
Abstract
This study investigated the effect of various magnetic flux densities of extremely low frequency electromagnetic fields (ELF-EMF) on expression of T-box transcription factor (T-bet) and GATA binding protein-3 (GATA-3) genes in the spleen and thymus of rats injected with human serum albumin (HSA). Moreover, serum levels of interferon (IFN)-γ and interleukin (IL)-4 were evaluated at two phases, that is, prestimulation and poststimulation with HSA. Eighty rats were separated into five groups, and four groups were exposed daily to 50 Hz EMF of 1, 100, 500, and 2000 μT magnetic flux densities for 60 days. To activate the immune system, 100 μg HSA was intraperitoneally injected into each rat on days 31, 44, and 58 of the regimen. Splenic and thymic T-bet and GATA-3 messenger RNA (mRNA) expression on day 61 was evaluated by reverse transcription quantitative PCR. Serum IFN-γ and IL-4 (in blood on day 31 before HSA and again on day 61) levels were evaluated by enzyme-linked immunosorbent assay. Expression of T-bet and GATA-3 mRNA was decreased in the spleen in hosts exposed to densities of 1 and 100 μT. Serum IFN-γ and IL-4 levels were also significantly decreased in 100 μT-exposed rats, but only at the prestimulation phase. From these findings, it appears that (30 and 60 days) ELF-EMF exposure could suppress the expression of some key genes associated with T helper (Th) cells and on some of their associated functions, that is, the ability to generate (in some cases, spontaneously) select cytokines. Whether this is attributable to effects on Th1/Th2 levels in the hosts and/or due to potential effects of the EMF on cellular functions remains to be determined.
Introduction
H
Recently, some studies investigated the effects of ELF-EMF on human health with special focus on the relationship with immune system functions (Silkstone and Wilson 2016; Varani and others 2017). Changes in the immune system components are key factors in almost all disease conditions. Unintentional environmental sources or intentional therapeutic sources of ELF-EMF can impact immune system functions (Quaglino and others 2000; Pesce and others 2013). The balance between T helper (Th)1, Th2, Th17, and regulatory T (Treg) cells plays a critical role in regulating the immune system function during activation against antigens. On the contrary, imbalance in their functions and cytokine production causes inflammatory and autoimmune diseases. Interestingly, each of these Th subsets is characterized and distinguished based on its own signature cytokines and unique designing genes (Jadidi-Niaragh and Mirshafiey 2011; Raphael and others 2015).
In our previous work, the effect of ELF-EMF on Th17 and Treg cell signature cytokines and related genes was reported. Therefore, in this study, the expression of T-box transcription factor (T-bet), transcription factor GATA binding protein-3 (GATA-3), serum interferon (IFN)-γ, and interleukin (IL)-4 associated with Th1 and Th2 subsets, respectively, was investigated (Mahdavinejad and others 2018).
T-bet expression in naive CD4 + T cells causes them to differentiate into Th1 cells, in turn producing very important cytokines such as IFN-γ, IL-1, and tumor necrosis factor (TNF) to help induce cellular immune response. Among these cytokines, IFNγ is the signature cytokine for Th1 and has a key role in host defenses against intracellular pathogens and tumors (Zamani and others 2009; Hoyler and others 2013; Fujiwara 2017). In contrast, Th2 cells express GATA-3 and regulate humoral responses, in part, by producing IL-4, −5, −9, and −13. GATA-3 acts as a major activator of Th2 cell differentiation in the thymus and peripheral tissues. IL-4 is a signature cytokine of Th2 cells and is a key factor to host responses in antiparasitic immunity and allergic diseases (Tindemans and others 2014; Sahoo and others 2016; Fujiwara 2017).
The spleen and thymus represent key primary organs of the immune system, and T cell numbers and functions (as well as that of lymphocytes in general) are greatly influenced by the status of these organs (Luo and others 2016).
Jonai and others investigated cytokine production by human peripheral blood mononuclear cells (hPBMC) under exposure to 50 Hz EMF. These investigators found that IFN-γ production from the hPBMC was decreased due to ELF-EMF exposure at density of 10000 μT (Jonai and others 1996). In addition, in other in vitro experiments, splenic lymphocytes of mice have been exposed to ELF-EMF with different densities of 250, 500, and 1000 μT for 6 hr. The results revealed that expression of T-bet, GATA-3, and supernatant levels of IFN-γ and IL-4 were not altered (Luo and others 2016).
In an in vivo experiment that was carried out in our laboratory in 2013, we found that exposure of rats to 100 μT density of ELF-EMF for 3 months had no effect on serum IFN-γ and IL-4 (Salehi and others 2013). Boscolo and others also studied the effect of ELF-EMF exposure on eight women who were employees in a museum. They showed that exposure (range of 0.2–3.6 μT) for 20 hours a week caused a decrease in serum and supernatant levels of IFN-γ. Meanwhile, there was no statistical change in the levels of IL-4 (Boscolo and others 2001).
While there have been some studies that examined effects of ELF-EMF on serum cytokine production as mentioned above, to our knowledge, none has examined in vivo effects on the expression of genes for T-bet and GATA-3 - key factors that regulate eventual differentiation of T cells into the major Th type 1 and 2 subtypes. Furthermore, there have been very few studies that specifically examined effects in vivo of a wide range of magnetic flux densities and serum IFN-γ and IL-4. Thus, the present study was undertaken to examine potential effects of 50 Hz EMF with different magnetic flux densities (ie, 1–2000 μT) on T-bet and GATA-3 gene expression in the spleen and thymus of exposed rats, whose immune system had been stimulated with antigen. From a functional standpoint, immune system integrity was evaluated in these exposed hosts by monitoring formation of IFNγ and IL-4 in situ (ie, via measures of circulating forms) as potential reflections of any changes in Th1 and Th2 cells arising from the exposures. This is not to say that any changes observed in circulating IFNγ and IL-4 would mean that ELF-EMF altered Th1 and Th2 cell levels; it could mean that force led to alterations in cell functions alone, or in combination with changes in cell numbers.
Materials and Methods
Animals
Eighty Wistar rats (male, 202.5 [ ± 7.5] g, 8 weeks of age) were purchased from the animal facilities of Hamadan University of Medical Sciences (Hamadan, Iran) and housed in specific pathogen-free (SPF) facilities maintained at 21–22°C and a relative humidity of 55–65%, with a 12-hr light/dark cycle. All rats had ad libitum access to standard rodent chow and tap water. All protocols applied in these studies were approved in advance by the Ethics Committee for Hamadan University of Medical Sciences (Hamadan, Iran). All rats were allowed to acclimate for 1 week before initiation of any exposures (Salehi and others 2013; Mahdavinejad and others 2018).
Electromagnetic exposure system and exposure protocol
The electromagnetic exposure solenoids used here were polyvinyl chloride (PVC) tubes (2 m long, 20 cm radius) coiled with 2-mm-diameter copper wires. Each solenoid had a different number of turns and was provided different electrical potentials and currents. By this, these exposure systems were able to produce 50 Hz EMF with magnetic flux densities of 1, 100, 500, or 2000 μT. An ELF-EMF survey meter (HI-3604, Holaday Industries, Eden Prairie, MN) was used to measure densities. Background magnetic flux density in the room where those rats were maintained was consistently ≈0.07 ± 0.03 μT (Salehi and others 2013; Mahdavinejad and others 2018).
For the exposures, rats were randomly allocated into four ELF-EMF-exposed and one sham-exposed control group (n = 16/group). The rats in the four exposure groups were then exposed daily for a total of 2 hr (ie, from 8–10 AM) to magnetic flux densities of 1, 100, 500, or 2000 μT. Control rats were placed in the exposure device for the same amount of time, but with no electricity on, unplugged. After 1 mo of exposures, blood samples were collected from the retro-orbital plexus of each rat; these materials were designated “prestimulation phase” samples. To then stimulate the immune system of each host, 100 μg human serum albumin (HSA) (Sigma, Munich, Germany) in a volume of 0.5 ml was injected intraperitoneally into each rat on days 31, 44, and 58 of the ELF-EMF exposure regimen (ie, days 1, 14, and 28 of stimulation phase). The aim of the HSA injection of rats was to stimulate the rats' immune system. According to the literature and concept of different Th subset patterns, they can be produced after activation from naive CD4 + T cells. It is important to note that rats were treated with the EMF throughout the entire HSA treatment period; also, there were no rats in this study that did not receive HSA. All the rats were weighed and euthanized (via ether asphyxiation) on day 61 (ie, 1 day after the final exposure). On death, blood was immediately collected from the vena cava of each rat and serum was then generated, isolated, and stored at −70°C for later analysis. At necropsy, the thymus and spleen of each rat were aseptically removed, blot-dried, and weighed (Mahdavinejad and others 2018).
Cytokine measurements
Serum levels of IFN-γ and IL-4 were measured using commercial rat IFN-γ and rat IL-4 ELISA Kits (Eastbiopharm, Hangzhou, China), following the manufacturer's protocols. The sensitivity of each kit was 0.51 pg/mL. All samples were analyzed in duplicate.
Reverse transcription quantitative PCR analyses
Five and 10 mg of the spleen and thymus tissues of each rat, respectively, were used to extract total RNA. Each tissue was homogenized in 1 mL cold kit-provided reaction solution (RNX-Plus; CinnaGen, Tehran, Iran) and RNA was then extracted using the kit, according to the manufacturer's instructions. After checking for isolate concentration and purity via absorbances at 260/280 nm with a NANO-200 microspectrophotometer (A&E Laboratories, Guangzhou, China), a total of 5 μg RNA/organ/rat in 1 μL was used as a template to generate single-strand complementary DNA (cDNA). This process utilized 10X buffer (2 μL), 40 μM oligonucleotide (dT) primer (1 μL), 10 mM deoxynucleotide (dNTP) mix (1 μL), 14.5 μL nuclease-free water, and 100 U/mL Moloney Murine Leukemia Virus (M-MuLV, CinnaGen) reverse transcriptase enzyme (0.5 μL). Total reaction volume in each case was 20 μL. The thermal program for cDNA amplification was 5 minutes at 65°C, 60 minutes at 42°C, and terminated at 85°C for 5 minutes (Zamani and others 2006; Mahdavinejad and others 2018).
T-bet and GATA-3 messenger RNA (mRNA) expression in the spleen and thymus was then evaluated using reverse transcription quantitative PCR. In brief, each generated cDNA was amplified using SYBR Green PCR Mix (Bioron GmbH, Ludwigshafen, Germany) and specific primers designed by AlleleID 7.6 software (Table 1). The comparative threshold cycle (Cq) method and a reference gene (β-actin) were used to normalize expression of each gene (2−ΔCq = 2−[Cq (T-bet or GATA-3) – Cq β-actin]). Then, the relative expression of mRNA for each target gene was calculated by dividing the 2−ΔCq each group by the 2−ΔCq control group (Schmittgen and Livak 2008). For each reaction, the thermal program used was 95°C for 10 minutes at the beginning, followed by 38 cycles of 10 seconds at 95°C, 60 seconds at 58°C for β-actin and T-bet, and 61°C for GATA-3, and terminated at 20 seconds at 72°C.
Specific Primers Used for Amplifications in Reverse Transcription Quantitative PCR
F, forward; R, reverse.
Statistical analysis
All data expressed as mean ± standard deviation. Data for T-bet and GATA-3 mRNA expression and for serum IFN-γ and IL-4 levels among the test groups were analyzed using a one-way analysis of variance (ANOVA). For further analysis, and to determine if there are significant differences between groups, a Tukey's post hoc test was performed. A P < 0.05 was uniformly accepted as significant. All data were analyzed using SPSS.16 software (IBM, Armonk, NY).
Results
Two rats, from 1 and 500 μT groups, died during the exposure to ELF-EMF. The data of the relative expression for T-bet and GATA-3 mRNA in the HSA-stimulated hosts revealed that T-bet and GATA-3 expression was altered only in the spleen by exposure to ELF-EMF (Fig. 1A, B). The data did not reveal such changes in thymic T-bet or GATA-3 expression, regardless of magnetic flux density (Fig. 1C, D). Analysis of relative splenic expression for T-bet mRNA in these HSA-injected rats indicated that T-bet expression was significantly reduced in the 1 (0.86 [±0.11]; P = 0.002) and 100 μT (0.81 [±0.11]; P < 0.001) groups compared with control (value = 1). There were no dose-related effects of flux on this parameter, as exposure to 500 and 2000 μT yielded nonsignificant changes in expression (Fig. 1A). With GATA-3, there was also significant reduction in expression to 0.88 [±0.10] (P = 0.007) in the 1 μT and 0.83 [±0.10] (P < 0.001) in 100 μT groups compared with levels in the control (value = 1) (Fig. 1B). All higher densities had no effect on splenic GATA-3 expression in the HSA-stimulated rats.
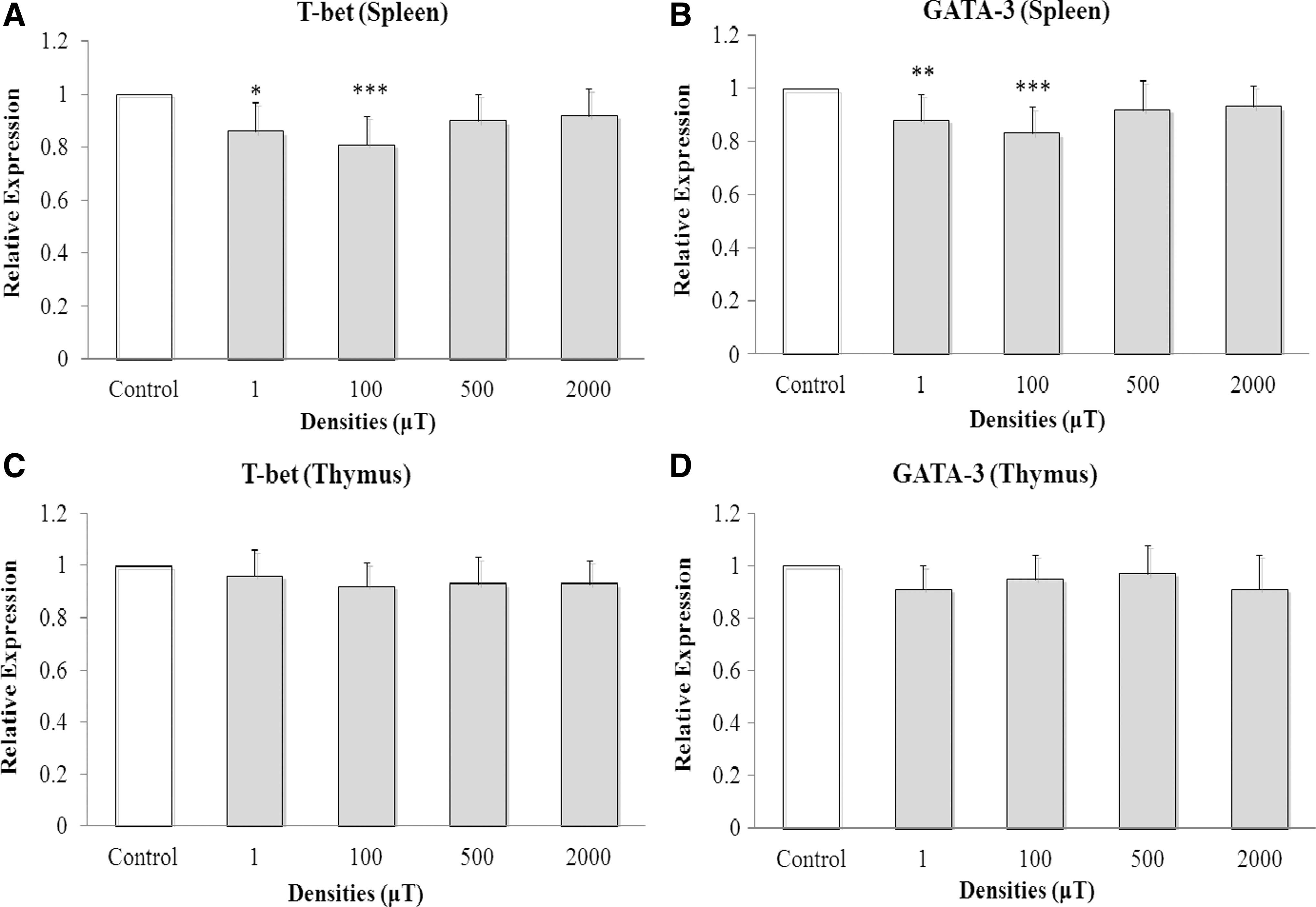
Relative expressions of T-bet and GATA-3 mRNA as a result of daily ELF-EMF exposure for 2 months. Relative expression is shown. Results are presented as mean ± standard deviation. All data are from rats that received human serum albumin injections over the course of a 4-week period before the end of the 60-day ELF-EMP exposures. One-way analysis of variance performed to compare relative expressions of T-bet and GATA-3 mRNA among the groups. A Tukey test was used to analyze splenic T-bet and GATA-3 mRNA expression.
Data for serum IFN-γ and IL-4 showed that levels were only altered at “prestimulation” (ie, before beginning HSA injection regimen; P < 0.001 versus control rat values for each cytokine) (Table 2). The serum levels of IFN-γ and IL-4 in control rats were 100.95 [±7.7] pg/mL and 44.72 [±4.6] pg/mL, respectively; these were reduced to 83.15 [±8.2] pg/mL and 29.21 [±3.1] pg/mL, respectively, due to host exposure to 100 μT ELF-EMF. This effect was lost as a result of time and/or HSA treatments; there were no significant differences in control values for IFN-γ and IL-4 following stimulation, regardless of the flux density used.
Effects of Various Extremely Low Frequency Electromagnetic Field Magnetic Flux Densities on Serum Levels of Interferon-γ and Interleukin-4
Results shown are mean ± standard deviation (pg/mL). Values of IFN-γ and IL-4 are from the groups at pre- and poststimulation of the immune system (multiple injections of human serum albumin over a 4-week period before end of ELF-EMF exposure). One-way analysis of variance and Tukey tests each showed that serum IFN-γ and IL-4 levels in rats exposed to 100 μT density were significantly altered at the prestimulation phase, that is, 30 days after start of daily ELF-EMF exposure. No significant differences were found among any of the test groups after the poststimulation period.
P < 0.001 compared with controls.
ELF-EMF, extremely low frequency electromagnetic fields; IFN-γ, interferon-γ; IL-4, interleukin-4.
In Figure 2, an attempt to model changes in magnetic flux densities with splenic T-bet and GATA-3 mRNA expression in the HSA-stimulated rats and serum IFN-γ and IL-4 levels in the prestimulation phase is provided. It seems that the effect of various magnetic flux densities on splenic T-bet and GATA-3 mRNA levels and serum IFN-γ and IL-4 levels followed parabolic curve models. This meant that with increases in magnetic flux density >100 μT, the effect on the measured factors would be expected to decrease.
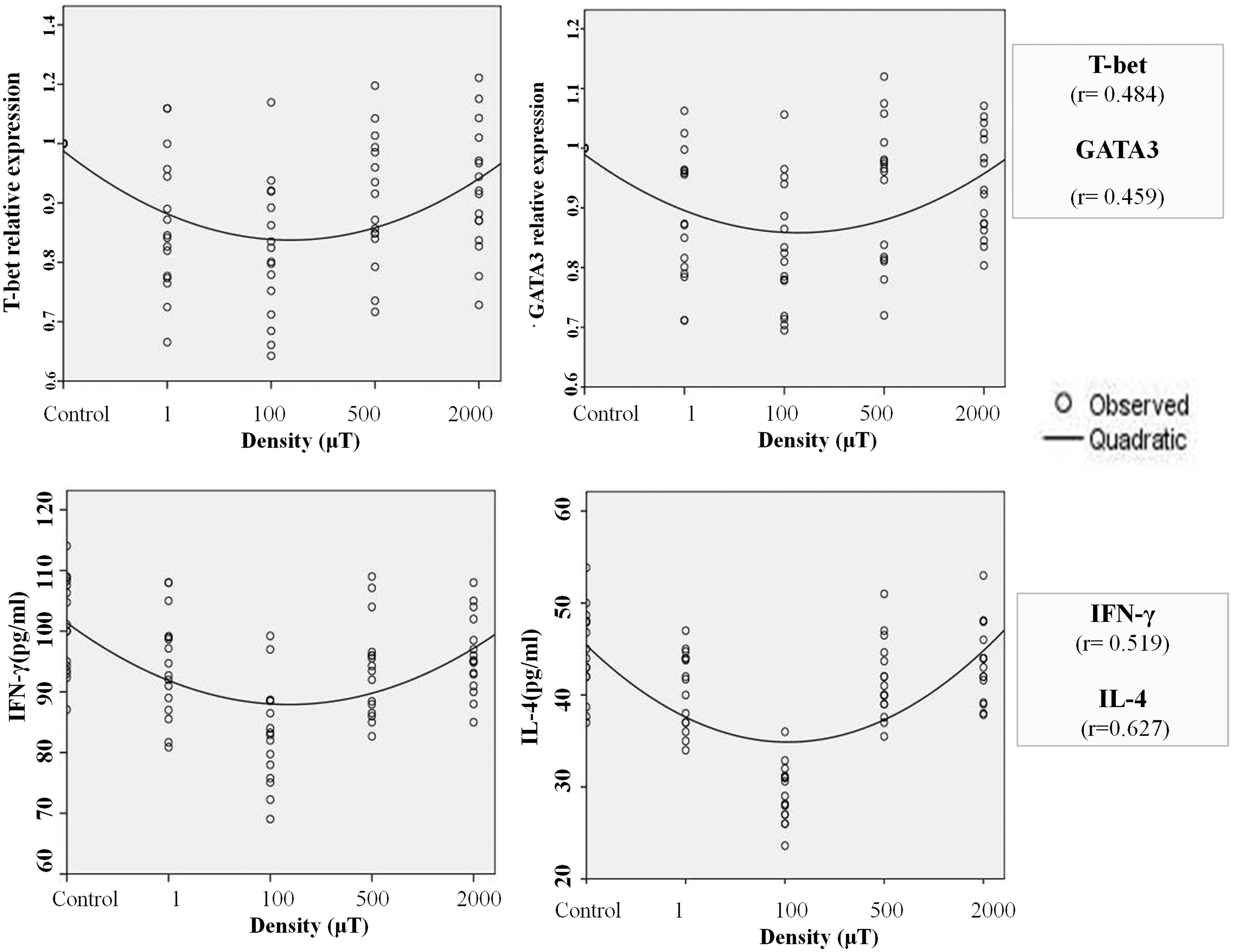
Curve estimation models (quadratic regressions) for T-bet, GATA-3, IFN-γ, and IL-4 endpoints. Modeling revealed effects of various flux magnetic densities (1, 100, 500, and 2000 μT) on splenic T-bet and GATA-3 mRNA expression and serum IFN-γ and IL-4 levels (during prestimulation phase), followed by a parabolic curve model. At lower densities of ELF-EMF, the effect was higher than that observed at higher densities.
Discussion
The results of the present study showed that exposure to ELF-EMF reduced expression levels of T-bet and GATA-3 genes in the spleen of the HSA-stimulated rats, but there was no significant effect on levels in the thymus. The data also indicated that serum levels of IFN-γ and IL-4 were decreased in a resting/unstimulated immune system, but those unaffected had exposure that occurred following host immune system stimulation with HSA.
Reductions in splenic expression of T-bet and GATA-3 mRNA were only observed with use of magnetic flux densities of 1 and 100 μT. The higher densities had no significant effect on expression. This is consistent with our previous reports about the effects of ELF-EMF on reduction of the transcription factor forkhead box P3 (Foxp3) expression only at 1 and 100 μT (Mahdavinejad and others 2018). A study of humans showed also a significant reduction in natural killer cell activity due to exposure to 1 μT ELF-EMF (Gobba and others 2009). In another study, it was also shown that expression levels of T-bet and GATA-3 mRNA were not altered in the spleens of exposed mice following host exposures to magnetic flux densities of 500 and 1000 μT (Luo and others 2016).
T-bet belongs to the family of T-box transcription factors. In the presence of antigen, IL-12, and IFN-γ, the gene of T-bet is induced and expressed in naive CD4+ T cells. This is the reason why in this study the rats were injected with HSA. The HSA was used to activate the resting naive CD4+ T cells and to shift them toward formation of possible functional patterns of Th1, Th2, Th17, and Treg cell subsets. The expression of T-bet in turn helps in the production of IFN-γ by the Th1 cells. Therefore, a positive amplification loop causes differentiation of more naive CD4+ T cells toward the Th1 subset. On the contrary, due to antigen activation and IL-4 production, GATA-3 gene expresses and activates the expression of IL-4, IL-5, and IL-13 genes and causes the production of these cytokines that drive the naive CD4+ T cells to Th2 subset. Like T-bet, production of IL-4 upregulates the expression of GATA-3 (positive amplification loop). The main subject that needs to be investigated in the future is that whether due to ELF-EMF, the decrease of IFN-γ and IL-4 caused the downregulation of the expression of T-bet and GATA-3 or vice versa (Hoyler and others 2013; Tindemans and others 2014; Raphael and others 2015).
The current study investigated the expression levels of T-bet and GATA-3 mRNA in the thymus as well. The results indicated no variation in thymic expression levels of these genes due to exposure to the various ELF-EMF densities. This finding may be related to higher expressions of T-bet and GATA-3 in the thymus compared with the spleen. By this, there could be a greater reservoir of mRNA, and so, any subtle effects from the EMF could be masked in the thymus. This difference in outcomes could also potentially be related to the distinctly differing types of cells that are present in the thymus and spleen (Kurata and others 2002) or to the overall architecture of these organs. Whether their respective locations in the body or the presence of any structural elements in each organ might impact on radiation penetrance, total dosage, and so on remains to be determined.
Serum IFN-γ and IL-4 levels were only significantly reduced by the magnetic flux density of 100 μT in a prestimulated immune system. These results contrast with those from a previous study showing that serum IFN-γ and IL-4 levels were not changed at 100 μT (Salehi and others 2013). In this case, the only variable that differed between these two studies was duration of exposure. In the 2013 study, the exposure duration was 3 months, whereas in the present study (at prestimulation phase) it was only 1 month. Thus, it seems that duration of exposure to ELF-EMF is an important factor in subsequent effects on the immune system.
An example of short-term in comparison with long-term effects from ELF-EMF can be inferred from research by Luo and others (2016). Those investigators measured the effect of 50 Hz EMF (with 500 μT flux density) on murine body weight change every 10 days over a period of 60 days of exposures to the EMF. The data showed that the body weights of the exposed mice were only significantly decreased on days 20 and 30. In a study by Kleijn and others (2016), effects of ELF-EMF with a magnetic flux density of 10 μT were assessed on leukocyte counts in mice after 1-week (short-term) and 15-week (long-term) exposures. Those studies showed that with the short-term but not long-term exposures, leukocyte counts in the hosts were significantly increased. Pro-opiomelanocortin (POMC) gene expression in the pituitary gland and plasma level of adrenocorticotropic hormone were also found to have declined in the short-term-exposed hosts (compared with control counterparts). It would seem that in the short-term exposure experiments, ELF-EMF caused an activation of the hypothalamic/pituitary/adrenal axis. However, with a more prolonged (long-term) exposure, there is no such activation (effect has waned) (Kleijn and others 2016).
Melatonin fluctuations might also be a means by which the ELF-EMF might have impacted the host immune cells. Besides the pineal gland, melatonin can be produced by leukocytes and is a modulation of immune system responses. Obayashi and others (2015) noted that increased melatonin secretion was associated with lower circulating leukocyte and platelet counts in a general elderly population (HEIJO-KYO cohort). How this relates to ELF-EMF can be seen in the study by Selmaoui and Touitou (1995), in which Wistar rats exposed to a 100 μT density of ELF-EMF for 30 days (short term) displayed significantly decreased serum melatonin levels. In contrast, Seifpanahi-Shabani and others (2016) saw that when Wistar rats were treated with a 100 μT density of ELF-EMF for 90 days (long term), serum melatonin levels were not changed relative to control host values. This ties back well to the Kleijn and others (2016) findings, that is, 1-week (short-term) exposure to ELF-EMF resulted in significant increases in circulating leukocyte counts in hosts (with lower melatonin levels). Concurrently, the loss of “increases” in melatonin in hosts that underwent prolonged exposure could then mean there was no up/downward pressure to affect levels of circulating leukocytes in these hosts (Kleijn and others 2016).
Finally, we want to note that the effects of various magnetic flux densities of ELF-EMF on T-bet and GATA-3 mRNA levels and serum IFN-γ and IL-4 could be modeled to a parabolic curve. The ELF-EMF caused a decrease in these factors at lower densities of 1 and 100 μT, but at higher densities (500 and 2000 μT) there were no longer any effects on these measured parameters. It would seem that the ELF-EMF suppressed some aspects of the immune system at lower density, but such effects were either not induced or mitigated (potentially by activation of some as-yet undefined protective factors). The latter is a concept that has been promulgated by Eroz and others (2016) and Saritas and others (2016).
In conclusion, it appears that a 60-day exposure to ELF-EMF (daily) induced a reduction in expression levels of splenic T-bet and GATA-3 mRNA in HSA-stimulated rats. It is not known if these same effects were manifested in unstimulated rats or if the same effects occurred only after 30 days of daily EMF exposures. Furthermore, the EMF had no significant effect on thymic T-bet and GATA-3 mRNA levels in HSA-stimulated rats. The same EMF exposures also resulted in decreases in serum levels of IL-4 and IFN-γ, but only before HSA stimulation of the hosts. Therefore, it seems that some functions of both Th1 and Th2 cells were compromised due to ELF-EMF exposures at lower densities, but the effect is transient and apparently might be related to the immune activation status of a given host.
Footnotes
Acknowledgment
This project was financially supported by the Vice Chancellor for Research and Technology, Hamadan University of Medical Sciences (Grant No. 9210173428).
Author Disclosure Statement
The authors declare no conflicts of interest. The authors alone are responsible for the content of this article.