
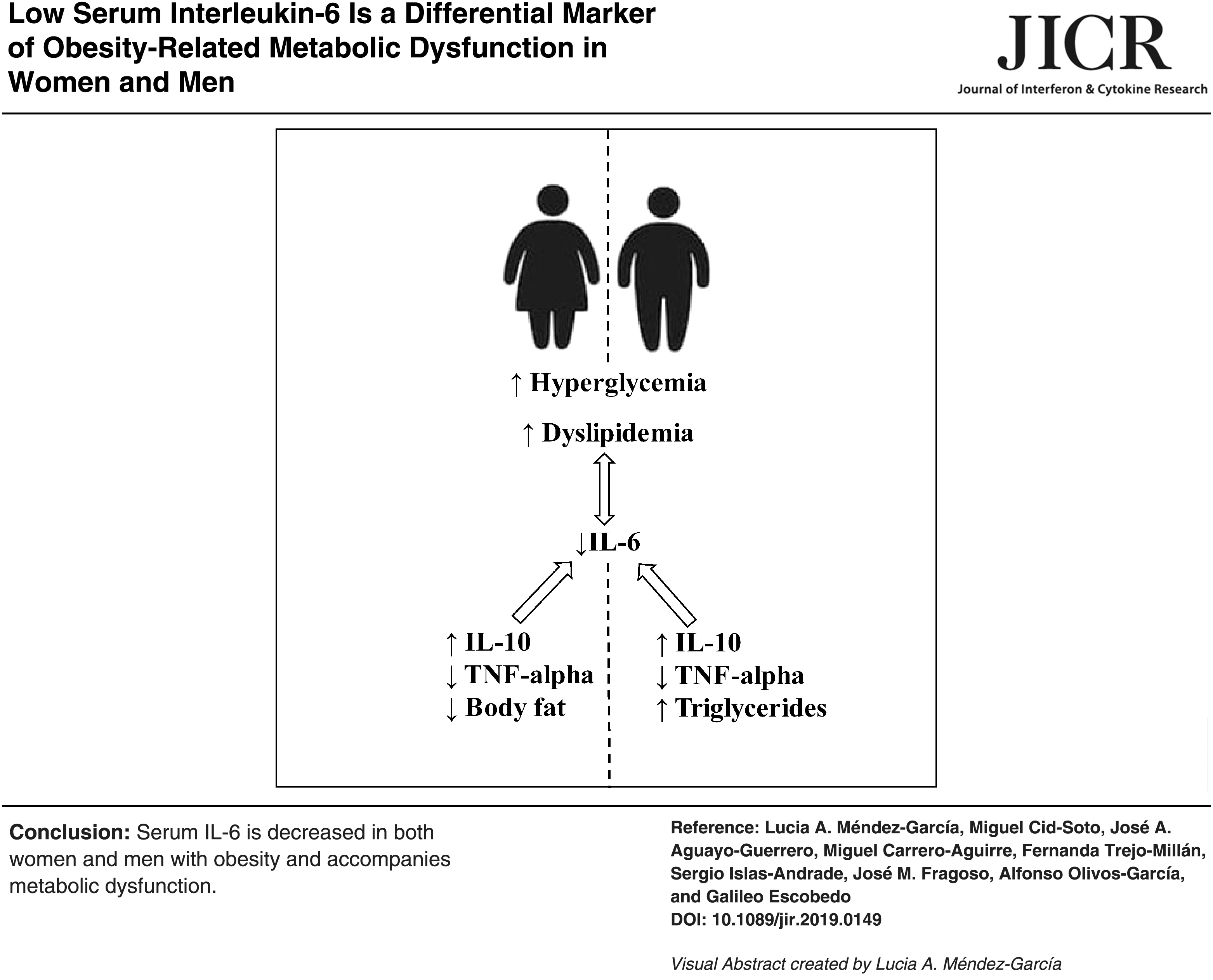
Abstract
There is scant information regarding the role of interleukin (IL)-6 in obesity-related metabolic dysfunction in humans. Thus, we studied the serum levels of IL-6 in normal weight, overweight, and obese subjects, and examined associations of IL-6 with hyperglycemia, insulin resistance, dyslipidemia, and systemic inflammation. One hundred three women and men were included in the study. Anthropometric parameters, blood glucose, insulin, total cholesterol, and triglycerides were measured. Serum levels of tumor necrosis factor-alpha (TNF-alpha), IL-10, and IL-6 were measured by enzyme-linked immunosorbent assay (ELISA). One-way analysis of variance (ANOVA) showed a 2.5-fold significant decrease in serum IL-6 in overweight and obese individuals when compared with normal weight controls. Serum IL-6 exhibited significant inverse correlations with body mass index (r = −0.39/P < 0.0001), waist circumference (r = −0.42/P < 0.001), blood glucose (r = −0.40/P < 0.0001), triglycerides (r = −0.34/P < 0.0001), and TNF-alpha (r = −0.48/P < 0.0001), whereas a strongly positive correlation was found with IL-10 (r = 0.77/P < 0.0001). Multiple linear regression analysis revealed that behavior of IL-6 was mainly influenced by IL-10 (beta = 0.28/P = 1.95 × 10−6), TNF-alpha (beta = −0.67/P = 0.0017), and body fat percentage (beta = −5.95/P = 7.67 × 10−5) in women. In contrast, IL-10 (beta = 0.37/P = 1.34 × 10−9), TNF-alpha (beta = −0.85/P = 0.0005), and triglycerides (beta = 1.07/P = 0.0007) were major influencing factors of IL-6 in men. This study demonstrates that IL-6 is a marker of metabolic dysfunction that is differentially regulated in obese women and men.
Introduction
Obesity affects millions of people worldwide with alarmingly high prevalence in the Mexican population that in turn increases the risk of developing insulin resistance, hyperglycemia, dyslipidemia, type 2 diabetes (T2D), and atherosclerosis (Ferrante 2007; Gnacinska and others 2009; DiBonaventura and others 2018).
Obesity is accompanied by a state of low-grade chronic systemic inflammation that is characterized by increased levels of proinflammatory cytokines including tumor necrosis factor-alpha (TNF-alpha) and interleukin (IL)-6, and reduced values of anti-inflammatory mediators such as IL-10 (Esser and others 2014). TNF-alpha has been shown to decrease phosphorylation of the insulin receptor substrate 2, which in turn reduces translocation of glucose transporter proteins into the cell membrane of adipocytes, hepatocytes, and skeletal muscle cells (Recasens and others 2004). IL-10 has been described to block TNF-alpha actions on adipose and skeletal muscle cells and numerous studies have shown that progressive loss of this cytokine is associated with increased insulin resistance (Dagdeviren and others 2017). However, the role of IL-6 in the development of insulin resistance and other metabolic disturbances remains elusive (Kern and others 2001).
IL-6 is a pleitropic cytokine that is secreted by macrophages, adipocytes, fibroblasts, and endothelial cells, and has prominent functions in lipid metabolism and body weight gain (Sindhu and others 2015). IL-6 has been shown to exert both protective and deleterious actions in obesity and insulin resistance. For instance, several studies have shown that serum levels of IL-6 increase in accordance with the level of adiposity in obese subjects (Bluher and others 2005; Park and others 2005). In contrast, a previous work reported that IL-6 promotes energy expenditure and glucose homeostasis in mice fed a high-fat diet, while also favors nutrient homeostasis and insulin sensitivity (Sadagurski and others 2010). Recent studies also show that administration of exogenous IL-6 in obese mice inhibits food intake and increases glucose metabolism by stimulating directly the paraventricular nucleus of the hypothalamus (Timper and others 2017).
As it can be seen, the role of IL-6 in obesity, insulin resistance, glucose homeostasis, dyslipidemia, and systemic inflammation is still controversial. Therefore, our main goal was to study the serum levels of IL-6 in a Mexican population of normal weight, overweight, and obese women and men, while also examining potential associations of IL-6 with parameters related to obesity, insulin resistance, hyperglycemia, lipid profile, and systemic inflammation.
Materials and Methods
Subjects
A total of 143 healthy Mexican adult volunteers from the south-central region of Mexico attending to the blood bank of the General Hospital of Mexico were included in the study. Participants provided written informed consent, previously approved by an Institutional Review Board of the General Hospital of Mexico, which guaranteed that the study was conducted in accordance with the principles described in the 1964 Declaration of Helsinki and its posterior amendment in 2013. Exclusion criteria were if subjects had previous or recent diagnosis of diabetes mellitus, cardiovascular diseases, chronic hepatic or renal disease, blood pressure higher than 140/90 mm Hg, inflammatory or autoimmune disorders, acute or chronic infectious diseases, cancer, and endocrine disorders. Additionally, pregnant or lactating women, and subjects under any kind of anti-inflammatory, immunomodulatory, hypolipidemic, antiaggregant, and antihypertensive medication were excluded from the study.
Anthropometric measurements
All of the participants were divided into 3 groups according to the World Health Organization criteria for body mass index (BMI) as follows: normal weight controls (BMI 18.5–24.9 kg/m2), overweight individuals (BMI 25–29.9 kg/m2), and obese subjects (BMI ≥30 kg/m2). BMI resulted of dividing weight by height squared (kg/m2). Waist circumference and body fat percentage were also determined in all study subjects. Specifically, waist circumference was measured considering the midpoint between the lower rib margin and the iliac crest, using a conventional tape in centimeters (cm). Abdominal obesity for women was considered when their waist circumference were 80 cm or higher, whereas waist perimeter of 90 cm or higher was considered as the presence of central obesity for men. Body fat percentage was measured using a body composition analyzer (TANITA®, Body Composition Analyzer, Model TBF-300A, Tokyo, Japan).
Biochemical measurements
After overnight fasting, blood samples were individually collected into pyrogen-free tubes (Vacutainer™; BD Diagnostics, NJ) at room temperature. Then tubes were centrifuged at 1,000 g/4°C for 30 min, and serum samples obtained and stored at −80°C in numerous aliquots until use. Total cholesterol and triglyceride were individually measured in triplicate by an enzymatic assay according to manufacturer's instructions (Roche Diagnostics, Mannhein, Germany). Serum insulin levels were determined in triplicate by the enzyme-linked immunosorbent assay (ELISA), following the manufacturer's instructions (Abnova Corporation, Taiwan). Serum glucose levels were individually determined in triplicate by the glucose oxidase assay, following the manufacturer's instructions (Megazyme International, Ireland). The estimate of insulin resistance was individually calculated by means of the homeostatic model assessment of insulin resistance (HOMA-IR), as follows: fasting insulin concentration (mU/L) × fasting glucose concentration (mM) divided by 22.5.
Immunologic measurements
Measurements of the serum levels of IL-6, TNF-alpha, and IL-10 were performed in each study subject. Briefly, blood samples were individually collected into pyrogen-free tubes (Vacutainer; BD Diagnostics) at room temperature. Collection tubes were then centrifuged at 1,000 g/4°C for 30 min, and serum samples obtained and stored at −80°C until use. Serum levels of IL-6, TNF-alpha, and IL-10 were determined in triplicate by ELISA, following the manufacturer's instructions (Peprotech, Mexico).
Statistics
Data from BMI, waist circumference, body fat percentage, fasting blood glucose, serum insulin, HOMA-IR, total cholesterol, triglycerides, serum TNF-alpha, IL-10, and IL-6 were analyzed by means of the Mann–Whitney U test or the Kruskal–Wallis test corrected by multiple comparisons using the Dunn's test (depending on the number of groups), and are expressed as median and inter-quartile range in a box plot analysis. The Spearman's correlation coefficients were calculated for examining the relationship of IL-6 with parameters related to obesity (BMI, waist circumference, and body fat percentage), glucose homeostasis (fasting blood glucose, serum insulin, and HOMA-IR), lipid metabolism (total cholesterol and triglycerides), and systemic inflammation (TNF-alpha and IL-10). All studied groups were matched by gender and age. Statistical analysis was performed using the GraphPad Prism 5 software. A multiple regression model was performed in the terminal R 3.5.1. All the parameters measured in this study were introduced as independent variables, to determine the influence of the aforementioned variables on the serum levels of IL-6 in the whole population and divided by gender. Data from multiple regression analysis are expressed as β-values and confidence intervals of 2.5% and 97.5%. Differences were considered significant when P < 0.05.
Results
As expected in a sex and age-matched study, we found no significant differences in age and women/men proportion among normal weight, overweight, and obese subjects (Table 1). In contrast, waist circumference, body fat percentage, fasting blood glucose, serum insulin, HOMA-IR, triglycerides, and serum TNF-alpha showed a significant increase in overweight and obese subjects when compared with normal weight controls (Table 1). Moreover, obese subjects exhibited a significant increase in total cholesterol with respect to normal weight controls but not overweight individuals (Table 1). In parallel, serum levels of IL-10 significantly decreased in overweight and obese subjects when compared with normal weight controls (Table 1).
Anthropometric and Biochemical Parameters of the Study Subjects
Data are presented as mean ± standard deviation. Differences were considered significant when P < 0.05, when comparing normal weight subjects versus overweighta and obeseb individuals.
a.u., arbitrary units; BMI, body mass index; HOMA-IR, homeostatic model assessment of insulin resistance; IL-10, interleukin 10; NS, nonsignificant; TNF-alpha, tumor necrosis factor-alpha.
The serum levels of IL-6 showed a significant decrease in overweight and obese subjects when compared with normal weight controls. The mean value of IL-6 in normal controls was 25.64 ± 15.12 pg/mL, whereas in obese and overweight individuals it was significantly reduced to 8.79 ± 4.54 and 12.02 ± 4.16 pg/mL respectively (Fig. 1A). When studied in terms of abdominal obesity, serum IL-6 levels decrease in subjects with increased waist circumference (10.17 ± 3.69 pg/mL) when compared with individuals showing normal waist perimeter (20.17 ± 14.30) (P < 0.05) (Fig. 1B).
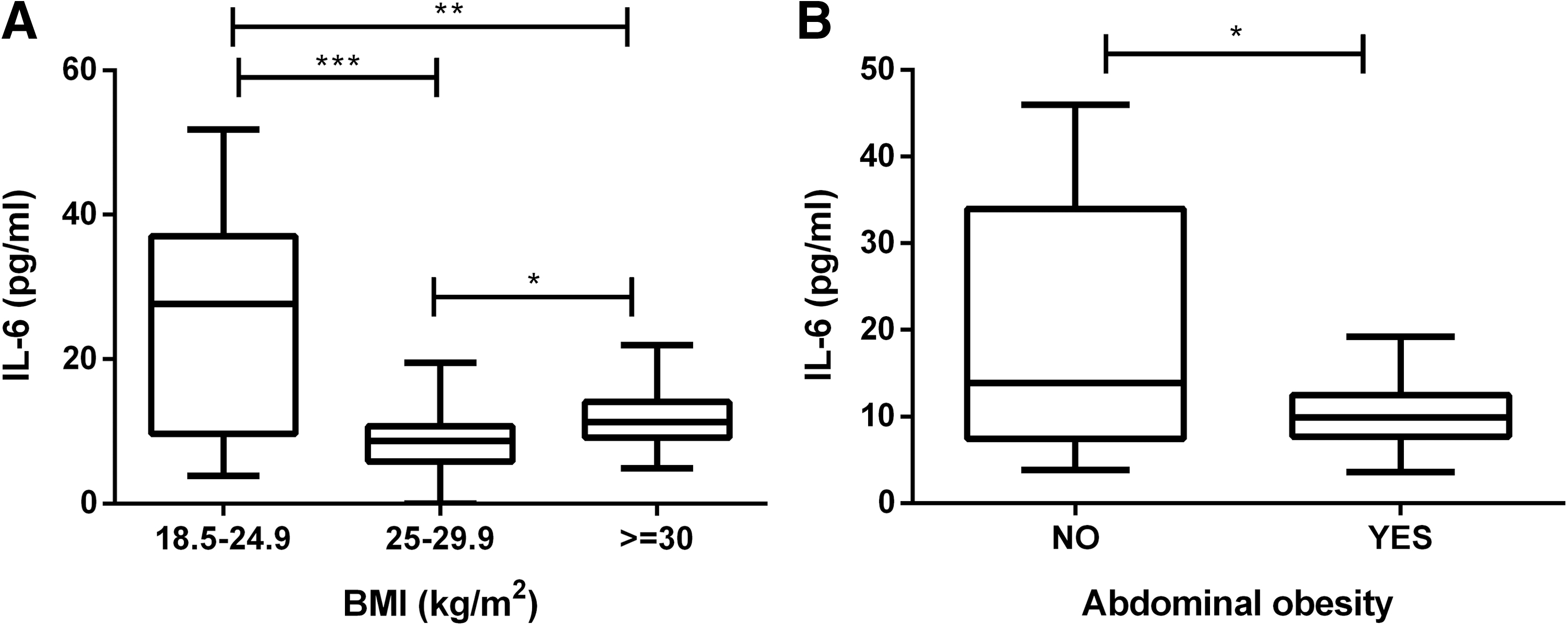
Serum levels of IL-6 in obese and nonobese subjects.
IL-6 serum levels showed an inverse correlation with BMI, where IL-6 significantly decreases as BMI increases (r = −0.3927, P < 0.0001) (Fig. 2A). As expected, we also found an inverse correlation between IL-6 serum levels and waist circumference in the study population (r = −0.4186, P < 0.001) (Fig. 2B). On the contrary, IL-6 serum levels had a weak inverse correlation with body fat percentage (r = −0.1746, P = 0.0370) (Fig. 2C).
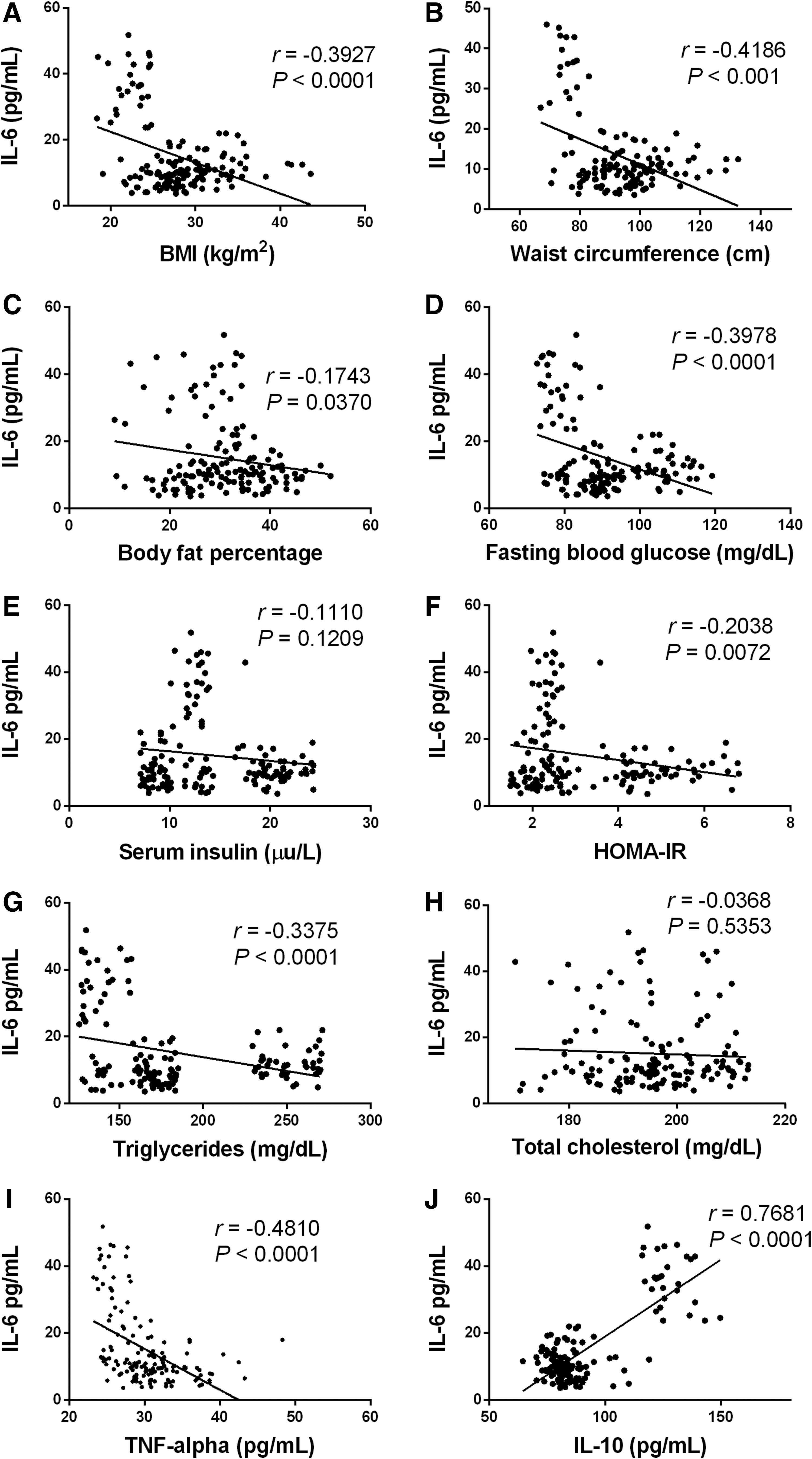
Statistical correlations of IL-6 with parameters related to obesity, glucose homeostasis, dyslipidemia, and systemic inflammation. Serum levels of IL-6 were inversely correlated with BMI
With regard to parameters related to insulin resistance, IL-6 serum values showed inverse correlations with fasting blood glucose (r = −0.3978, P < 0.0001) and HOMA-IR (r = −0.2038, P = 0.0072) (Fig. 2D, E, respectively) but failed to associate with serum insulin (r = −0.1110, P = 0.1209) (Fig. 2F). In terms of lipid metabolism, IL-6 serum levels exhibited an inverse correlation with triglyceride values (r = −0.3375, P < 0.0001) (Fig. 2G). On the other side, no significant association was found between IL-6 and total cholesterol levels (r = −0.0368, P = 0.5353) (Fig. 2H).
With respect to markers of low-grade chronic inflammation, IL-6 serum values exhibited a significant inverse relationship with TNF-alpha (r = −0.4810, P < 0.0001) (Fig. 2I). On the contrary, IL-6 had a strongly positive correlation with IL-10 serum levels (r = 0.7681, P < 0.0001) in the study population (Fig. 2J).
Multiple linear regression models revealed that both TNF-alpha and IL-10 differentially predict about 68% of the variability observed in the serum levels of IL-6 in the whole population (P < 2 × 10−16), where TNF-alpha was inversely associated, but IL-10 showed a positive correlation (Table 2). When predictor variables were analyzed by gender, multiple linear regression analysis showed that IL-10, TNF-alpha, and body fat percentage are able to explain about 73% of the variability seen in the serum levels of IL-6 in women (P < 2 × 10−16) (Table 3). In contrast, IL-10, TNF-alpha, and triglycerides predicted about 66% of the variability observed in IL-6 serum values of men (P = 1 × 10−12) (Table 3).
Multiple Linear Regression Model to Determine the Main Influencing Factors on Interleukin-6 Serum Levels in the Whole Study Population
Multiple linear regression model was performed under the following criteria: CI: 2.5%–97.5%, residual standard error: 65.44 on 136 degrees of freedom. Multiple R-squared: 0.6812, Adjusted R-squared: 0.6672, F-statistic: 48.44 on 6 and 136 degrees of freedom, P < 2 × 10−16.
β, beta value; CI, confidence interval.
Multiple Linear Regression Models to Determine the Main Influencing Factors on Interleukin-6 Serum Levels in Women and Men Separately
For women, multiple linear regression model was performed under the following criteria: CI: 2.5%–97.5%; residual standard error: 70.58 on 72 degrees of freedom. Multiple R-squared: 0.7294, Adjusted R-squared: 0.7181, F-statistic: 64.7 on 3 and 72 degrees of freedom, P < 2 × 10−16. For men, multiple linear regression model was performed under the following criteria: CI: 2.5%–97.5%, residual standard error: 45.97 on 60 degrees of freedom. Multiple R-squared: 0.6646, Adjusted R-squared: 0.6311, F-statistic: 19.82 on 6 and 60 degrees of freedom, P = 1 × 10−12.
Discussion
IL-6 is a member of the 4-alpha-helix cytokine family with functions classically related to the inflammatory response (Jones and Vignali 2011). However, a growing number of studies has suggested that IL-6 is also involved in obesity and obesity-related comorbidities but its role is still unclear (Sindhu and others 2015). As a matter of fact, IL-6 has been positively correlated with adiposity in Mexican American obese children, in which serum levels of IL-6 rise as the degree of obesity increases (McFarlin and others 2007). Similarly, IL-6 and TNF-alpha serum levels tend to elevate in Mexican American adults diagnosed with T2D (Mirza and others 2012). On the contrary, administration of recombinant human IL-6 (rhIL-6) increases lipolysis and fatty acid esterification in healthy humans without showing deleterious secondary-effects (van Hall and others 2003). Likewise, a study conducted in Mexican adolescents from the central-region of Mexico showed that IL-6 and TNF-alpha serum levels are lower in overweight adolescents than in normal weight controls (Pardo-Morales and others 2011). Notably, our data concur with latter evidence by demonstrating that IL-6 serum levels have an inverse correlation with BMI and waist circumference, where IL-6 increases as BMI and central obesity decrease. These apparently controversial results may be explained by considering that despite Mexican American individuals and Mexican subjects show similar genetic background, they have different diet and lifestyle that in turn may influence the serum levels of IL-6 (Eder and others 2009). For instance, it has been previously reported that nondiabetic normal subjects show increased IL-6 serum levels when feeding a high-fat diet (Nappo and others 2002). Additionally, it has been demonstrated that circulating levels of IL-6 are produced by contracting muscles during exercise (Pedersen and Fischer 2007). For these reasons, further studies should be conducted in normal weight and obese individuals taking into account additional factors that may modify IL-6 serum levels including genetic background, physical activity, and diet.
Another phenomenon captured in our study involves the relationship of IL-6 serum levels with carbohydrate and lipid metabolism. A previous study demonstrated that healthy men performing leg extension exercises show increased IL-6 serum levels that in turn are positively related to exercise intensity and muscle glucose uptake, thus suggesting a role for IL-6 as a possible glucose sensor in the body (Helge and others 2003). In parallel, in vitro exposure of primary human skeletal muscle cells to rhIL-6 induces increased expression of the fatty acid transporter 4 and oxidation of palmitic acid (Al-Khalili and others 2006). Concurring with prior data, our study reveals that IL-6 serum levels increase while blood glucose and triglycerides values decrease in normal weight subjects, leading to the notion that IL-6 has an inverse relationship with the aforementioned parameters in healthy conditions.
Additionally to the inverse correlation with BMI, waist circumference, blood glucose, and triglycerides, serum IL-6 was importantly associated with both TNF-alpha and IL-10. TNF-alpha has been shown to play prominent inflammatory role in multiple pathologic scenarios including acute infections, chronic inflammatory diseases, obesity, T2D, glucose intolerance, and insulin resistance (Nieto-Vazquez and others 2008). Interestingly, numerous studies have shown that IL-6 is able to exert anti-inflammatory effects by inhibiting TNF-alpha expression (Tilg and others 1994). Furthermore, it has been previously reported that hrIL-6 infusion accompanied by physical activity is able to inhibit endotoxin-induced TNF-alpha production in humans (Starkie and others 2003). Notably, Starkie and coworkers suggested that negative regulation of IL-6 on TNF-alpha expression may associate with improved insulin sensitivity in humans. In this sense, our results appear to concur with previous evidence by revealing that IL-6 serum levels are inversely associated with TNF-alpha and blood glucose. The possible mechanism by which IL-6 is capable of downregulating the serum levels of TNF-alpha involves the role of IL-10 and will be further discussed.
Recent experiments in mice have shown that IL-6 has regulatory effects on the central nervous system (CNS). Upregulation of IL-6 in the CNS improves energy expenditure and glucose homeostasis, and reduces body weight gain in obese mice by inducing STAT3 activation (Timper and others 2017). STAT3 is a transcription factor in charge of orchestrating IL-10 expression in nearly all leukocytes including macrophages, neutrophils, eosinophils, mast cells, and B cells, among other (Hutchins and others 2013). As a matter of fact, a previous work reported that IL-6 requires the expression of IL-10 to suppress hyperphagia and insulin resistance by inhibiting the nuclear factor kappa B-dependent signaling pathway in mice (Ropelle and others 2010). Interestingly, our data agree with previous information by revealing a very strong positive correlation of IL-6 with IL-10, and suggest that IL-6 may reduce TNF-alpha serum levels by promoting IL-10 production. Multiple linear regression analyses support such hypothesis by demonstrating that IL-6 serum levels are mainly influenced by IL-10 and TNF-alpha systemic values in the whole population. However, our work is only an associative study and further research is needed to elucidate the mechanisms through which IL-6 would be able to inhibit TNF-alpha expression by upregulating IL-10 in nonobese and obese individuals.
Finally, our results also show that IL-6 serum levels are influenced by different factors in women and men. In fact, IL-6 serum levels are better explained by IL-10, TNF-alpha, and body fat percentage in women. On the other hand, IL-6 systemic values are better explained by IL-10, TNF-alpha, and triglycerides in men. Notably, it has been previously reported that South Asian women show increased serum IL-6 when compared with European women and that in turn is directly related to body fat percentage (Peters and others 2013). Thus, it is feasible to suppose that IL-6 production is not only influenced by gender but also genetic background and ancestry. On the contrary, another study demonstrated that IL-6 serum levels are intimately related to impaired insulin action and play a role as a proatherogenic factor in Caucasian men but not women (Fernandez-Real and others 2001), thus suggesting a gender-dependent function for IL-6. Additionally, it has been also reported that IL-6 is differentially expressed in woman and men (Bonafe and others 2001). Current evidence concurs with our results by demonstrating that IL-6 serum levels are influenced by gender-specific factors that are differentially produced in women and men. Therefore, additional clinical and experimental studies are needed to characterize the biological factors that are potentially capable of modulating IL-6 production in a sex-dependent fashion.
In conclusion, this study demonstrates that serum levels of IL-6 decrease in overweight and obese subjects with respect to normal weight controls, and are inversely correlated with BMI, waist circumference, blood glucose, triglycerides, and serum TNF-alpha. Our results also show for the first time that IL-6 serum levels increase as IL-10 elevates, which in turn might be associated with low circulating values of TNF-alpha in normal weight subjects. This study supports the role of IL-6 as a marker of obesity, hyperglycemia, dyslipidemia, and systemic inflammation in humans, and encourages further research to evaluate its usefulness in monitoring patients with metabolic syndrome and T2D.
Footnotes
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported by grant no. SALUD-2017-02-290345 from the Fondo Sectorial de Investigación y Desarrollo en Salud y Seguridad Social SS/IMSS/ISSSTE/CONACYT-México to G.E. L.A.M.-G. is a Posdoctoral Research Fellow in the General Hospital of Mexico that has been supported by grant no. SALUD-2017-02-290345.