
Abstract
The innate immune response is defined as an immensely complex and sophisticated process aimed at defending the organism against any disturbance in the body homeostasis, including invading pathogens. It requires a close cooperation of a vast amount of different cell types, recognized as inflammatory migrating cells, as well as stationary cells that form tissues. Moreover, innate immune mechanisms require an efficient functioning of various humoral components that exert a significant impact on physiological and pathological processes. Apart from commonly mentioned humoral factors, this group also includes a family of proteins known as adipocytokines that may act as pro- or anti-inflammatory agents or act both ways. Leptin, predominantly characterized as a proinflammatory adipokine, plays a crucial role in endothelium remodeling and regulation, as well as in cell survival and production of numerous cytokines. Adiponectin, similar to leptin, acts on the endothelial cells and the phagocytic properties of immune cells; however, it exerts an anti-inflammatory impact. Resistin has a documented role in the control of angiogenesis and stimulation of proinflammatory mediator generation and release. Furthermore, there are adipokines, ie, visfatin and chemerin, whose participation in the inflammatory processes is ambiguous. This review focuses on the current knowledge on the extensive role of selected adipokines in innate immune response.
Introduction
T
The functioning of the innate immune system requires a close cooperation of numerous cellular components, such as natural killer (NK) cells, dendritic cells, mast cells, eosinophils, basophils, and phagocytic cells, including macrophages and neutrophils. Apart from migratory cells, certain stationary units, such as fibroblasts, endothelial and epithelial cells, are important in modulating the innate immune machinery too. All these cells are the primary line of defense through interaction, detection, and removal of cellular debris, foreign particles, or invading microbes. Nevertheless, depending on which particular cell is activated, different mechanisms, such as apoptosis, phagocytosis, proliferation, or synthesis and release of mediators, are triggered or inhibited (Riera Romo and others 2016; Gasteiger and others 2017).
Likewise, humoral factors are principal for the proper functioning of the innate immune system. Humoral components include various elements such as complement proteins, acute phase proteins, heat shock proteins, and antimicrobial peptides. They contribute to the innate immune response directly by disrupting the membrane of target cells leading to cell lysis, or indirectly by regulating immune cell activities. It should be emphasized that some cytokines also have a substantial impact on the innate immune function. This array of mediators secreted by various innate immune cells includes proinflammatory [ie, tumor necrosis factor (TNF), interleukin (IL)-1β, IL-4, IL-6, IL-12, IL-18, interferon (IFN)-α, IFN-β] and anti-inflammatory factors [ie, IL-1Ra, IL-10, transforming growth factor (TGF)-β]. Furthermore, a family of chemokines plays an essential role in the immune system and strongly regulates the innate processes through controlling the recruitment and positioning of immune cells at the site of inflammation (Riera Romo and others 2016).
Adipocytokines (or adipokines) are biologically active molecules, mainly produced by adipocytes. However, it is important to note that they are also synthesized by other cells present in adipose tissue, including macrophages, neutrophils, or mast cells. This group comprises numerous members, such as leptin, adiponectin, resistin, visfatin, chemerin, vaspin, apelin, omentin (Table 1), and some classical cytokines/chemokines, including TNF, IL-1β, IL-6, CCL2, CXCL8, as well as retinol-binding protein-4 (RBP4), plasminogen activator inhibitor-1 (PAI-1), and insulin-like growth factor-binding proteins (IGFBPs). Considering the role of adipokines, they can be divided into 3 categories: those that have mainly proinflammatory properties (leptin, resistin, TNF, IL-1β, IL-6, CCL2, CXCL8, RBP4, PAI-1), those that are anti-inflammatory (adiponectin, omentin, IL-10), and those that exert a regulatory effect (visfatin, chemerin, apelin, vaspin) (Fasshauer and Blüher 2015). The balance between pro- and anti-inflammatory adipocytokine production appears to play a key role in several aspects of physiology, including energy homeostasis, regulation of body metabolism, angiogenesis, and cardiovascular function. However, many proinflammatory adipocytokines, eg, leptin and resistin, are overproduced with increasing adiposity, while some of these factors with anti-inflammatory properties, such as adiponectin or omentin, are decreased. Dysregulation of adipokine production contributes to “low-grade inflammation” in adipose tissue, a state accompanying obesity (Cao 2014). Additionally, the inflammatory milieu in adipose tissue is characterized by an increased number of macrophages, granulocytes, and adaptive immune cells. The accumulated evidence indicates that innate lymphoid cells also contribute to low-grade inflammation orchestration (Vivier and others 2018). Consequently, adipose tissue inflammation and adipokine imbalance decrease insulin sensitivity in various target organs by impairing insulin signal transduction and lead to the development of type 2 diabetes mellitus. Moreover, a dysregulated adipocytokine synthesis contributes to an increased risk of developing hypertension, arteriosclerosis, hypercholesterolemia, and hypertriglyceridemia, which may be factors for cardiovascular disease. Adipocytokines are also engaged in the pathogenesis of inflammatory diseases such as rheumatoid arthritis, osteoarthritis, and atherosclerosis (Fasshauer and Blüher 2015).
PBMC, peripheral blood mononuclear cell; WAT, white adipose tissue.
There is growing evidence indicating that adipocytokines are crucial humoral components of the immune response. Hence, these factors have been shown to be involved in innate immune mechanisms and inflammatory processes. Adipocytokines influence innate immune cells affecting the expression of adhesion molecules, endothelial permeability, migration, apoptosis, and activate these cells to produce a wide spectrum of various mediators. This review focuses on the current knowledge on the broad role of selected adipokines in innate immune response.
Leptin
Leptin, an adipocytokine-like hormone, is a protein with a molecular mass of 16 kDa. In humans, it is a product of LEP gene, which is a homolog of the murine obese (ob) gene. Leptin is mainly produced by fat cells known as adipocytes. To a lesser extent, it is also synthesized by immune cells, such as T cells, basophils, and mast cells. The circadian rhythm of leptin synthesis is regulated by the timing of feeding and changes in insulin levels. Secretion of this protein is also dependent on sex steroids, growth hormone, and inflammatory mediators, ie, IL-1 and TNF. This adipocytokine is markedly recognized for its role as a hypothalamic modulator of food intake, body weight, and energy expenditure (Park and Ahima 2015).
Leptin exerts its impact by acting on its receptor (LepR or ObR) representing class I cytokine receptor family. Alternative splicing of LepR generates 6 receptor isoforms (LepRa-f) with a different length of intracellular domain. Based on its structural differences, the receptor isoforms are divided into 3 classes: long, short, and soluble. LepR is expressed in many organs, including the hypothalamus, pituitary gland, gonads, spleen, and liver. Many immune cell types, including B and T cells, NK cells, macrophages/monocytes, neutrophils, basophils, eosinophils, dendritic cells, and mast cells, are also characterized by LepR expression. Therefore, numerous studies show that leptin considerably influences immune homeostasis, exerting various effects on the initial immune defense (Pérez-Pérez and others 2017).
Multifaceted Proinflammatory Action of Leptin in Innate Immune Response
A good deal of evidence indicates that leptin has a significant role as a proinflammatory adipokine in the immune response. Firstly, it is known to affect the cell expression of adhesion molecules that are central to the interactions with the vascular endothelium, including the capture of white blood cells from the blood, adhesion, diapedesis, recruitment, and migration of these cells to the sites of inflammation. In endothelial cells, this adipocytokine enhances integrin αvβ5, integrin α4, and intercellular adhesion molecule (ICAM)-1 expression (Schroeter and others 2008; Manuel-Apolinar and others 2013). Leptin also significantly upregulates the surface expression of CD18 and ICAM-1 but suppresses L-selectin and ICAM-3 in eosinophils (Wong and others 2007). It is a potent chemoattractant for neutrophils, inducing a maximal chemotactic response at 50 ng/mL. Nevertheless, this protein is also able to inhibit neutrophil migration in response to classical chemoattractants, ie, N-formyl-methionyl-leucyl-phenylalanine (FMLP), chemokine CXCL8, and polypeptide C5a (Montecucco and others 2006). Leptin seems to be a potent basophil and eosinophil chemoattractant too. Leptin's migratory effect on basophils is both chemotactic and chemokinetic, whereas the impact of this adipocytokine on eosinophil attraction is chemokinetic rather than chemotactic (Wong and others 2007; Suzukawa and others 2011). This protein induces the migration of monocytes and macrophages, but the maximal effect is observed at 50-fold lower concentrations than in the case of neutrophil migratory response (Gruen and others 2007). Moreover, it has been shown that leptin promotes chemotaxis of mast cells (Żelechowska and others 2018). The leptin-stimulated increase of migration is also demonstrated by endothelial (Goetze and others 2002) and dendritic cells (Mattioli and others 2008). Also, leptin stimulation of dendritic cells leads to the upregulation of CCR7 surface expression and enhancement of CCL19-induced dendritic cell migratory response (Mattioli and others 2008).
Leptin has an antiapoptotic effect on monocytes by upregulating p42/44 MAPK prosurvival signaling (Najib and Sánchez-Margalet 2002). It delays spontaneous neutrophil and eosinophil death as it performs its antiapoptotic function by retarding the cleavage of proapoptotic members of Bcl-2 protein family such as Bid and Bax and the mitochondrial release of cytochrome c (Bruno and others 2005). Leptin also enhances NK and dendritic cell survival and protects them from the process of apoptosis, possibly also through modulating Bcl-2 family gene expression (Mattioli and others 2005; Lo and others 2009). This adipocytokine stimulates the proliferation of monocytes. It has been suggested that this effect of leptin may be regulated by an increased production of granulocyte-macrophage colony-stimulating factor (GM-CSF) (Santos-Alvarez and others 1999). It also exerts a stimulatory effect on the proliferation of NK cells (Zhao and others 2003). However, in contrast to short-term leptin stimulation, long-term leptin exposure inhibits NK cell proliferation (Wrann and others 2012).
When considering leptin's role in the mechanisms of the innate immune system, it should be stressed that this adipocytokine can modulate the activity of immune cells. Leptin appears to enhance the activity of phagocytes (Mancuso and others 2004). Furthermore, a significant impact of leptin on monocyte (Dayakar and others 2016) and neutrophil (Caldefie-Chezet and others 2001) reactive oxygen species (ROS) generation was observed. More specifically, leptin-induced ROS production supports phagolysosome formation and oxidative killing of bacteria and parasites by host cells (Mancuso and others 2006; Dayakar and others 2016).
Finally, as for leptin's action in innate immune response, it has been demonstrated that this adipokine stimulates immune cells to generate and release numerous mediators that modulate the course of inflammation in different ways. In general, this protein increases the secretion of proinflammatory cytokines, chemokines, and other mediators. Monocytes/macrophages exposed to leptin produce cytokines, such as TNF, IL-1β, IL-6, IL-18, as well as chemokines CCL2 and CCL3 (Acedo and others 2013; Jitprasertwong and others 2014; Lee and others 2014). It is also observed that macrophage pretreatment with this adipocytokine increases the synthesis of eicosanoids, ie, cysteinyl leukotrienes (cysLTs), LTB4, and prostaglandin E2 in response to calcium ionophore and zymosan (Mancuso and others 2004). In eosinophils, leptin augments the release of IL-1β, IL-6, CCL2, CXCL1, and CXCL8 (Wong and others 2007). This adipokine induces IL-13, IFN-γ, and CCL2 production, while reducing IL-4, IL-6, IL-10, and CCL3 synthesis by murine bone marrow-derived mast cells (BMMCs) (Zhou and others 2015). Moreover, leptin stimulates rat mast cells into generating and releasing proinflammatory mediators, ie, CCL3 and cysLTs (Żelechowska and others 2018). This protein also directly induces basophil and mast cell degranulation and histamine release (Suzukawa and others 2011; Zhou and others 2015; Żelechowska and others 2018). There is ample evidence supporting the presumption that leptin may contribute to the amplification of proinflammatory response by influencing anti-inflammatory mediator synthesis. Namely, it inhibits the production of IL-10 by NK cells (Shirshev and others 2016), dendritic cells (Mattioli and others 2005), and decreases TGF-β production by macrophages (Toossi and others 1996).
Adiponectin
Adiponectin is a 30-kDa protein that in humans is encoded by the ADIPOQ gene. This adipocytokine is produced mainly by adipocytes. Nowadays, it is known that adiponectin is also synthesized by mononuclear cells, such as lymphocytes and neutrophils (Crawford and others 2010). There are 2 various forms of adiponectin: a full-length protein and a globular adiponectin containing the C-terminal domain resulting from a cleavage process, which is mediated by monocyte/neutrophil elastase. Both forms of adiponectin participate significantly in processes related to the immune system; however, the high-molecular-weight (HMW) variant of globular form orchestrates adiponectin-associated activity in the liver, while the low-molecular-weight (LMW) variant is more important for brain functioning (Simpson and Whitehead 2010).
Adiponectin acts through 2 different receptors. AdipoR1 is expressed mainly in the skeletal muscles, whereas AdipoR2 is found in the liver. Expression of adiponectin receptors is also identified in monocytes, neutrophils, NK cells, and B cells (Pang and Narendran 2008). Furthermore, modulation of the expression of these receptors is strongly dependent on various cytokines, ie, TNF, IFN-γ, IL-6, and leptin (Chen and others 2006).
Anti-Inflammatory Action of Adiponectin in the Innate Immune System
Although the role of adiponectin in the immune system has not been definitely established, increasing evidence indicates that it exerts predominantly anti-inflammatory properties. It acts as an endogenous modulator of endothelial cell function as it suppresses TNF-induced cell surface expression of adhesion molecules such as ICAM-1, vascular cell adhesion molecule (VCAM)-1, and E-selectin (Ouchi and others 1999). Interestingly, in vivo studies on leukocyte–endothelium interactions in adiponectin-deficient (Ad−/– ) mice demonstrate that adiponectin deficiency is associated with an increased leukocyte rolling and adhesion in the microcirculation (Ouedraogo and others 2007). This adipokine potently suppresses endothelial cell migration (Bråkenhielm and others 2004) and inhibits vascular endothelial growth factor-stimulated endothelial cell migration through cAMP/PKA-dependent signaling (Mahadev and others 2008). It is also indicated that adiponectin effectively inhibits eotaxin-induced eosinophil chemotactic response without affecting cell viability (Yamamoto and others 2013).
There are still controversies about the function of adiponectin in apoptosis. Adiponectin induces apoptosis in monocytes by downregulating the expression of an antiapoptotic gene, ie, Bcl-2, without affecting the expression of Bax, Bak, and p53 genes (Yokota and others 2000). In macrophages, this protein stimulates the generation of intracellular and mitochondrial ROS. The same study reveals that the expression ratio of Bcl-2 to Bax is reduced in adiponectin-treated macrophages. Overexpression of Bcl-2 inhibits adiponectin-induced apoptosis in macrophages and also reduces both intracellular and mitochondrial ROS generation stimulated by adiponectin treatment (Akifusa and others 2009). On the contrary, adiponectin reduces apoptosis in resting neutrophils, decreasing caspase-3 cleavage and maintaining Mcl-1 expression by stabilizing this antiapoptotic protein (Rossi and Lord 2013b). Adiponectin also inhibits palmitate-induced apoptosis by the suppression of ROS generation in endothelial cells (Kim and others 2010). The anti-inflammatory effects of adiponectin involve the suppression of IL-2-induced NK cell cytotoxic activity (Kim and others 2006).
Adiponectin interferes with the macrophage and neutrophil phagocytic function. Treatment of cultured macrophages with adiponectin notably inhibits their phagocytic activity and their production of TNF in response to lipopolysaccharide (LPS) stimulation (Yokota and others 2000). Consequently, adiponectin inhibits neutrophil phagocytic ability as well (Rossi and Lord 2013a).
This adipokine regulates the activity of immune cells by the induction of anti-inflammatory mediator synthesis. It stimulates the production of IL-1Ra and IL-10 in monocytes, macrophages, and dendritic cells (Wolf and others 2004). It is worth noting that the production of TNF, IL-6, and IFN-γ in LPS-stimulated macrophages is significantly decreased by adiponectin pretreatment (Wulster-Radcliffe and others 2004). Adiponectin exerts its anti-inflammatory effects on endothelial cells by suppressing TNF-stimulated CXCL8 mRNA and protein expression (Kobashi and others 2005).
There is some evidence that different isoforms of adiponectin perform distinct biological functions. Full-length adiponectin affects IL-6 and TNF production in macrophages differently by upregulating the protein synthesis of these cytokines, but not their mRNA expression. However, globular adiponectin appears to have proinflammatory properties as it substantially enhances IL-6 and TNF mRNA and protein secretion in macrophages. Hence, some authors indicate that the mechanism of action of full-length adiponectin in the transcription and translation of IL-6 and TNF is different (Kyriazi and others 2011). HMW adiponectin form can induce the production of CCL2 and CXCL8 by peripheral blood mononuclear cells (PBMCs) and endothelial cells, whereas LMW adiponectin does not induce this chemokine synthesis (Song and others 2009).
Resistin
Resistin is a 12.5-kDa cysteine-rich member of a family called resistin-like molecules (RELMs). In rodents, RELM gene family consists of 4 members, including resistin, RELMα, RELMβ, and RELMγ, while in humans only 2 genes are discovered, for resistin and RELMβ (Adeghate 2004). This protein is produced by brown and white adipose tissue (WAT), pancreas, hypothalamus, pituitary and adrenal glands. It has also been identified in macrophages, PBMCs, and bone marrow cells. Moreover, macrophages are the primary source of resistin in humans, while in rodents it is produced entirely by adipocytes. Murine resistin is involved in the pathogenesis of obesity-mediated insulin resistance and type 2 diabetes. However, the function of resistin as a molecule that links obesity to type 2 diabetes in humans remains ambiguous (Park and Ahima 2013).
There is little information available on the receptor for resistin and signaling mechanisms. So far, adenylyl cyclase-associated protein-1 (CAP1) has been identified as a resistin receptor that mediates the proinflammatory effects of human resistin in monocytes and WAT cells. An isoform of decorin and tyrosine kinase-like orphan receptor-1 have been proposed as receptors for murine resistin (Daquinag and others 2011; Sánchez-Solana and others 2012).
Resistin as a Potent Stimulator of Adhesion Molecule Expression and Cytokine Production
There is a growing body of evidence that resistin is involved in the regulation of innate immune responses, including the inflammatory processes. More specifically, resistin has been postulated to play a meaningful role in endothelial function. This adipokine increases the expression of adhesion molecules, including ICAM-1, VCAM-1, platelet endothelial cell adhesion molecule-1, and P-selectin in endothelial cells (Kunnari and others 2009; Manduteanu and others 2010; Hsu and others 2011). Consequently, it augments monocyte adhesion to endothelial cells (Hsu and others 2011). Resistin increases endothelial permeability through enhanced oxidative stress and ROS production (Jamaluddin and others 2013). Furthermore, resistin increases monocyte–endothelial cell adhesion by boosting integrin α4β1 expression on monocytes (Cho and others 2011).
Resistin induces endothelial cell migration in a dose-dependent manner with the maximal effect at 40 ng/mL. Moreover, this adipokine at a concentration of 25 ng/mL significantly attenuates leukocyte chemotaxis. Nevertheless, the inhibitory effect of resistin on leukocyte chemotaxis is partially reversible, and lower resistin concentration might improve leukocyte chemotactic response (Cohen and others 2008).
Although limited data indicate the active participation of resistin in apoptosis of immune cells, it has been shown that it may indirectly influence cell survival by decreasing ROS production, which regulates the apoptotic functions of macrophages and neutrophils (Chao and others 2015). In leukocytes, resistin is reported to impair the oxidative burst induced by Escherichia coli (Cohen and others 2008).
Resistin augments the expression of proinflammatory cytokines such as TNF-α, IL-1β, and IL-6 in PBMCs (Bokarewa and others 2005), and increases the secretion of TNF and IL-12 from human and rodent macrophages (Silswal and others 2005). Moreover, in macrophages, resistin leads to an increased expression of interleukins, ie, IL-5, IL-6, IL-23, IL-27, and chemokines such as CCL1, CCL4, and CXCL1 (Tucureanu and others 2016), while the production of IL-1β was significantly reduced due to lowered activation of inflammasome, which is responsible for IL-1β and IL-18 maturation and secretion (Chao and others 2015). Resistin can also directly stimulate WAT to express mediators at both gene and protein levels, ie, IL-6 and CXCL8, or exclusively at gene expression, ie, CCL2 (Nagaev and others 2006). Resistin induces CCL2 production in endothelial cells (Verma and others 2003).
Visfatin
Visfatin is a 52-kDa protein, synthesized and released preferentially in visceral adipose tissue. Within adipose tissue, visfatin is also produced and secreted by inflammatory cells, including macrophages whose infiltration in this tissue is notably increased in obesity. The bone marrow, liver, as well as the muscles are the sources of the highest expression of visfatin; however, its presence is also detectable in the brain, lungs, spleen, kidneys, and testicles (Sonoli and others 2011). There is sufficient data indicating upregulated concentrations of visfatin in humans suffering from diabetes mellitus. Undoubtedly, visfatin plays a crucial role in the regulation of pro- and anti-inflammatory cytokine production; thus, it may contribute to several inflammatory disorders. The specific receptor of visfatin has not been identified yet, but it is believed that this adipocytokine shows proinflammatory activity by insulin receptor signaling pathways (Jacques and others 2012).
Visfatin as a Cell Migration-Stimulating Adipokine and Chemokine Production Regulator
The involvement of visfatin in the innate immune system is suggested in numerous reports. However, it is still debated whether it acts as a proinflammatory or anti-inflammatory adipokine. Undoubtedly, visfatin with its proinflammatory effects may promote the expression of cell adhesion molecules ICAM-1, VCAM-1 (Kim and others 2008), or E-selectin in endothelial cells (Lee and others 2009). This adipokine can also promote endothelial cell migration in a concentration- and time-dependent manner (Adya and others 2008). In addition, visfatin may serve directly as a chemotactic factor leading to an increased motility of monocytes in a dose-dependent manner. Moreover, visfatin may indirectly influence the migration of various cells by inducing the expression and release of several chemokines in endothelial cells (Singh and Benencia 2017) and fibroblasts (Meier and others 2012).
Furthermore, visfatin enhances IL-1β-dependent induction of IL-6 and CD36 expression, and consequently leads to a more rapid differentiation of monocytes into macrophages. Moreover, the same study revealed that this adipocytokine enhances the phagocytic capacity of differentiating monocytes (Yun and others 2014). Other studies also confirm the effect of visfatin on phagocytosis of human monocytes, specifically by a pathway associated with the mannose receptor (Moschen and others 2007).
Undeniably, extensive data indicate that visfatin is a potent antiapoptotic factor. It augments the neutrophil proinflammatory function, inhibiting their apoptosis in a dose-dependent manner by decreasing the activity of caspase-3 and 8 (Jia and others 2004) and enhancing oxidative burst activity and ROS generation (Malam and others 2011). Whereas the inhibition of visfatin exerts an opposite effect (Roberts and others 2013). Furthermore, it prevents macrophages from programmed cell death (Li and others 2008).
The proliferation of immune cells is another crucial ability among the already recognized functions of visfatin. It indirectly triggers peripheral blood lymphocyte proliferation through the stimulation of monocytes in a dose-dependent manner (Moschen and others 2007). Moreover, it has been documented that this adipocytokine may enhance endothelial cell proliferation as well (Adya and others 2008).
Visfatin can also contribute to the functioning of innate immune mechanisms through its immunomodulatory properties on immune cell cytokine/chemokine production. In PBMCs, it induces a dose-dependent production of both proinflammatory (IL-1β, IL-6, TNF) and anti-inflammatory (IL-1Ra, IL-10) cytokines. Nevertheless, the most pronounced effects are observed for IL-6 synthesis, a mediator with a pleiotropic effect on the immune response and inflammation (Moschen and others 2007). Visfatin upregulates the gene expression of various mediators, including IL-6, CCL2, CXCL2, and CXCL8, as well as it augments the release of CXCL2 and CXCL8 in endothelial cells (Singh and Benencia 2017). In fibroblasts, it also upregulates several chemokines at gene and protein levels, ie, CXCL1–3, CXCL5, CXCL6, CCL2, CCL13, and increases the release of CXCL8, CCL5, CCL20, and IL-6 (Meier and others 2012).
Chemerin
Chemerin is an adipokine that constitutes the G protein-coupled receptor CMKLR1 (ChemR23) ligand. This protein is highly expressed at mRNA level in adipose tissue, liver, and kidneys, or to a lesser extent also in heart, spleen, lungs, testicles, and ovaries (Roman and others 2012). Chemerin exerts its impact through a specific receptor known as CMKLR1 or ChemR23, which is expressed in adipocytes (Bozaoglu and others 2007). Moreover, in contrast to the chemerin itself, CMKLR1 is present in skeletal muscle cells (Sell and others 2009), endothelial cells (Kaur and others 2010), and, to a greater extent, in lungs, spleen, and heart, but is barely expressed in liver, kidneys, and testicles (Bozaoglu and others 2007). The activated receptor regulates adipogenesis, the metabolism of adipocytes or insulin sensitivity, and glucose uptake in skeletal muscle cells (Sell and others 2009). Furthermore, since both chemerin and its receptor are upregulated in differentiated adipocytes, it is suggested that this adipokine stimulates CMKLR1 in an autocrine/paracrine way (Roh and others 2007).
Chemotactic Chemerin in Innate Immune Mechanisms
Undoubtedly, chemerin plays an essential part in the innate immune system; however, it is hard to describe it explicitly as a pro- or anti-inflammatory factor. Despite its well-known role in chemotaxis, some reports indicate the stimulating role of chemerin in the inflammatory process, also proving that this protein can inhibit proinflammatory mechanisms. Chemerin stimulates anti-inflammatory processes through the downregulation of TNF-induced VCAM-1 expression in endothelial cells, and contributes to the anti-inflammatory NO release (Yamawaki and others 2012). In contrast, some data demonstrate the proinflammatory activity of this adipokine, due to an increased macrophage adhesion to VCAM-1 through the regulation of integrin α4β1. This effect, however, is triggered by the stimulation of ChemR23 (Hart and Greaves 2010). Chemerin is mostly recognized as a potent chemotactic factor that stimulates the directional migration of cells expressing ChemR23, including NK cells, dendritic cells, and monocytes/macrophages (Carlino and others 2012). Moreover, chemerin acts selectively on plasmacytoid dendritic cell chemotaxis rather than on myeloid dendritic cells due to a lack of CMKLR1 in the latter ones (Zabel and others 2005). However, there are data denying this statement and proving chemerin receptor expression in plasmacytoid and myeloid dendritic cells. The data also reveal the migratory response in both dendritic cell populations (Vermi and others 2005).
More data suggest chemerin to have the ability to stimulate phagocytosis, not only the autophagy of endothelial cells but also to engulf apoptotic neutrophils by other immune cells. More accurately, chemerin regulates the activity of macrophage phagocytic functions against pathogen-derived structures and apoptotic cells through the activation of ChemR23 (Laranjeira and others 2018).
Apart from the stimulation of endothelial cell proliferation, chemerin regulates the differentiation of various stationery cells, which may contribute to the innate immune response. Firstly, this adipokine enhances both proliferation and migration of vascular smooth muscle cells resulting in vascular remodeling (Kunimoto and others 2015). Secondly, in contrast to endothelial and smooth muscle cells, chemerin decreases the differentiation of fibroblasts (Eisinger and others 2012).
Undoubtedly, chemerin plays a pivotal role in the inflammatory process initiation and amplification through the regulation of cytokine production and release. Moreover, this adipokine upregulates its receptor on endothelial cells, since TNF, IL-1β, and IL-6 lead to a higher expression of ChemR23 in a dose-dependent manner (Kaur and others 2010). However, in an LPS-induced lung inflammation mouse model, chemerin revealed its anti-inflammatory function due to a decreased production of proinflammatory proteins such as TNF, IL-1β, IL-6, and CXCL1 (Luangsay and others 2009). Furthermore, synovial fibroblast stimulated with chemerin expresses higher levels of CCL2 (Eisinger and others 2012).
Other Adipokines in the Innate Immune Response
Several lines of evidence suggest that other adipocytokines such as vaspin, apelin, and omentin can also modulate the mechanisms of the innate immune response. Particularly, they influence endothelial cell biology and activity. Vaspin decreases TNF-induced expression of adhesion molecules, such as ICAM-1, VCAM-1, and E-selectin (Jung and others 2014). Likewise, omentin decreases TNF-induced expression of ICAM-1 and VCAM-1 (Zhong and others 2012). On the contrary, apelin increases the expression of ICAM-1 and VCAM-1 on the endothelium (Lu and others 2012). Furthermore, vaspin and omentin augment the proliferation and inhibit the apoptosis of these cells (Maruyama and others 2012; Nakatsuka and others 2013). Besides, these adipokines may affect macrophage functions. Vaspin reduces apoptosis of macrophages (Lin and others 2016), and apelin downregulates chemotaxis and suppresses the phagocytic activity of these cells (Izgüt-Uysal and others 2017), whereas omentin stimulates the differentiation of macrophages into the anti-inflammatory M2 phenotype (Watanabe and others 2016). Vaspin decreases the release of TNF, CCL2, CXCL8, and apelin suppresses TNF and IL-6 production in these cells (Izgüt-Uysal and others 2017). Omentin reduces LPS-induced mRNA expression of TNF and IL-6 (Hiramatsu-Ito and others 2016) and impairs TNF-induced production of IL-1α, IL-1β, and IL-6 in macrophages (Rao and others 2018). Moreover, Saalbach and others (2016) showed that vaspin reduces the release of proinflammatory mediators from monocytes (TNF, IL-1β), neutrophils (CXCL8, ROS), and dendritic cells (TNF, IL-6, CCL2, CXCL8).
The role of classical cytokines/chemokines, representing the adipocytokine family, in the innate immune response orchestration is pleiotropic. Indeed, these cytokines/chemokines are essential humoral factors that regulate and control the course of immunological processes, and their role is well known and defined. It is worth noting that these cytokines/chemokines regulate the mechanism of cell extravasation to the tissue. They influence blood vessels through the modulation of their dilation and enhance adhesion molecule expression on endothelium, as well as other cells involved in the inflammatory process. Classical cytokines/chemokines also influence blood vessels through the modification of vascular permeability. Consequently, these mediators may contribute to the regulation of margination, rolling, activation, tight adhesion, diapedesis, and chemotaxis of blood cells to the site of inflammation. Furthermore, classical cytokines/chemokines affect the phagocytic abilities of cells and regulate the generation and release of ROS from phagocytes. It should be stressed that the innate immune system cells use pattern recognition receptors to identify pathogens by engaging pathogen-associated molecular patterns, and this recognition may be substantially regulated by classical cytokines/chemokines. These factors also differently affect the survival and apoptosis of cells involved in the inflammatory process. Finally, they activate innate immune cells to produce and release various mediators that modulate the innate immune response in different ways.
Conclusion
The innate immune processes are considered highly intricate and advanced mechanisms in which all components must precisely cooperate to exert the desired effect. This response is orchestrated side-by-side by various immunological cells in a way that ensures a proper functioning of innate immune mechanisms. Likewise, different humoral factors, ie, cytokines, chemokines, complement components, acute phase proteins, or heat shock proteins, contribute to the physiological control and organization of immune processes. Considering the broad spectrum of adipokine activities, it can be hypothesized that they are one of the major components in the humoral constituent network having a significant impact on the functioning of the immune system and the regulation of homeostasis. Some of them may be considered to be mainly proinflammatory factors, such as leptin and resistin (Table 2), or anti-inflammatory, such as adiponectin and omentin (Table 3), or regulatory, such as visfatin, chemerin, vaspin, and apelin (Table 4), acting on the various mechanisms of the immune response, ie, the activity of endothelium and adhesion molecule expression, chemotaxis, apoptosis, phagocytosis, as well as mediator production and release (Fig. 1). Although the role of adipokines in organism defense processes is underestimated, based on the presented facts, it can be stated that they are physiologically and immunologically important. Hence, adipocytokines may play a prominent role in the host defense against bacterial, viral, fungal, and parasitic infections. Therefore, it is of prime interest to provide a new insight into the role of adipokines and understand their mechanisms of action so as to draw effective therapeutic implications.
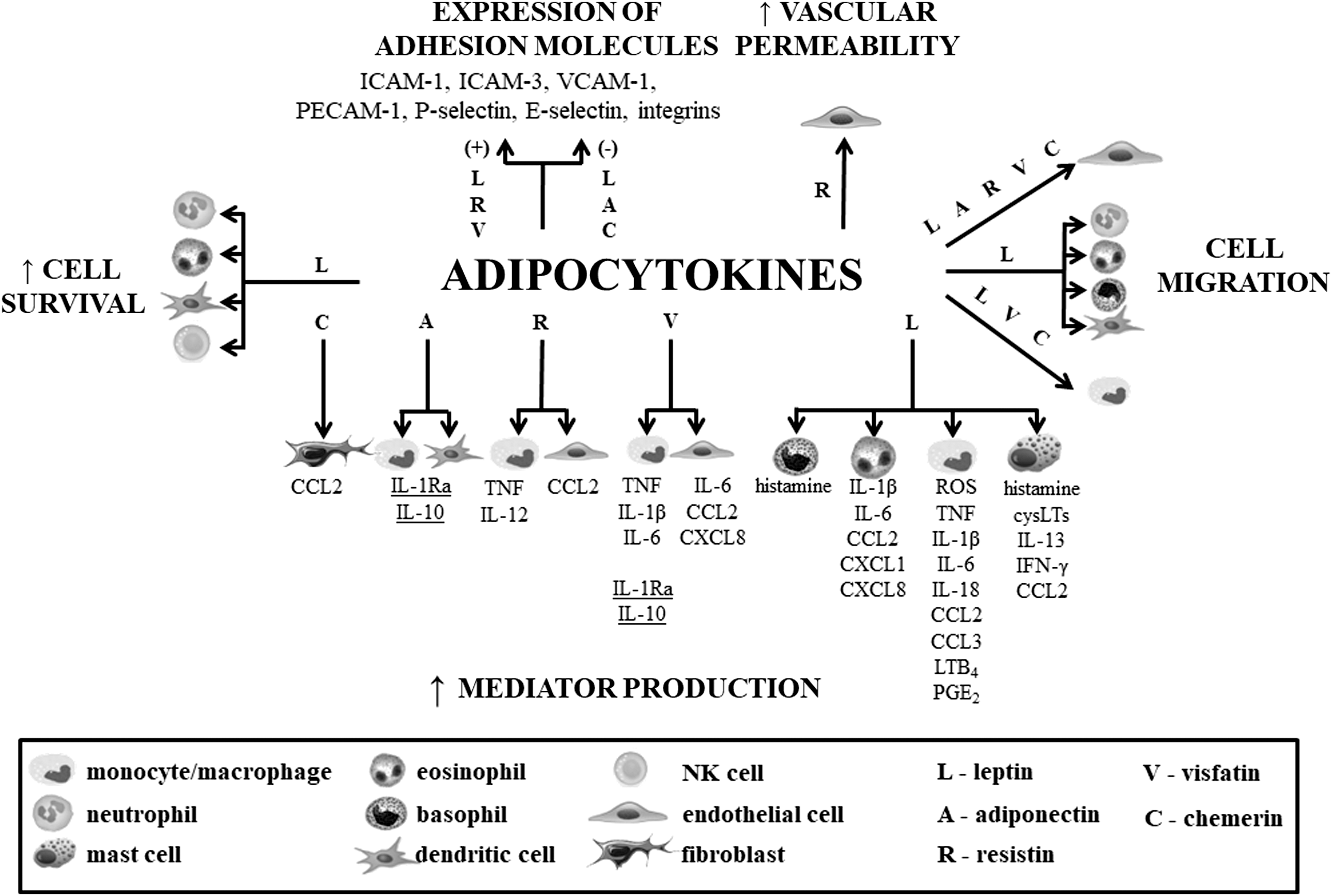
Effects of adipocytokines on innate immunity cells. Underlined mediators constitute the classic anti-inflammatory cytokines.
BMMC, bone marrow-derived mast cell; cysLT, cysteinyl leukotrienes; ICAM, intercellular adhesion molecule; IFN, interferon; IL, interleukin; NK, natural killer; PECAM, platelet endothelial cell adhesion molecule; PGE2, prostaglandin E2; TGF, transforming growth factor; TNF, tumor necrosis factor; VCAM, vascular cell adhesion molecule.
ROS, reactive oxygen species.
Footnotes
Acknowledgment
This work was supported by the grants no. 502-03/6-164-01/502-64-105 and 503/6-164-01/503-66-001 from the Medical University of Lodz, Poland.
Author Disclosure Statement
No competing financial interests exist.