
Abstract
Genetic variation within the IFNL loci is associated with several diseases and evidence indicates that the IFNL genes have been subjects of strong selection during recent human evolution. The nonsynonymous rs30461 single nucleotide polymorphism (SNP), generating interferon (IFN)-λ1 D188N, shows a strong signature of positive selection in European and Asian populations. Nevertheless, genetic association studies have failed to show any coupling of rs30461 to diseases such as psoriasis and periodontitis. Based on these observations, we purified IFN-λ1 N188 and IFN-λ1 D188 to compare the biological activity of these 2 IFN-λ1 versions. Furthermore, we evaluated the secretion of the 2 different IFN-λ1 versions. We were unable to observe any differences between IFN-λ1 N188 and IFN-λ1 D188 based on biological activity or secretion that could account for the positive selection.
Introduction
Interferons (IFNs)
The induction of type I and type III IFNs is mediated by pattern-recognition receptors (PRRs) in response to invading virus (Onoguchi and others 2007). PRRs detect pathogen-associated molecular patterns and initiate the transcription of type III IFNs through nuclear factor-κB, IFN regulatory factor (IRF) 3, and IRF7. Although type I and type III IFNs utilize distinct receptor complexes, they both signal through the receptor-associated tyrosine kinases, JAK1/TYK2, which mediate the phosphorylation of signal transducer and activator of transcription proteins (Dumoutier and others 2004; Zhou and others 2007). This leads to the assembly of the IFN-stimulated gene factor 3 (ISGF3) complex, which translocates to the nucleus and activates the transcription of ISGs (Durbin and others 2013).
Linkage disequilibrium maps divide the human IFN-λ loci into 3 distinct regions, encoding the IFNL3 and IFNL4 genes, the IFNL2 gene, and the IFNL1 gene, respectively (Manry and others 2011). Genetic variation within the IFNL3/4 region has been mapped to both hepatitis C virus clearance and liver inflammation (Ge and others 2009; Suppiah and others 2009; Tanaka and others 2009; Thomas and others 2009; Rauch and others 2010; Prokunina-Olsson and others 2013; Eslam and others 2015, 2017). An in-depth study of the evolution of human IFNs identified positive selection acting at all 3 type III IFN linkage groups, but in a largely independent manner. Furthermore, this selection differs significantly between different ethnic groups (Manry and others 2011). In particular 1 single nucleotide polymorphism (SNP), known as rs30461 (chr.pos. 39298475 A > G, minor allele frequency [MAF] = 0.2516), displays a strong signature of positive selection in European and Asian populations (Manry and others 2011), and is in fact one of the strongest signatures of positive selection found within the human genome. The rs30461 SNP is located within the last of the 5 exons of the IFNL1 gene and causes a nonsynonymous amino acid substitution at position 188 in the IFN-λ1 protein, thus giving rise to 2 different versions of IFN-λ1, namely IFN-λ1 N188 and IFN-λ1 D188, within the human population. This amino acid substitution is suggested to alter protein function according to the PolyPhen algorithm (Manry and others 2011) and the strong positive selection suggests that altered protein functionality is of significant benefit to the host.
A correlation between the rs30461 SNP and diseases has also been sought without any results so far. Psoriasis patients were genotyped for polymorphisms in the genes encoding the class II cytokines and their receptors and the susceptibility to psoriasis was investigated (Galimova and others 2012). The rs30461 SNP was one of the examined polymorphisms in this study, but no association was found between this SNP and psoriasis. Furthermore, the effect of the SNP was assessed in periodontitis patients (Shivaprasad and Pradeep 2015). This study found an increase of IFN-λ1 in gingival crevicular fluid and plasma of patients with aggressive periodontitis. However, the rs30461 SNP was unrelated to susceptibility to periodontitis.
Therefore, even though the rs30461 SNP displays a selective advantage the mechanism is still unidentified. This article seeks to investigate the effect of the N188D variant upon the activity and secretion of IFN-λ1 protein. IFN-λ1 N188 and IFN-λ1 D188 were purified to compare the activity of the 2 protein variants and the secretion of the 2 different protein versions was evaluated without any significant difference between the 2 variants confirming the findings from the clinical studies.
Materials and Methods
Alignment and model of IFN-λ1
The multiple alignment of IFN-λ1 from different primates was performed in Clustal Omega with default settings from European Bioinformatics Institute and visualized in Jalview 2.10.5. All the primate protein sequences originate from NCBI GenBank with the following accession codes (in order of appearing in the alignment): NP_742152.1, XP_003270428.1, XP_002829244.1, XP_004060753.1, XP_003812294.1, XP_007994951.1, EHH30024.1, XP_011763121.1, XP_003915552.1, XP_011933378.1, XP_011828751.1, XP_011807814.1 and XP_010379221.1.
The modeled location of rs30461 in IFN-λ1 was generated in PyMOL using the structure of human IFN-λ1 and IFN-λ1R (PDB entry code: 3OG6).
Cell lines
HLLR1-2.2, HepG2, and HEK293T cells were cultured in Dulbecco's modified Eagle's medium (DMEM) (Sigma-Aldrich®, Brondby, Denmark) supplemented with 10% (v/v) fetal bovine serum (FBS) (Sigma-Aldrich), 100 U/mL penicillin (Sigma-Aldrich) and 100 mg/mL streptomycin (Sigma-Aldrich). The culture medium for the HLLR1-2.2 cells was additionally supplemented with 1 × HAT (Sigma-Aldrich). When the HepG2 cells were incubated with virus, the medium was supplemented 2% (v/v) FBS.
Plasmids
The pET-15b/N-6xHis IFN-λ1 D188 plasmid for bacterial expression of IFN-λ1 N188 has been described previously (Dellgren and others 2009). Using this plasmid as a template, the pET-15b/N-6xHis IFN-λ1 D188 plasmid for bacterial expression of IFN-λ1 D188 was made by site-directed mutagenesis. The pEF2/IFN-λ1 D188 C-FLAG plasmid for mammalian expression of IFN-λ1 D188 was a kind gift from Sergei Kotenko (Rutgers Cancer Institute of New Jersey, Newark, USA). Using this plasmid as a template, the pEF2/IFN-λ1 N188 C-FLAG plasmid for mammalian expression of IFN-λ1 N188 was made by site-directed mutagenesis.
Protein purification
Human IFN-λ1 N188 and IFN-λ1 D188 were purified as previously described (Dellgren and others 2009).
Activity assay
HLLR1-2.2 cells were seeded in 24-well tissue culture plates at a density of 1.5 × 105 cells per well in 500 μL culture medium the day before titration. For the titration, 2-fold serial dilutions of the cytokines (IFN-λ1 N188 and IFN-λ1 D188) from 2 × 103 to 4.8 × 10−4 ng/mL were prepared with culture medium and the cytokine dilutions were then added to each well. One well was used for control and only culture medium was added. The cells were incubated for 24 h before removing the culture medium and lysing the cells with Cell Culture Lysis 1 × Reagent (Promega, Nacka, Sweden). The lysates were transferred to 1.5 mL tubes and centrifuged (1 min at 10,000 rcf) to remove cell debris. To measure luciferase activity, 10 μL lysate was mixed with 40 μL Luciferase Substrate (OZ Biosciences, Marseille, France) and the luminescence was then measured. The measured luminescence in counts per second (CPS) were analyzed in GraphPad Prism ver. 5, where the EC50 and exponential slope values were calculated by fitting a sigmoidal curve to the plot. Values for baseline were restrained to zero.
Real-time quantitative polymerase chain reaction
HepG2 cells were seeded at a density of 1 × 105 cells per well in 24-well tissue culture plates, and after 24 h the cells were stimulated with culture medium supplemented with 3.5 ng/μL of IFN-λ1 N188 or IFN-λ1 D188 or nothing. After 6 h of incubation, cells were lysed and RNA was purified using E.Z.N.A. Total RNA Kit I (OMEGA bio-tek) according to the manufacturer's instructions. Complementary DNA synthesis was performed with 0.5 μg RNA using RevertAid Reverse Transcriptase and random hexamer primer according to the manufacturer's instructions (ThermoFisher Scientific). The cDNA obtained from cells was quantified by real-time quantitative polymerase chain reaction (RT-qPCR) using SYBR Green I (Roche) and a LightCycler 480 Instrument II (Roche). The following primers were used: GAPDH, forward 5′-GCTCCTCCTGTTCGACAGTCA-3′ and reverse 5′-CCTTCCCCATGGTGTCTGA-3′; MX1, forward 5′-GAAAAATCCAGGCTCGGTGG-3′ and reverse 5′-TCAATGAGGTCGATGCAGGG-3′. The cycling parameters were 95°C for 10 min followed by 45 cycles of 95°C for 10 s, 60°C for 20 s, and 72°C for 12 s. The crossing points of the amplification curves were determined using the second derivative method on the LightCycler 480 Instrument II software 1.5 (Roche). The level of MX1 mRNA was normalized against the GAPDH mRNA content. Significance was tested by an unpaired t-test.
Antiviral assay
The antiviral activity of human IFN-λ1 N188 and IFN-λ1 D188 was measured as previously described (Dellgren and others 2009). The EC50 and exponential slope values were calculated in GraphPad Prism ver. 5 by fitting a sigmoidal curve to the plot. Values for baseline were restrained to zero.
Activity of secreted IFN-λ1 N188 and IFN-λ1 D188
HEK293T cells were seeded in a 12-well tissue culture plate at a density of 4 × 105 cells per well in 1 mL culture medium the day before transfection. The cells were transfected with 500 ng/mL plasmids coding for either IFN-λ1 N188 or IFN-λ1 D188 using polyethylenimine (PolyPlus-Transfection) using the specifications from the manufacturer. The empty plasmid was cotransfected with the IFN-λ1 coding plasmids to maintain equivalent DNA amount. After overnight incubation, the medium was collected and its antiviral activity measured using the assay described previously.
Results
The rs30461 G is the major allele but it is rs30461 A that is the ancestral allele
Before addressing the effect of the rs30461 SNP upon the biological activity and secretion of IFN-λ1 the allele frequencies were examined. According to the SNP gene report for IFN-λ1 from NCBI the SNP is located at chromosome position 39298475 and consists of an A- or G-nucleotide. The A-allele is reported as the major allele and the MAF of rs30461 is 0.2516. As mentioned before, the rs30461 SNP located in the last exon of IFNL1 creates a nonsynonymous amino acid substitution at position 188 in IFN-λ1. The major allele and the minor allele encode IFN-λ1 N188 and IFN-λ1 D188, respectively. When investigating the allele frequencies using the international HapMap project, populations originating from Africa are found to have a higher MAF (0.564–0.759) than non-African populations (0.028–0.144), suggesting that the IFN-λ1 N188 variant is not the ancestral protein (Fig. 1A). To determine the ancestral allele, a multiple sequence alignment using human IFN-λ1 and orthologs from other primate species was created with Clustal Omega. From this alignment (Fig. 1B), it is apparent that only the human IFN-λ1 protein contains the asparagine (N) in position 188, whereas the remaining primate sequences show an aspartate (D) at this position. Even one of the closest living relatives to human, the Pan paniscus (pygmy chimpanzee), has an aspartate (D) at position 188. This suggests that the IFN-λ1 N188 arose in humans at some point after the separation from the last common ancestor and was positively selected for to achieve the common day high allele frequency.

The rs30471 SNP gives rise to 2 variants of IFN-λ1 in the human population.
The amino acid at position 188 of IFN-λ1 does not seem to interact with IFN-λR1
The IFN-λ1 protein interacts with IFN-λR1 through helices A and F as well as the AB loop and with IL10R2 through helices A, C, and D (Miknis and others 2010; Mendoza and others 2017). Residue 188 is expected to be located in or very close to helix F of IFN-λ1 and the rs30461 SNP could thus potentially have an effect upon its binding to the IFN-λR1 receptor chain, which in turn would affect its activity. To explore whether this might be the case, residue 188 was visualized on a model of the crystal structure of IFN-λ1 bound to IFN-λR1 (Fig. 1C). When looking closer at the model of the crystal structure, residue 188 is located right at the C-terminus of helix F of the IFN-λ1 protein and does not appear to interact with IFN-λR1.
The biological activities of IFN-λ1 N188 and IFN-λ1 D188 are equivalent
To compare the activity of IFN-λ1 N188 and IFN-λ1 D188, both proteins were first expressed in Escherichia coli and purified (Fig. 2A). The antiviral activity of the 2 versions of IFN-λ1 was analyzed in HepG2 cells. HepG2 are hepatoma cells that are naturally responsive to type III IFNs. Antiviral activity was measured as the ability of IFN-λ1 to protect HepG2 cells from lysis by encephalomyocarditis virus in an assay previously described (Dellgren and others 2009). Sigmoidal curves were fitted to the data using GraphPad Prism software, allowing for the variables of the EC50, the efficacy (the maximal cell protection induced after IFN stimulation), and the exponential slope. The 2 different IFN-λ1 versions exhibited insignificant differences in their activity (Fig. 2B). We obtained EC50 values of 3.61 ng/mL (confidence interval [CI] 95%: 2.57–5.08) and 3.45 ng/mL (CI 95%: 2.27–5.26) for IFN-λ1 N188 and IFN-λ1 D188, respectively. The efficacies of the IFN-λ1 N188 and IFN-λ1 D188 were 96.74% (CI 95%: 90.22–103.3) and 104.5% (CI 95%: 96.04–112.9). To verify the results, a second protein batch of both IFN-λ1 N188 and IFN-λ1 D188 was purified and analyzed. The second protein batches also showed insignificant differences when analyzed (data not shown).
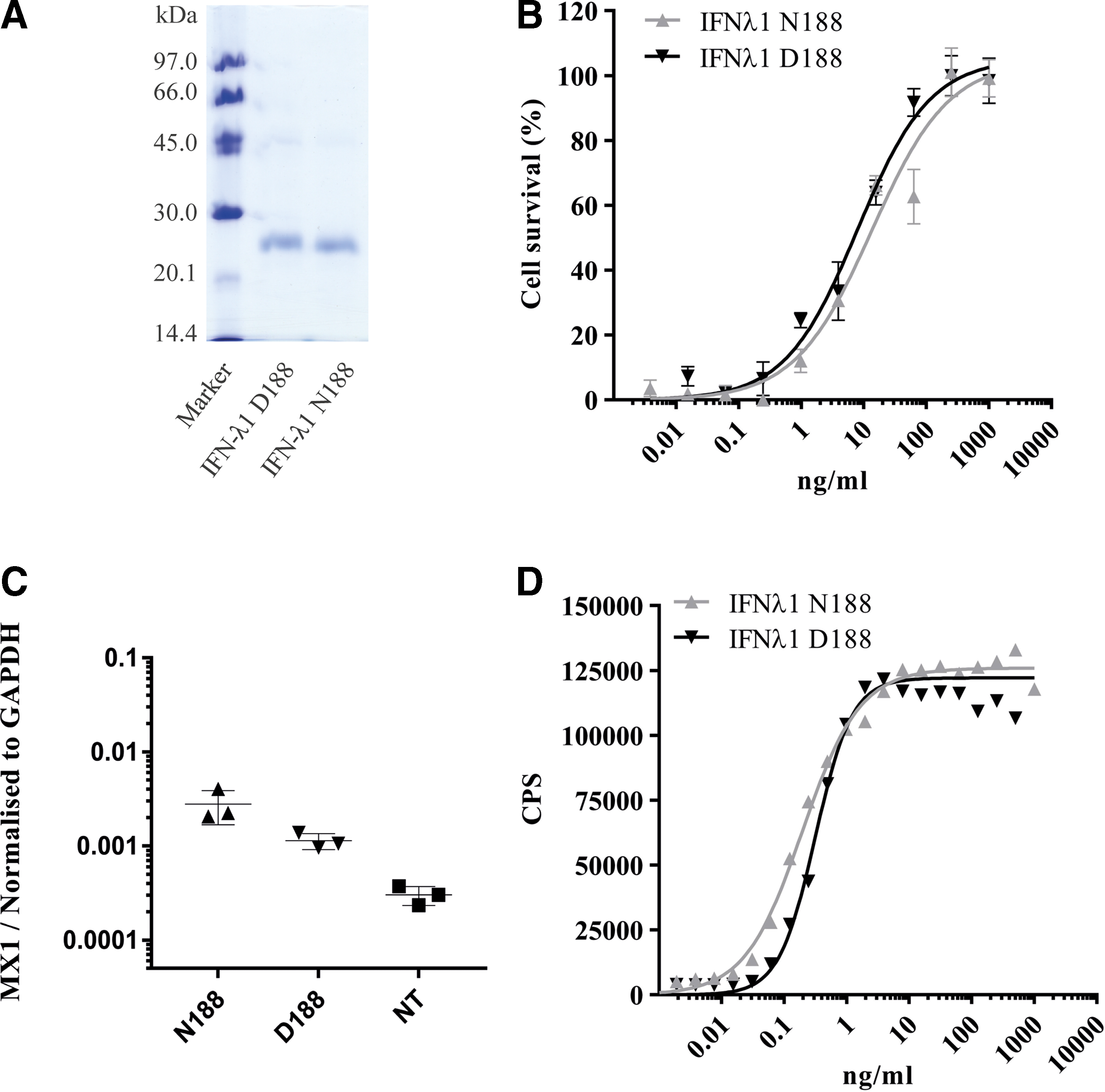
Biological activity of IFN-λ1 N188 and IFN-λ1 D188.
To further investigate the activity of IFN-λ1 N188 and IFN-λ1 D188, we also examined the induction of MX1 in HepG2 cells by RT-qPCR after stimulation with 3.5 ng/mL of either IFN-λ1 N188 or IFN-λ1 D188. We obtained mean values of 0.0028 and 0.0011 (MX expression normalized to GAPDH) for IFN-λ1 N188 and IFN-λ1 D188, respectively (Fig. 2C). Comparison by an unpaired t-test gave a P value of 0.063, displaying a nonsignificant difference between the 2 versions of IFN-λ1.
To confirm that the observation concerning the ISG expression covers other ISGs, we compared the signal in a reporter assay using HLLR1-2.2 cells, which stably express the IFN-λR1 receptor chain and have a firefly luciferase gene under transcriptional control by the IFI6 promoter (Uze and Monneron 2007). After stimulation with different concentrations of IFN-λ1, the luciferase activity was measured and plotted as a function of IFN-λ1 concentration. Same as before, the data were fitted using the GraphPad Prism software, allowing for the variables of the EC50, the efficacy (the maximal luciferase activity after IFN stimulation) and the exponential slope. Again, we did not observe any significant difference in activity between IFN-λ1 N188 and IFN-λ1 D188 (Fig. 2D). We obtained EC50 values of 0.19 ng/mL (CI 95%: 0.16–0.23) and 0.32 ng/mL (CI 95%: 0.21–0.48) for IFN-λ1 N188 and IFN-λ1 D188, respectively. The efficacies of the IFN-λ1 N188 and IFN-λ1 D188 were 125 965 CPS (CI 95%: 122 650–129 280) and 122 187 CPS (CI 95%: 112 703–131 671), respectively. To verify the results, a second protein batch of both IFN-λ1 N188 and IFN-λ1 D188 was purified and analyzed. The second protein batches also showed insignificant differences when analyzed (data not shown).
The rs30461 SNP does not affect protein secretion
Since IFN-λ1 is secreted after translation and functions extracellular, it was tested whether the nonsynonymous rs30461 SNP affected protein secretion. For this purpose, HEK293T cells were transfected with empty plasmid alone or plasmids encoding either IFN-λ1 N188 or IFN-λ1 D188 and by measuring the antiviral activity of the culture medium as previously, secretion of IFN-λ1 was then assessed. We could not observe any antiviral activity in culture medium from cells transfected with empty plasmid, whereas the culture medium from cells transfected with plasmids encoding IFN-λ1 showed significant antiviral activity (Fig. 3). However, the difference between the 2 different versions of IFN-λ1 was insignificant with EC50 values of 3.48 μL/mL (CI 95%: 2.61–4.63) and 3.84 ng/mL (CI 95%: 2.83–5.20) for IFN-λ1 N188 and IFN-λ1 D188, respectively. Assuming similar specific activities between the IFN-λ1 made in E. coli and the IFN-λ1 secreted by the human cells, the observed antiviral activity of the culture medium corresponds to ∼900–1,000 ng/mL IFN-λ1. To verify our result, the transfection was repeated twice and although the antiviral activity of the medium varied slightly between different transfections, the difference between IFN-λ1 N188 and IFN-λ1 D188 remained insignificant (data not shown).
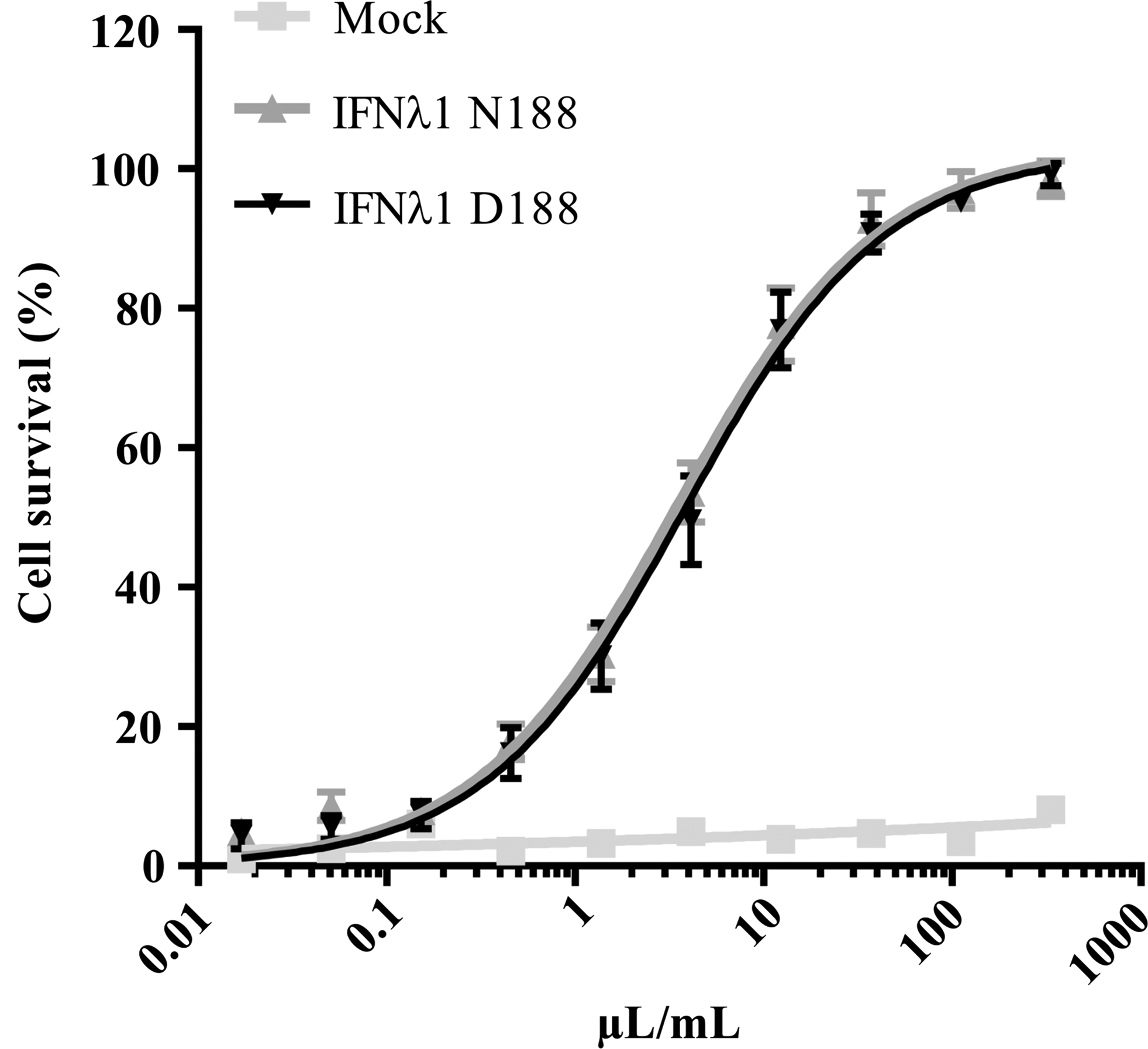
Secretion of IFN-λ1 N188 or IFN-λ1 D188. HEK293T cells were transfected with plasmids expressing IFN-λ1 N188 or IFN-λ1 D188, respectively, or empty plasmid as control. The amount of secreted IFN-λ1 N188 or IFN-λ1 D188 was then measured by incubating HepG2 cells with serial dilutions of the medium from transfected the cells before challenging them with EMCV. Data are presented as a percentage of a noninfected cell control. Error bars depict SD of 4 replicates.
Discussion
In this study, we demonstrate that despite genetic indications for positive selection, the activity and secretion of IFN-λ1 is unaffected by the nonsynonymous rs30461 SNP. When comparing the results from the activity assays to the crystal structure model of IFN-λ1 and its high-affinity receptor chain (Fig. 1C), it can be noticed that residue 188 does not seem to interact with IFN-λR1. This lack of interaction confirms the results from this study showing that the amino acid substitution caused by the rs30461 SNP does not influence the activity of the protein but is in stark contrast to the strong positive selection observed at this site by Manry and others (2011). These findings support the previous discoveries of Galimova and others (2012) and Shivaprasad and Pradeep (2015) who investigated the rs30461 SNP in psoriasis and periodontitis, respectively, without any significant findings.
We reinvestigated the allele frequencies in publicly available databases, as a significant amount of novel genetic information has been made available since the original study in 2011 and we confirming the findings by Manry and others (2011). The non-African populations have lower MAFs compared with African populations. By comparing the major allele encoded protein in the human population, the IFN-λ1 N188 variant, with orthologs from other primate species, it became evident that the ancestral variant is the IFN-λ1 D188 encoded by the rs30461 minor allele. This observation not only further supports the conclusion presented by Manry and others (2011) but also leaves us with an enigma: Why do we observe a large positive selection signature at this site when we are not able to observe any change in protein function?
There are several possible explanations for these discrepancies. It is possible that the genetic differences in the human population are a result of a genetic bottleneck from the early human migrations out of African. This could explain why IFN-λ1 N188 is overrepresented in the non-African populations. However, this explanation does not account for the fact that the rs30461 major allele is maintained in the non-African populations. Alternatively, it is possible that the selective pressure on the rs30461 SNP is maintained because of a confounder effect. However, Manry and others (2011) do not identify other SNPs in proximity to the rs30461 SNP. Nor has other SNPs, to the authors' knowledge, been found linked to the rs30461 by whole genome sequencing projects, making this scenario unlikely. Further studies of IFNL1 genetics in both humans and other species could elucidate whether the distribution of the rs30461 SNP in the human population is truly due to a positive selection or a confounder effect. Our study only investigates the influence of the rs30461 SNP on the protein itself. It is possible that the rs30461 SNP is having an epigenetic influence on the IFNL1 gene requiring further studying.
The selective advantage behind the rs30461 SNP found in genetic investigations is still unclear and the discrepancy between the results presented in this article as well as the clinical findings and the genetic observations makes it obvious that our knowledge about the impact of genetic variants is insufficient. With the current knowledge in the field, we are unable to explain the occurring observations. Whether the apparent evolutionary benefit needs to be sought in further biochemical examinations or in further disease association studies is uncertain, but further research is required.
Footnotes
Acknowledgments
This project was financially supported by the Riisfort Foundation, the Toyota Foundation, and the Danish Council for Independent Research, Medical Research (grant 7016-00331B).
Author Disclosure Statement
No competing financial interests exist.