity to STAT3-mediated signals through elevated expression of lnc-DC in the hours subsequent to TLR activation. However, whether this signal-amplifying role might extend to other STAT proteins has not been explored, nor has the contribution of lnc-DC to pDC function been defined. Notably, lnc-DC polymorphisms associate with SLE incidence and levels are diminished in SLE patient blood relative to healthy controls (Li and others 2017), despite elevated IFN-I signaling and pDC activation in this context, indicating possible negative regulatory feedback in chronic exposure states in vivo. We found that a well-described cytoplasmic ceRNA, lnc-ROR, is also induced upon TLR activation in CAL-1 cells. In epithelial tissues, this transcript sponges miR-145 from targets critical to control of pluripotency (Pan and others 2016), suggesting that in pDCs, lnc-ROR may exert ceRNA functions that relieve repression of mRNAs essential to pDC activation. Among the many lncRNAs downregulated following TLR7 stimulation, we identified lnc-1133, a cytoplasmic ceRNA, which has been reported to sponge miRNAs from targets critical to tumorigenesis and proliferation in several cancers (Zhang and others 2015, 2017; Kong and others 2016; Wu and others 2017). We show that in pDC and myeloid lineages, lnc-1133 undergoes rapid and sustained downregulation following TLR stimulation. We did not observe an impact on cell proliferation or viability upon its knockdown (data not shown), thus lnc-1133 may exert other lineage-specific functions in this cell type. Further study of this transcript in hematopoietic lineages and TLR-activated epithelial tissues is warranted to determine the full range of its functions across tissue types.
Restraint of inflammatory responses following TLR engagement is regulated by diverse means, including signaling by anti-inflammatory cytokines as well as transcriptional and posttranscriptional repression, and is critical in limiting tissue damage (Murray and Smale 2012). Several lncRNAs have been shown to function as negative regulators of inflammation (Willingham and others 2005; Yoon and others 2012; Rapicavoli and others 2013; Cui and others 2014; Ouyang and others 2014; Liu and others 2015; Atianand and others 2016; Castellanos-Rubio and others 2016; Jiang and others 2018), including IFN-I production (Kambara and others 2014b; Valadkhan and Gunawardane 2016). In our data set, we observe a trend of substantially more noncoding genes being downregulated than upregulated following TLR stimulation compared to an equivalent distribution of coding DEG directionality. This phenomenon appears in our study to be a broad feature of lncRNAs and is unrestricted to certain lncRNA biotypes. We hypothesize that many of these downregulated transcripts act as negative regulators of cellular processes that are augmented following pDC TLR activation. Relief of the repression enforced by these lncRNAs by their downregulation after TLR activation would facilitate a burst of the cellular activities they restrain. Notably, downregulation of many lncRNAs has been observed in many cancers (Cancer Genome Atlas Research Network and others 2013; Wu and others 2016; Terashima and others 2017; Yang and others 2017), a phenomenon that is often a product of chronic inflammation, as well as following administration of probiotic dietary supplements (Nunez-Acuna and others 2017). While our findings indicate IRF- and NF-κB-driven control of IFN-I- and TLR-inducible lncRNAs, many factors such as miRNA-mediated decay or transcriptional repression may influence the downregulation of lncRNAs following TLR activation. Further study of the factors controlling lncRNA downregulation is warranted and may yield insight into potential transcript functional mechanisms. Many of the downregulated lncRNAs identified in this study may function as a critical brake on inflammatory processes; thus, dysfunction in their upstream regulators might have deleterious consequences due to uncontrolled inflammation. In addition, the TLR7- and IFN-I-induced lncRNAs we have identified could also act as negative regulators of other repressors, as in the case of ceRNAs sponging miRNAs away from their target mRNAs, thereby facilitating increased mRNA stability and translation of inflammatory proteins. We hypothesize that many of the IFN-/TLR-induced and TLR-suppressed lncRNAs identified in this study thus act as critical regulators of the cytokine production, antigen presentation, and migratory activity that ensue once pDCs become activated.
Such regulatory activities have been described for lncRNAs in other pathways such as pluripotency, but in the context of innate immune PRR activation, these roles may contribute to the rapid and robust cytokine production that ensues following ligand engagement. This is particularly relevant to the function of pDCs, whose primary role is production of IFNs and inflammatory cytokines following PRR activation. It has been reported that individual DCs vary greatly in their cytokine production in response to a common stimulus (Shalek and others 2014; Alculumbre and others 2018), and that in pDCs, such individual variability in IFN-I production is independent of the amplifying IFN-I feedback loop (Bauer and others 2016). Differences in the propensity of individual cells to produce inflammatory cytokines could be dictated by the status of regulatory lncRNAs such as those identified in this study, which may be acting as potent repressors of cytokines in otherwise precocious producers. If the activity of such lncRNAs is dysregulated due to polymorphism or other anomalous influences, exaggerated cytokine production in response to infectious or innocuous stimuli may yield the inflammatory foundation upon which the development of autoimmune pathogenesis depends. Further genetic and functional investigations of the lncRNAs identified in this study are thus warranted to identify their regulatory roles in IFN-I and inflammatory cytokine production, or other cellular activities that may contribute to antiviral responses or aberrant pDC activity in systemic autoinflammatory disease.
Our dissection of the contribution of IFN-I to gene expression changes in the context of TLR7 activation uniquely identifies coding and noncoding genes whose upregulation in activated human pDCs critically depends on IFN-I feedback. This phenomenon is reported to be due to the amplification of interferon response factor 7 levels by IFN-I feedback (Prakash and others 2005), which subsequently drives elevated TLR7-dependent gene induction, and our data support this hypothesis (Supplementary Fig. S1E). Disparate activity of these IFN-I-dependent genes may underlie the mechanisms responsible for the effects of IFN-I antagonism therapy in SLE patients (Kirou and Gkrouzman 2013), and the protection from disease pathogenesis provided by IFNAR1−/− or blockade in murine SLE models. Thus, explorations of functional relevance and polymorphic risk factors within these coding and noncoding genes may reveal potential causal mechanisms in SLE and other IFN-I-driven autoreactivities and interferonopathies.
Our description of TLR-/IFN-modulated lncRNAs provides a foundation for identification of novel functional mechanisms and extensive interrogation of potential players in disease. The paucity of pDCs in peripheral blood and the generally low expression levels of lncRNAs are major limitations to functional lncRNA analysis in primary pDCs ex vivo. Through comparison with influenza-infected primary human pDC, we have established that lncRNA signatures in the CAL-1 pDC line resemble primary pDCs and defined their core common elements, demonstrating that this is a suitable model amenable to the biochemical manipulations required for subsequent interrogations of lncRNA function in pDCs. As noncoding loci are sites of evolutionary pressure at host–pathogen interface, we posit that the lncRNA transcripts identified in this study may play vital roles in host defense to viral infection and, when aberrantly expressed or dysfunctional, in autoimmune disease. Further interrogations of the mechanisms utilized by these lncRNAs to regulate pDC function are thus warranted in the in vitro system we have established. Given demonstrated effectiveness of RNA-based interventions in disease (Lieberman 2018), this characterization of the lncRNA landscape during the innate immune response reveals potential novel targets for treatment of viral infection and IFN-driven autoimmunity. We foresee the capacity of lncRNAs defined in this study to act through diverse mechanisms to modulate DC activities, wielding outcome-defining influence during infection and autoimmune disease.
Footnotes
Acknowledgments
This work was supported, in part, by R21AI137956 (R.S.), TL1 TR000422 (R.C.J.), and T32 HL007312 (A.F.). We thank the members of R.S. laboratory for helpful discussions.
Author Disclosure Statement
No competing financial interests exist.