
Abstract
Chronic infection with the hepatitis C virus (HCV) is a major cause of cirrhosis and hepatocellular carcinoma. In 2009, genome-wide association studies (GWAS) strongly linked genetic variants in the interferon lambda (IFN-λ) chromosomal region to HCV clearance. In 2013, discovery of the IFNL4 gene provided a functional explanation for those GWAS findings. The IFNL4-ΔG/TT (rs368234815) variant controls generation of the IFN-λ4 protein. Paradoxically, the IFNL4-TT allele, which abrogates IFN-λ4, associates with higher rates of spontaneous HCV clearance and better response to treatments for HCV infection. The finding that a “knock-out” allele for IFN-λ4 enhances HCV clearance challenges the paradigm of IFNs as antiviral cytokines. Genetic variants in the IFN-λ region have also been associated with hepatic inflammation and fibrosis from various etiologies, however, alleles that are linked with improved HCV clearance associates with worse inflammation and fibrosis. These studies demonstrate that GWAS of infectious diseases may yield important and unexpected biological insights.
Hepatitis C Virus Infection
Chronic infection with hepatitis C virus (HCV) affects ∼71 million people worldwide, or 1% of the global population (Polaris Observatory HCV Collaborators 2017). In the United States, at least 3.5 million people have chronic hepatitis C (CHC) (Edlin and others 2015). HCV is primarily transmitted through exposure to infected blood or blood products via injection drug use, unscreened blood transfusions or iatrogenic transmission (World Health Organization 2017). Approximately 70%–80% of individuals with an acute HCV infection are unable to clear the virus spontaneously and develop CHC. Among individuals with CHC, 10%–20% will develop advanced liver disease of whom, 1%–5% will go on to develop hepatocellular carcinoma (HCC) (Lavanchy 2011). Globally, CHC is responsible for 25% of cirrhosis and HCC cases (Perz and others 2006); in developed countries CHC is a leading indication for liver transplantation (Brown 2005).
CHC leads to liver injury through hepatocyte apoptosis and necrosis, driven by sustained inflammation and activation of immune mechanisms. Liver damage seen in HCV infection is likely caused by the host's immune response rather than the virus itself (Liang and others 2000). Prolonged inflammation of the liver can lead to the accumulation of excess connective tissue, resulting in fibrosis (Guidotti and Chisari 2006; Arzumanyan, Reis, and Feitelson 2013). Cirrhosis, the most advanced stage of fibrosis, is characterized by scarring of the liver and alteration of the normal liver structure into abnormal nodules (Anthony and others 1978). Inflammation, accompanied by oxidative stress and cellular proliferation, provokes somatic mutations and carcinogenesis. In patients with CHC, the risk of HCC is highly correlated with the severity of liver fibrosis (Forner and others 2012; Arzumanyan, Reis, and Feitelson 2013).
Previously, treatment with pegylated interferon alpha combined with the nucleoside inhibitor ribavirin (peg-IFNα/ribavirin) for 48 weeks was the standard of care for HCV treatment (Scheel and others 2013). Successful treatment cures HCV infection by producing a sustained viral response (SVR), defined as an undetectable level of HCV RNA in serum 12–24 weeks after the end of treatment. Treatment efficacy with the peg-IFNα/ribavirin regimen is modest and varies with the viral genotype (VGT) (McHutchison and others 1998). HCV strains are classified into 7 genotypes; VGT-1 and VGT-3 are the predominant genotypes globally and VGT-1 is the most prevalent strain in the United States (Messina and others 2015). Only 40%–50% of patients infected with VGT-1 and ∼70% infected with VGT-3 achieve SVR after peg-IFNα/ribavirin therapy (Fried and others 2002; Shiffman and others 2007).
In addition to viral factors, certain demographic and clinical factors are associated with better response to peg-IFNα/ribavirin (Reddy and others 1999). Race associates strongly with achieving SVR after peg-IFNα/ribavirin therapy and African American patients are less likely than white patients to have an SVR even after other factors are considered (Reddy and others 1999; Kinzie and others 2001; Layden-Almer and others 2003; Muir and others 2004; Conjeevaram and others 2006; Wilder and others 2016). Those findings supported a hypothesis that genetic factors play a role in response to treatment for CHC. Research efforts to address that question resulted in important discoveries with relevance to HCV infection and beyond.
IFN-λ Region Genetic Variants Strongly Associate with HCV Treatment Response and Spontaneous Clearance
Between 2009 and 2010, 4 groups of investigators independently reported results of genome-wide association studies (GWAS) that linked genetic variants in the IFN-λ chromosomal region with response to peg-IFNα/ribavirin therapy for CHC (Ge and others 2009; Suppiah and others 2009; Tanaka and others 2009; Rauch and others 2010). In a GWAS, individuals are tested for hundreds of thousands of single nucleotide polymorphisms (SNPs) that have been selected to cover all regions of the human genome with the goal of identifying regions that harbor functional genetic variants (Feero and others 2010). Ge and others (2009) found rs12979860 to be the SNP most significantly associated with SVR. Whereas most GWAS of noninfectious outcomes have yielded associations of modest effect size (Feero and others 2010), individuals with the rs12979860-CC genotype had a 2-fold greater rate of SVR than those with the rs12979860-CT or -TT genotypes. In addition, Ge and others (2009) reported that African American participants were less likely to have the favorable rs12979860-CC genotype, which offered an explanation for previous observations of poorer treatment response in that group. Genotype for rs12979860 also associated with the level of HCV RNA before treatment, but in a surprising fashion. Higher HCV RNA is generally associated with poorer treatment response (Ge and others 2009), however, the rs12979860-CC genotype associated with both higher pretreatment viral levels and a higher rate of SVR (Ge and others 2009).
In reports published almost simultaneously with that of Ge and others, 2 other groups reported consistent findings. The rs12979860 was not included on the array used by these investigators and their findings focused on a nearby SNP (rs8099917). Among individuals of European ancestry who were treated with peg-IFNα/ribavirin, Suppiah and others (2009) observed that the rs8099917 SNP was strongly related with a 2-fold association with SVR. Those with at least 1 copy of the rs8099917-G allele had higher SVR rates compared with noncarriers. The strongest associations were seen in homozygotes for the unfavorable genotype (rs8099917-TT) compared to individuals who carry the rs8099917-G allele (Suppiah and others 2009). In a Japanese population, Tanaka and others (2009) also found that individuals who were homozygous for the rs8099917-T allele had poorer response to IFNα-based therapy and were less likely to clear the virus than those who carry at least one copy of the rs8099917-G allele. The strong associations reported in these landmark studies suggested that the identified SNPs might have clinical utility for predicting response to treatment of CHC and established the IFN-λ region as playing a major role in HCV clearance.
Soon thereafter, studies linked these SNPs with spontaneous clearance of HCV. Thomas and others (2009) observed that rs12979860-CC genotype associated with spontaneous clearance in a manner similar to the findings for treatment response; homozygotes for the rs12979860-C allele were 3 times more likely to have cleared HCV as those who carried the rs12979860-T allele. Those investigators also were the first to provide a global map of the distribution of the rs12979860 variant, demonstrating dramatic differences in allele frequencies among African, European, and Asian populations. The favorable rs12979860-C is the major allele in Asians and Europeans, but the minor allele in Africans (Thomas and others 2009). In a Swiss cohort, Rauch and others (2010) showed that genotype for rs8099917 associated with both spontaneous HCV clearance and response to treatment with peg-IFNα/ribavirin.
At the time of those groundbreaking findings regarding HCV infection, the discovery of the IFN-λ region itself was still relatively recent. In 2003, 2 independent groups of investigators reported the discovery of 3 novel genes with similarity to type I IFNs, although with signaling via a receptor with expression that was much more limited than those for type I IFNs (Kotenko and others 2003; Sheppard and others 2003). Because IFN-λ was thought to be largely redundant to type I IFNs, the connection between genetic variation in the IFN-λ region and clearance of HCV infection was not obvious.
Discovery of IFNL4 Explains Association of IFN-λ Variants with HCV Clearance
Once a relevant chromosomal region is identified based on associations with SNPs in a GWAS, additional studies are needed to attempt to identify the functional genetic variant underlying that signal (Collins and others 1997; Feero and others 2010). Knowledge of linkage disequilibrium (LD), the nonrandom correlation between alleles at different loci (Slatkin 2008), is used to identify possible functional variants. It is noteworthy that although GWAS have implicated specific genetic regions for a wide range of disease phenotypes, in most cases a functional variant is yet to be identified (Feero and others 2010). The discovery of IFNL4 provides an exception to the common situation.
IFNL4 and the IFN-λ4 protein were discovered by our collaborative group during a search for that functional genetic variant (Prokunina-Olsson and others 2013). In discovering IFNL4, we showed that the rs12979860 SNP lies within intron 1 of IFNL4 and that rs8099917 lies nearer IFNL4 than IFNL3 (Fig. 1) (Prokunina-Olsson and others 2013). IL28B was the official gene symbol for IFNL3 at the time of the 2009 HCV GWAS articles (the current nomenclature was adapted upon the discovery of IFNL4 in 2013) and confusion on the location of these GWAS markers continues with these SNPs still frequently referred to as IFNL3 or “IL28B” variants. Unfortunately, this problem extends beyond semantics. There is a common misunderstanding that variation in IFNL3 underlies genetic associations with HCV clearance.

SNPs associated with response to peg-IFNα/ribavirin therapy and spontaneous clearance of HCV in GWAS (Ge and others 2009; Suppiah and others 2009; Tanaka and others 2009). rs12979860 lies within IFNL4 and rs8099917 lies nearer IFNL4 than IFNL3, therefore, these SNPs are properly termed variants of IFNL4, which was discovered subsequent to the GWAS reports (Prokunina-Olsson and others 2013). GWAS, genome-wide association studies; HCV, hepatitis C virus; IFN, interferon; SNP, single nucleotide polymorphism.
The rs12979860 and rs8099917 SNPs are in LD with a common dinucleotide frameshift variant that we call IFNL4-ΔG/TT (rs368234815; Fig. 2). This variant controls IFNL4, with the IFNL4-ΔG allele creating an open reading frame for full-length IFN-λ4 protein and the alternative allele (IFNL4-TT) abrogating the protein (Prokunina-Olsson and others 2013). Thus, IFNL4-ΔG is a functional variant that controls the generation of IFN-λ4 and IFN-λ4 is produced only by those with at least one copy of the ancestral IFNL4-ΔG allele.
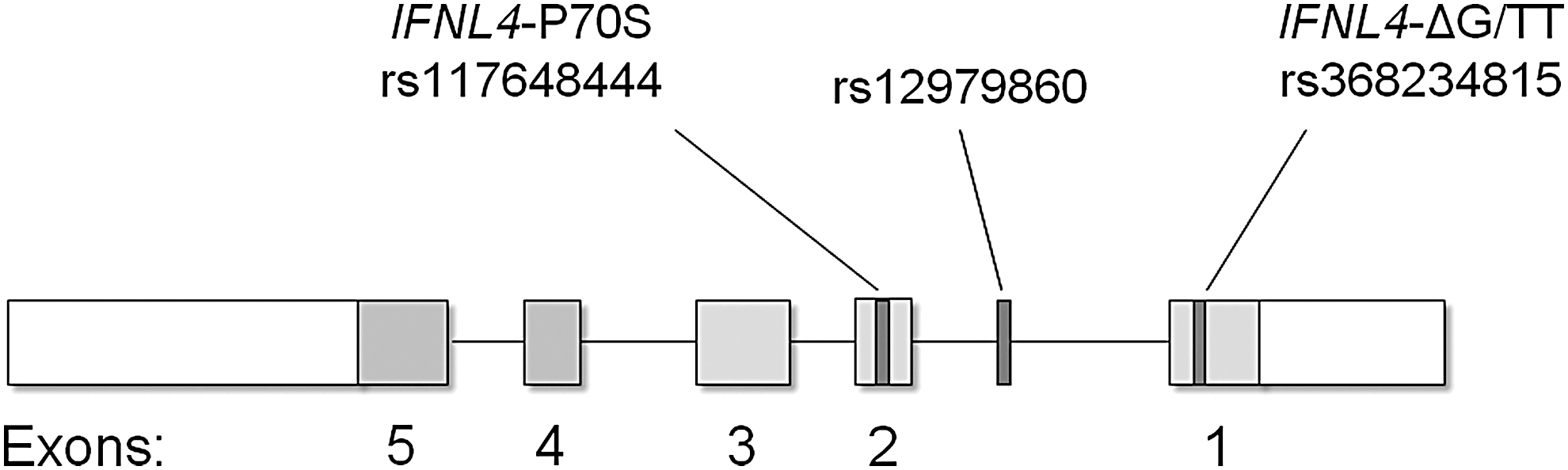
IFN-λ4 structure and genetic variants. The IFNL4-ΔG/TT variant (rs368234815), which controls generation of IFN-λ4, lies in exon 1; GWAS marker rs12979860 lies in intron 1; the IFNL4-P70S variant (rs117648444), which modifies IFN-λ4, lies in exon 2 (Prokunina-Olsson and others 2013).
Genotype for the IFNL4-ΔG/TT variant explains associations between IFN-λ variants and HCV clearance that were observed in GWAS. The IFNL4-ΔG allele is in high LD with the rs12979860-T allele in Asians (r 2 = 1.00), correlates to a lesser extent in Europeans (r 2 = 0.91), and has the weakest LD in Africans (r 2 = 0.71) (Prokunina-Olsson and others 2013). To compare the relative strength of associations for IFNL4-ΔG/TT variant and the rs12979860 GWAS marker, we conducted analyses among African Americans, in whom LD between these variants is weaker than those of European ancestry. Examining decline in HCV RNA levels through the first 28 days of treatment with peg-IFNα/ribavirin among African American participants in the Virahep-C Trial, genotype for the IFNL4-ΔG/TT variant was a stronger predictor than IFNL4-rs12979860 genotype (P = 0.015; Fig. 3) and the IFNL4-ΔG allele, which generates the IFN-λ4 protein, was linked to worse response (Prokunina-Olsson and others 2013). We found consistent results in analyses for spontaneous HCV clearance among African American individuals (Prokunina-Olsson and others 2013; Aka and others 2014). Other investigators demonstrated that treatment outcomes were more strongly associated with IFNL4-ΔG/TT genotype than rs12979860 genotype in Europeans (Bibert and others 2013; Franco and others 2014). It seems paradoxical that expression of an additional IFN-λ interferon interferes with viral clearance (O'Brien and others 2014). Other articles in this special issue address this issue (Prokunina-Olsson and others, JICR submitted?).
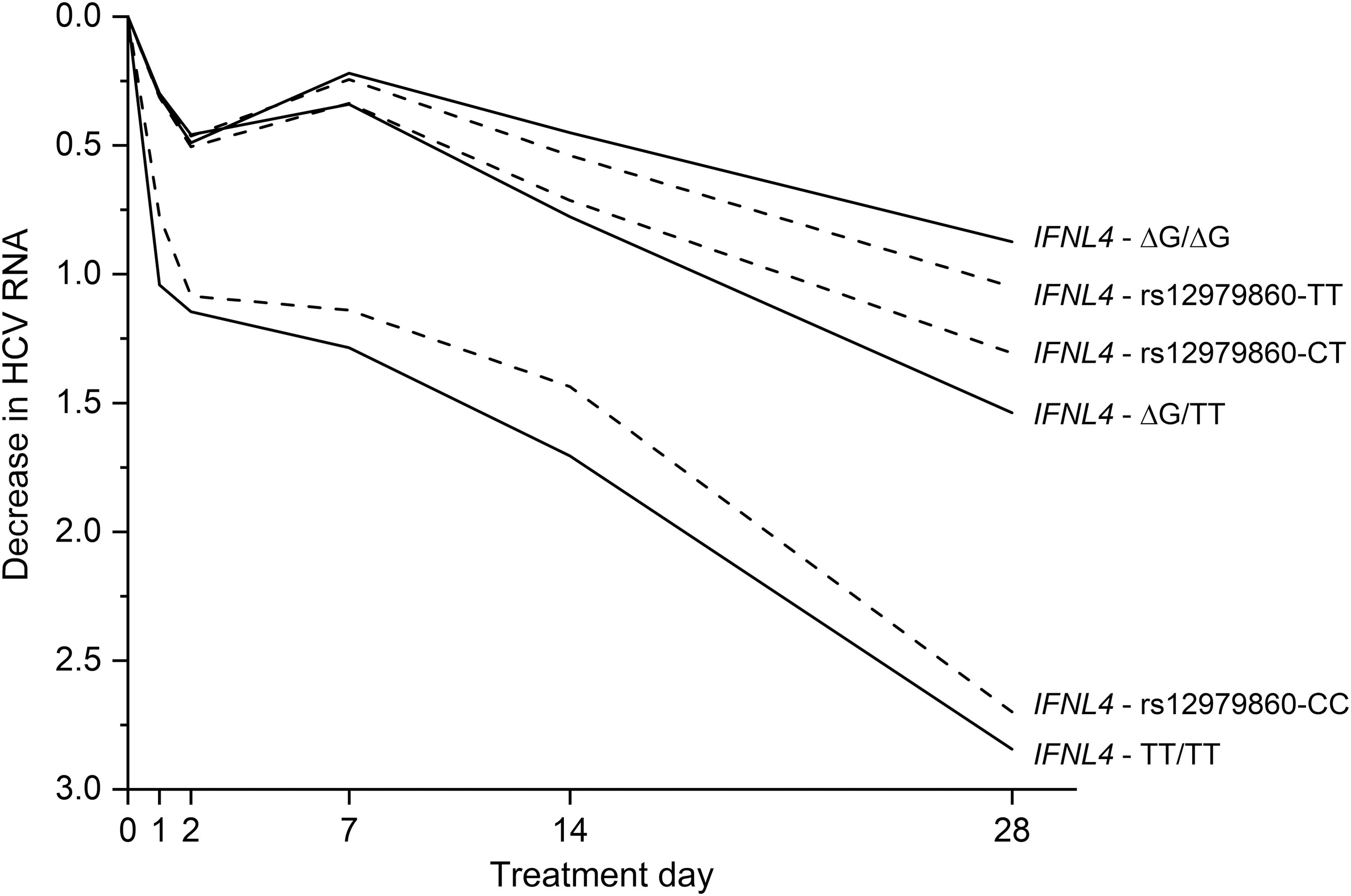
Decline in HCV RNA levels in response to treatment with peg-IFNα/ribavirin among African American participants in the Virahep-C Trial. Genotype for IFNL4-ΔG/TT was a stronger predictor than that for GWAS marker IFNL4-rs12979860 (P = 0.015) (Prokunina-Olsson and others 2013).
Nonsynonymous variant rs117648444 within exon 2 of IFNL4 is a second functional polymorphism that affects both the IFN-λ4 protein and HCV clearance (Fig. 3). This mutation alters amino acid position 70 of IFN-λ4, resulting in the substitution of serine (S70) for proline (P70). Compared to IFN-λ4 P70, the derived IFN-λ4 S70 form produces lower expression of intrahepatic IFN stimulating genes and diminished antiviral activity in vitro, yet individuals with a genotype that yields this weakened form of IFN-λ4 have higher rates of spontaneous HCV clearance and response to peg-IFN-α/ribavirin treatment than those with the P70 variant (Galmozzi and Aghemo 2014; Terczyńska-Dyla and others 2014), which is consistent with the overall finding that generation of IFN-λ4 impairs HCV clearance.
Studies of the protein altering IFNL4-ΔG/TT and IFNL4 P70S polymorphisms firmly establish IFNL4 as the primary gene involved in HCV clearance, however, a functional polymorphism found in the 3′ untranslated region of IFNL3 is also of interest. The rs4803217 SNP affects the degradation of IFNL3 mRNA and binding of HCV-induced microRNA (McFarland and others 2013). Similar to the relationships between the IFNL4-ΔG/TT and IFNL4-rs12979860 variants, LD between the IFNL3 rs4803217-T allele, which enhances IFNL3 mRNA degradation, and the IFNL4-ΔG allele is complete in Asians (r 2 = 1.00) and strong in Europeans (r 2 = 0.88), but weaker in Africans (r 2 = 0.63) (Prokunina-Olsson and others 2013). Again, strong LD makes it difficult to differentiate a possible effect of IFNL3 rs4803217 from that of IFNL4-ΔG/TT in Asians or Europeans. To overcome that issue, we compared associations for closely linked genotypes in African Americans. We found that IFNL4-ΔG/TT was more strongly associated with both response to peg-IFNα/ribavirin therapy and spontaneous HCV clearance than rs4803217 (O'Brien and others 2015). Further, those with the IFNL4-ΔG:rs4803217-G haplotype, a combination that combines the unfavorable IFNL4 allele with the putatively favorable IFNL3 allele, was associated with the lowest SVR rates (O'Brien and others 2015). These results indicate that expression of the IFN-λ4 protein is the primary driver of HCV clearance. The possible contribution of IFNL3 rs4803217 to HCV clearance deserves further examination.
IFNL4 Genotypes Associate with Response to New Therapies for Hepatitis C
Direct-acting antiviral agents (DAAs) that attack viral replication and assembly were introduced in 2011(Scheel and others 2013) and, by 2014, highly effective regimens based on 2 or more second generation DAAs became available. These therapies, which target the HCV NS3/4A protease, NS5B RNA polymerase and NS5A viral proteins, produce cure rates >90% (Messina and others 2015) with fewer adverse effects than IFN-based therapies (Scheel and others 2013; Baumert and others 2019). DAAs have revolutionized the treatment of HCV and led to ambitious goals for reducing the prevalence of CHC and its associated morbidity and mortality worldwide.
Sofosbuvir, which inhibits the HCV NS5B protein, forms the backbone of several DAA regimens. Sofosbuvir can be combined with ribavirin for HCV treatment and Meissner and others (2014) showed that IFNL4-ΔG associated with slower viral decay and decreased drug efficacy in patients with HCV genotype 1 who were treated with this regimen. Sofosbuvir is more effective when combined with other DAAs. Both rs12979860-CC and rs8099917-TT were associated with higher SVR after 12 weeks treatment with a regimen that combined sofosbuvir with daclatasvir, which inhibits the HCV NS5A protein (Khan and others 2019). An assessment of treatment response to a variety of DAA regimens among HCV-1 infected patients treated in the Veterans Health Administration found that those with the IFNL4-ΔG/ΔG genotype had a much lower cure rate than either IFNL4-ΔG/TT or IFNL4-TT/TT genotypes (Backus and others 2018).
Standard treatment duration for sofosbuvir-based regimens is 12 weeks, however, a shorter course may be highly effective in patients with a favorable IFNL4 genotype. The ION-3 trial evaluated the combination of the NS5A inhibitor ledipasvir with sofosbuvir (Kowdley and others 2014). In reanalyzing data from that study, we found that only 2% of patients with the IFNL4-rs12979860-CC genotype failed to respond to treatment at 8 weeks compared to higher rates of failure among carriers of the unfavorable IFNL4-rs12979860-T (O'Brien and others 2017a). Similarly, we observed a 1% relapse rate for patients with the IFNL4-rs12979860-CC genotype who received 8 weeks of a regimen that combined sofosbuvir with the NS5A inhibitor velpatasvir and voxilaprevir, which inhibits HCV NS3/4A protease (O'Brien and others 2017b). These results demonstrate the potential utility of IFNL4 genotyping to inform decisions concerning the appropriate duration of therapy with sofosbuvir regimens. Shorter duration treatments could address the high cost of DAA regimens that is a barrier to fuller implementation of these curative treatments. Decisions regarding the appropriate duration of DAA treatment might be informed through an algorithm based on IFNL4 genotype and other relevant factors (O'Brien and others 2017a). Given patterns observed for spontaneous clearance and response to peg-IFN-α/ribavirin, an analysis of response to DAA treatment that considered both IFNL4-ΔG/TT and IFNL4 P70S might provide the best clinical prediction. Future studies should address that question. Alternatively, in regions such as East Asia where the frequency of the IFNL4-TT/TT genotype is very high, it might be cost saving to treat all patients for a shorter duration. Thus, knowledge regarding genotype for the IFNL4-ΔG/TT variant at either the individual or population level, might improve cost-effectiveness of treating the 71 million who are infected with HCV.
IFN-λ Region Variants Associate with Hepatic Inflammation and Fibrosis
Evidence that variants in the IFN-λ region associated with HCV clearance led to interest in whether these polymorphisms might also play a role in the progression of CHC. Necroinflammatory grade is thought to be the best predictor of fibrosis progression, which can lead to cirrhosis (Goodman 2007). In 2012, Bochud and others reported results from a study of ∼2,000 European patients with CHC. They found that the rs8099917-G allele, which is linked with impaired HCV clearance and was subsequently shown to be in LD with the IFNL4-ΔG allele, associated with decreased necroinflammatory activity, fibrosis, and fibrosis progression (Bochud and others 2012). Also among patients with CHC, Noureddin and others found rs12979860-CC genotype was associated with significantly higher portal inflammation and alanine aminotransferase levels. However, in a longitudinal analysis of paired biopsy results, genotype for rs12979860 was not associated with the frequency of fibrosis progression in that cohort (Noureddin and others 2013). In a subsequent study, Eslam and others examined the relationship between the IFNL4 rs12979860 marker and hepatic inflammation and fibrosis progression among 4,000 European-ancestry patients. Consistent with Bochud and others, they demonstrated that the rs12979860-CC genotype, which is associated with increased HCV clearance, associated with increased inflammation and fibrosis in CHC patients. Furthermore, they showed that this relationship extended to individuals with chronic hepatitis B and nonalcoholic fatty liver disease (NAFLD) (Eslam and others 2015). In 2017, Petta and others (2017) reported that the functional IFNL4 rs368234815-TT allele, which abrogates IFN-λ4, is associated with more severe fibrosis in NAFLD patients.
Taken together, those studies suggested that IFN-λ4 protects against hepatic inflammation and fibrosis of both infectious and noninfectious etiologies. However, results from a second article by Eslam and others challenged that hypothesis. Among ∼2000 patients of European ancestry with CHC, they examined associations of genotype for the IFNL4-ΔG/TT, IFNL4 rs12979860, and IFNL3 rs4803217 variants with hepatic inflammation, hepatic fibrosis, and inflammatory cell counts in liver biopsy specimens (Eslam and others 2017). Given the high LD between these genetic variants in that study population, they were unable to determine which polymorphisms had the strongest associations with these outcomes, however, they found no difference in inflammation or fibrosis between the variants that create the IFN-λ4 P70 and IFN-λ4 S70 proteins (Eslam and others 2017), a finding that was inconsistent with previous observations for HCV clearance (Galmozzi and Aghemo 2014; Terczyńska-Dyla and others 2014). Eslam and others also found that quantitative IFNL3 mRNA from liver biopsies of patients with CHC associated with IFNL4 rs12979860 genotype, hepatic inflammation, and hepatic fibrosis. On that basis, they concluded that IFN-λ3, rather than IFN-λ4, was the likely mediator of genetic associations with hepatic inflammation and fibrosis.
Because of the strong LD between IFNL3 and IFNL4 genetic variants in European populations, it remains an open question as to which genetic variant most strongly associates with hepatic inflammation/fibrosis (Park and others 2018). It is possible that the presence or absence of IFN-λ4 plays a role in hepatic inflammation, but the IFN-λ4 P70S does not affect that outcome. Future studies that include populations of African ancestry, in which LD between INF-λ region variants is less than that in Europeans, might help resolve this question. Better understanding of the mechanisms underlying the relationship between INF-λs and fibrosis progression could lead to interventions to prevent cirrhosis. Further, genetic factors could be used to improve risk stratification models for fibrosis progression (Tamaki and others 2015; Eslam and others 2016).
Future Directions—IFN-λ Variants in Infections and Other Conditions
Several lines of evidence suggest genetic variants in the IFN-λ region could play a role in a wider range of infections and other conditions. There was strong selection for the IFNL4-TT allele (which abrogates IFN-λ4), such that the IFNL4-ΔG/TT polymorphism is among the top 0.5% for differences between African and Asian populations differences genome-wide (Key and others 2014). The mutation that inactivated IFN-λ4 expression developed before the out-of-Africa migration and the ancestral IFNL4-ΔG allele is the most common form among Africans, however, subsequent selection resulted in IFNL4-TT becoming the major allele among Europeans and Asians (Prokunina-Olsson and others 2013), with allele frequencies for IFNL4-TT exceeding 90% in East Asian populations. HCV infection is a bloodborne infection transmitted primarily through use or misuse of medical devices, therefore, it was probably not a common infection before the modern era. Given the long period between initial infection and development of medical conditions that would impair reproduction, it is unlikely that HCV infection is the infectious driver behind the selective pressure of IFNL4 mutation. Therefore, loss of IFN-λ4 production may be advantageous in the context some of other infectious or inflammatory conditions (O'Brien and others 2014).
Recent studies in mice demonstrate that IFN-λs protect tissue barriers against a wide range of viral infections. Lazear and others (2015) demonstrated that exogenous IFN-λ3 tightens endothelial junctions of the blood brain barrier in response to West Nile Virus infection, thereby, reducing viral neuroinvasion. Mice lacking the IFN-λ receptor exhibit increased susceptibility to viral pathogens including norovirus (Nice and others 2015), rotavirus (Hernández and others 2015), influenza (Crotta and others 2013; Klinkhammer and others 2018), severe acute respiratory syndrome (Mahlakõiv and others 2012), and herpes simplex virus-2 (Ank and others 2006). Treatment of influenza-infected mice with IFN- λ therapy reduces viral titers and shedding (Crotta and others 2013; Galani and others 2017). Among human volunteers challenged with influenza (Egli and others 2014) or rhinovirus (Contoli and others 2006), reduced viral replication correlated with increased IFN-λ production. In contrast to these apparent benefits, IFN-λ increased bacterial levels of Staphylococcus and Pseudomonas in mice (Cohen and Prince 2013). Understanding the role of IFN-λ in these various conditions has important implications for potential treatment modalities.
In humans, genotype for the IFNL4-ΔG/TT variant has been associated with opportunistic infections in immunocompromised patients and certain cancers. Cytomegalovirus (CMV) retinitis causes visual impairment and blindness. Bibert and others (2014) found a 2-fold increase in the risk of developing CMV-related retinitis among HIV-positive patients with the IFNL4-ΔG/ΔG genotype compared to those with IFNL4-ΔG/TT and IFNL4-TT/TT genotypes. Similar results have been observed for the risk of CMV replication after solid-organ transplantation, whereby carrying the IFNL4-ΔG allele increases an individual's risk of infection (Manuel and others 2015). Human herpes-virus 8 is the causal agent for Kaposi's sarcoma. The haplotype that produces IFN-λ4 P70 was associated with an increased risk of Kaposi's sarcoma, while that which generates the less active IFN-λ4 S70 form was not (Bibert and others 2018). In a GWAS of mucinous ovarian carcinoma, a rare cancer sometimes confused with a metastasis to the ovary, the IFNL4-ΔG allele had a protective association (Kelemen and others 2015). Genotype for the IFNL4-ΔG/TT variant has also been associated with prostate cancer (Minas and others 2018; Tang and others 2018). Taken together, these findings support the hypothesis that genotype for IFNL4-ΔG/TT variant could affect a wide range of conditions. Further research in this area is clearly needed.
Footnotes
Acknowledgments
The authors would like to thank David Check for his assistance with the figures in the article. This research was supported by the Intramural Research Program of the National Institutes of Health (National Cancer Institute, Division of Cancer Epidemiology and Genetics).
Disclaimer
The content of this publication does not necessarily reflect the views or policies of the Department of Health and Human Services, nor does mention of trade names, commercial products, or organizations imply endorsement by the U.S. Government.
Author Disclosure Statement
No competing financial interests exist.