
Abstract
Bone marrow mesenchymal stem cells (BM-MSCs) are cells with the potential to differentiate into adipocytes in oxidative stress. In this study, tert-butyl hydroperoxide is used as a stimulator that promotes reactive oxygen species in BM-MSCs. The results demonstrate that knockdown of LKB1 inhibits the transformation of BM-MSCs into adipocytes in the presence of oxidative stress. In addition, β3 adrenergic receptor agonists, a positive stimulatory molecule for the transformation of BM-MSCs into adipocytes, restores the transformation ability of BM-MSCs caused by LKB1-siRNA. As an upstream signal of adenosine monophosphate-activated protein kinase (AMPK), LKB1 activates the AMPK pathway and promotes the expression of PPARγ and CCAAT/enhancer binding proteins (C/EBPα). This indicates that the regulation of LKB1 on BM-MSCs is dependent on the AMPK pathway. Immunofluorescence localization experiments reveal that the LKB1 and AMPK localizations partially overlap, oxidative stress promotes their expression in the cytoplasm. In general, LKB1 promotes the transformation of BM-MSCs to adipocytes by activating AMPK pathway under oxidative stress.
Introduction
Bone marrow mesenchymal stem cells (BM-MSCs) are multidirectionally differentiated and self-renewing precursor cells (Chen and others 2017; Xu and others 2017; Mohanty and others 2018). BM-MSCs play an important role in certain fields through the induction differentiation of BM-MSCs, for example, inducing BM-MSCs to differentiate into cartilage that are common sources of seeding cells for cartilage tissue engineering with low-intensity pulsed ultrasound (Zuo and others 2013). Scaffold spatial structures have been shown to regulate the proliferation and differentiation of BM-MSCs in 3D directions, which may be used for the treatment of nerve defects by bridging the peripheral nerve gap (Chen and others 2017). The upregulation of CCAAT/enhancer binding proteins (C/EBPα) and PPARγ leads to the differentiation of BM-MSCs into adipogenic progenitors (Cao and others 1991). However, marrow fat accumulation impeded bone formation, suggesting that the differentiation of BM-MSCs into adipocytes may lead to the inhibition of the function of BM-MSCs (Moerman and others 2004; Huang and others 2016). So, could differentiation of adipocytes from BM-MSCs provide energy source under certain conditions?
Adipose tissue is composed of white adipose tissue (WAT) and brown adipose tissue, and plays crucial roles in energy metabolism (Qiao and others 2016; Higareda-Almaraz and others 2018). Research shows that increased GATA2 expression would suppress differentiation of BM-MSCs to adipocytes (Kamata and others 2014). Instead, miR-26a shifted mitochondrial morphology toward brown adipocytes by inducing pathways related to energy dissipation (Karbiener and others 2014). However, oxidative stress interferes with the normal fat metabolism process by regulation of the process of intracellular signaling and transcription factors, such as Notch signaling (Maslov and others 2019) and C/EBPα (Ba and others 2012). The mechanism of the relationship between BM-MSCs and adipogenesis was complicated, and oxidative stress may be a bridge between their differentiation.
Lkb1 (serine/threonine kinase 11) was a member of the serine/threonine kinase family, and regulates the process of cellular metabolism, growth, and immune balance through adenosine monophosphate-activated protein kinase (AMPK) signaling (Fogarty and others 2016; Wang and others 2019). Interestingly, oxidative stress inhibited proliferative effect that was associated with apoptosis and necrosis by changing AMPK signaling (Queiroz and others 2015). However, the relationship between the Lkb1/AMPK signaling pathway and the differentiation of BM-MSCs has not been reported.
In this study, we first demonstrated that LKB1 promotes the conversion of BM-MSCs to adipocytes in oxidative stress. More importantly, the regulation of LKB1 on BM-MSCs was dependent on the AMPK pathway. The proposed pathway explains the differentiation mechanism of BM-MSCs partially, and provides a theoretical basis for the rational utilization of BM-MSCs.
Materials and Methods
Cell culture
BM-MSCs were purchased from ATCC (PCS-500-012) and cultured in standard cell in culture medium in a constant temperature incubator. The oxidative stress cell model was BM-MSCs pretreated by 1.5 mmol/L tert-butyl hydroperoxide.
In vitro cell proliferation (3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl tetrazolium bromide) assay
MTT was dissolved at a final concentration of 5 mg/mL in medium. The medium in each culture well was delicately discarded before adding 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl tetrazolium bromide (MTT) solution. After incubation for 4 h (37°C, 5% CO2), DMSO was added in each culture well instead of medium, and the absorption value was tested at 570 nm wavelengths by a microplate reader.
Oil Red O staining
Pretreated cells were fixed by 4% cold paraformaldehyde for 20 min in cell culture plates and stained with working concentration of oil red dye solution for 15 min in the dark. Sixty percent ethanol was used to differentiate the stained cells with a colorless background. The cells were washed in distilled water and counterstained in Mayer hematoxylin. The cells were washed again and observed and analyzed in a microscope.
Western blot
Pretreated cells were collected and lysed in cell lysis buffer, and 30 μg of protein of every sample was separated by sodium dodecyl sulfate-polyacrylamide gel electrophoresis and transferred to a PVDF membrane. After blocking by skim milk solution for 1 h, the cells were incubated with primary antibodies at 4°C overnight. After phosphate-buffered saline washing and incubating with secondary antibodies for 1 h, the bands were visualized using enhanced chemiluminescence reagent.
Immunofluorescence assay
The BM-MSCs were seeded in a 24-well cell culture plate, grew to 20% confluence, fixed by 4% cold paraformaldehyde for 20 min, and then treated with triton X-100 for 20 min. After blocking with serum, the cells were incubated with primary antibodies at 4°C overnight. Then they were incubated with 2 different types of secondary antibodies for 2 h at room temperature. Then DAPI was added at a final concentration of 100 ng/mL in cell culture plate. The cells were observed and analyzed using an Olympus confocal microscope.
Statistical analysis
The data of each sample were analyzed in 3 replicates. Statistical analyses were performed using the SPSS16.0 software. Comparisons between groups were analyzed using the t-test and χ 2 test.
Results
The oxidative stress of β3-activator tert-butyl hydroperoxide on BM-MSCs
In the case of oxidative stress, cell mitochondria produce large amounts of reactive oxygen species (ROS), then ROS would induce physiological dysfunction in various cancer cells. Tert-butyl hydroperoxide, 1 β3-activator, is widely used as a stimulator that promotes ROS and then leads to oxidative damage in cancer cells (Piret and others 2005; Slamenova and others 2013). To investigate the best oxidative stress time for BM-MSCs, relative BM-MSCs activity analysis indicates that 1.5 mmol/L tert-butyl hydroperoxide exerts the best effect at 24 h by MTT assays in the model group combined with β3-activator; however, when si-LKB1 was transfected, the relative BM-MSCs activity was significantly decreased in the model group combined with β3-activator and si-LKB1 (Fig. 1A). Meanwhile, we measured the level of ROS in various groups, and the results confirmed that ROS presented in an opposite trend of MTT assay while rapamycin could not reverse the phenomenon in the model group combined with β3-activator and si-LKB1 (Fig. 1B). These results demonstrated that tert-butyl hydroperoxide exerted a great ROS damage to BM-MSCs and it really increased the content of ROS to a large extent.

The oxidative stress of β3-activator tert-butyl hydroperoxide on model of BM-MSCs.
LKB1-siRNA inhibits the transformation of BM-MSCs into adipocytes
As we know, AMPK has a certain effect on the differentiation of BM-MSCs into white fat or brown fat under oxidative stress. Interestingly, LKB1 is involved in the regulation of energy balance by AMPK, and induces the synthesis of fatty acid synthetic proteins FABP2 and adiponectin (Fogarty and others 2016; Wang and others 2019). To explore the link between LKB1 and the transformation of BM-MSCs in the environment of oxidative stress, we analyzed the differentiation ability of BM-MSCs that were treated with LKB1-siRNA by Oil Red O staining (Fig. 2A, B). The results indicate that knockdown LKB1 significantly inhibits the number of adipocytes transformed by BM-MSCs under oxidative stress (P < 0.05).
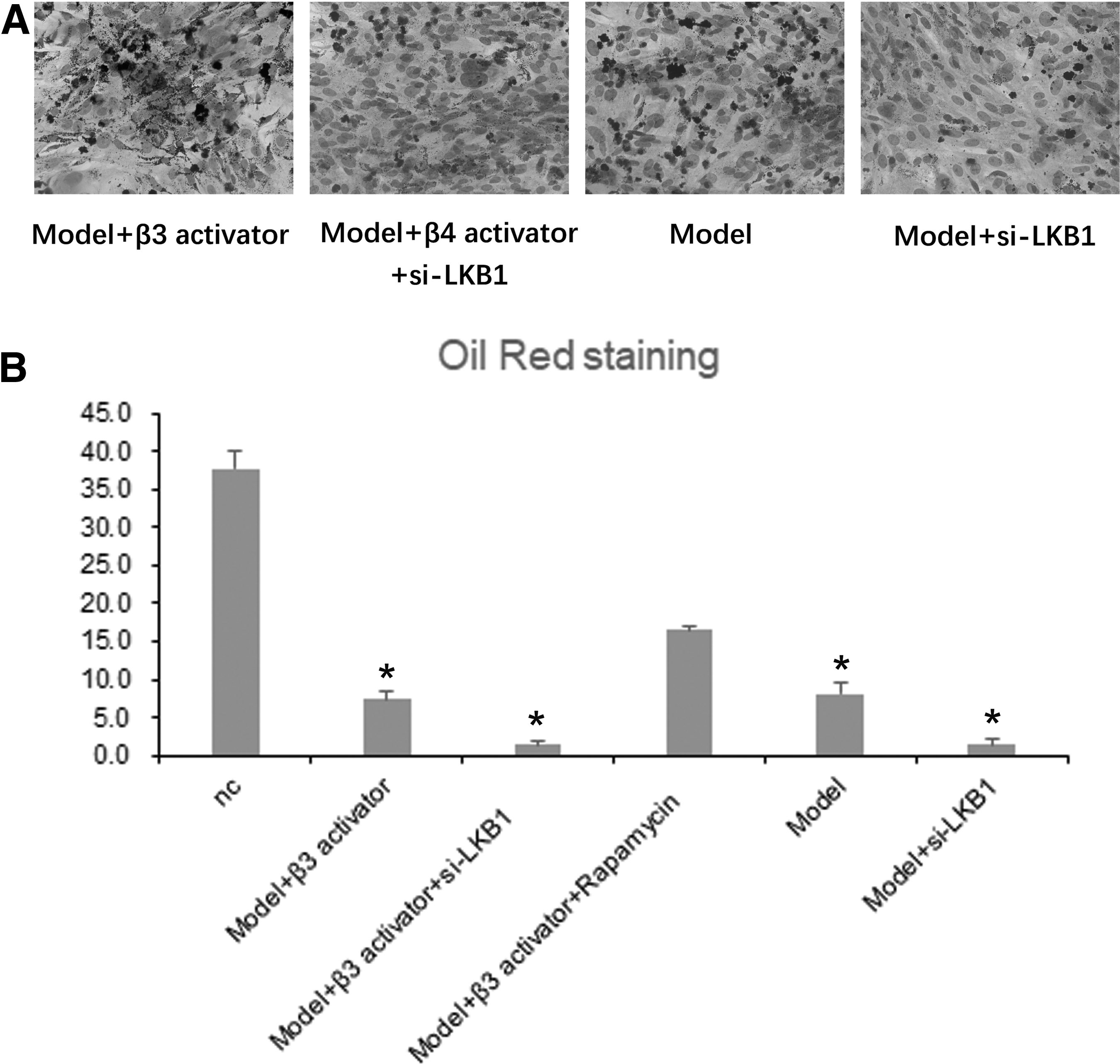
LKB1-siRNA inhibits the transformation of BM-MSCs into adipocytes.
Recent reports have proved that β3 adrenergic receptor agonists induced the transformation of WAT to a like brown adipose tissue in the specific situations (Danysz and others 2018). Our experimental results also proved the same conclusion; correspondingly, β4 adrenergic receptor agonists restored the transformation ability of BMC cell caused by LKB1-siRNA (Fig. 2B).
LKB1 induces the synthesis of fatty acid synthetic proteins by AMPK pathway
We have demonstrated that LKB1 enhances the transformation ability of BM-MSCs under oxidative stress, we next explore whether this effect is AMPK dependent. We analyze the protein of PPARγ and C/EBPα that is regulated by AMPK, the expression of these in the oxidative stress group is higher than that of the normal group obviously. Instead, knocking down LKB1 reduces the trend caused by oxidative stress. When β3 adrenergic receptor agonists are added, the expression levels of these proteins are again increased. This means that LKB1 increased the activity of AMPK pathway, inducing the overexpression of PPARγ and C/EBPα. Accordingly, LKB1-siRNA inhibits the expression of fat synthesis gene by decreasing the transformation of BM-MSCs (Fig. 3A, B). The results indicate that the effect of the differentiation of BM-MSCs by LKB1 depends on the AMPK pathway.

LKB1 induces the synthesis of fatty acid synthetic proteins by AMPK pathway.
LKB1 induces differentiation of adipocytes into brown adipocytes
Then we observed the morphological changes of BM-MSCs, electron microscopy scan reveals that β3 adrenergic receptor agonists can induce differentiation of adipocytes into brown adipocytes. Knocking down LKB1 inhibits the transformation effect and reduces the number of brown fat cells (Fig. 4A, B). The experimental results are consistent with the mentioned conclusions.

LKB1 induces differentiation of adipocytes into brown adipocytes. The morphological changes were observed under electron microscopy scan. When compared with model group, β3-activator induced the adipogenesis of BM-MSCs.
LKB1 and AMPK are located in the cytoplasm of BM-MSCs
The mentioned experiments demonstrate that the induction of BM-MSCs by LKB1 is highly correlated with AMPK. Immunofluorescence staining of the 2 proteins revealed that LKB1 and AMPK are located in the cytoplasm of BM-MSCs and partially overlap. In addition, oxidative stress increases the expression of LKB1 and AMPK simultaneously (Fig. 5A, B). The results of this experiment corroborate our mentioned views.
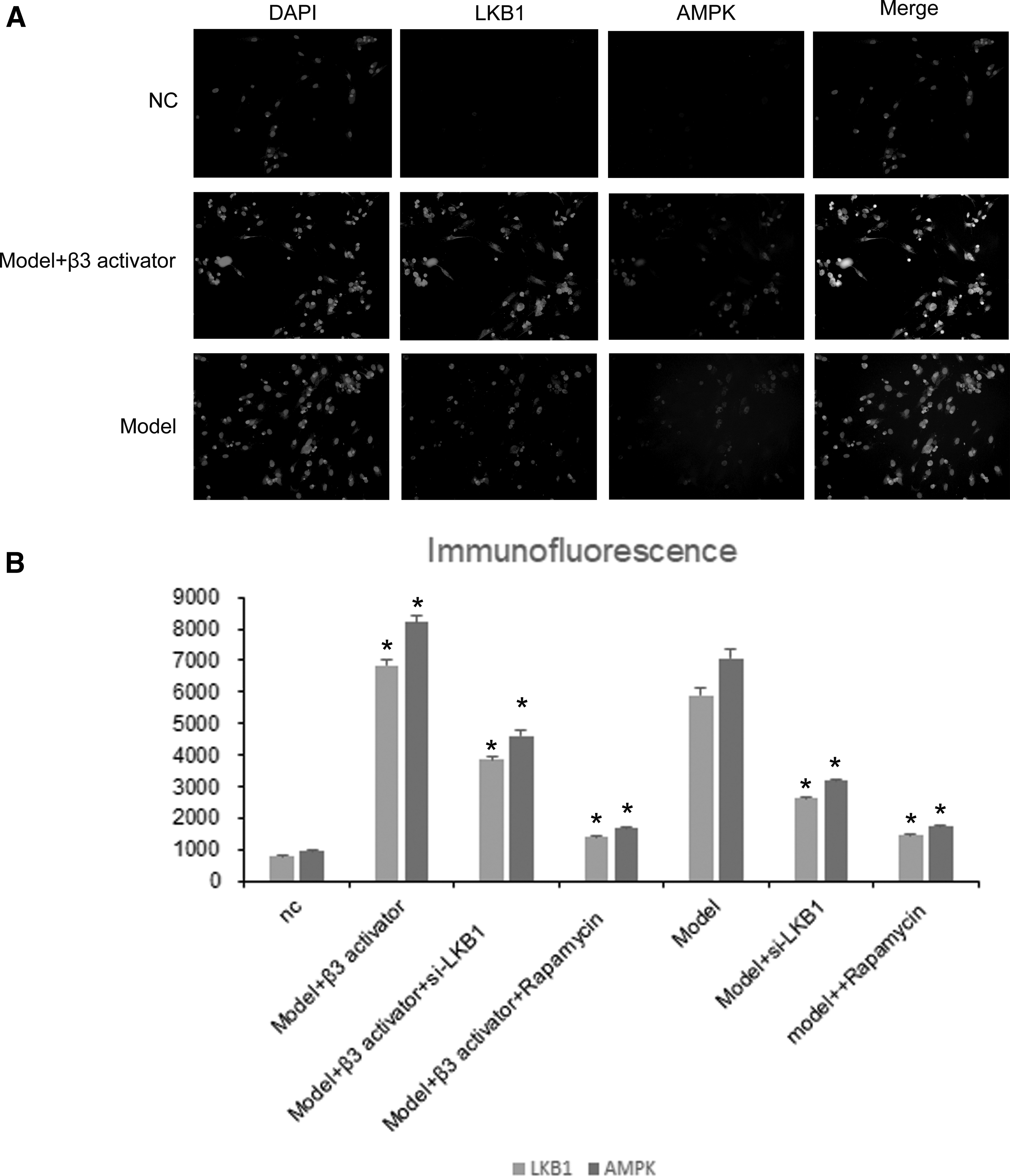
LKB1 and AMPK are located in the cytoplasm of BM-MSCs.
Discussion
MSCs are differentiated from hematopoietic stem cells in the bone marrow, the injured tissue-derived signals could promote the differentiation of resting MSCs (Bara and others 2014). The AMPK pathway is involved in a variety of important life processes, including regulating the differentiation of BM-MSCs (Xia and others 2015; Zhang and others 2016). In addition, oxidative stress promotes phosphorylation of AMPK and activates the AMPK pathway (Kar and others 2015). Previous studies have shown that AMPK regulates energy balance and induces the synthesis of fatty acid synthetic proteins FABP2 and adiponectin. Interestingly, when the energy is insufficient, AMPK promotes energy release by inducing lipolysis (Cao and others 1991). Our study demonstrates that oxidative stress promotes the expression of AMPK and enhances the regulation of AMPK on the transformation of BM-MSCs into adipocytes. This means that we can artificially regulate the activation of the APMK pathway and induce the differentiation of BM-MSCs.
LKB1 has kinase activity and can regulate activity by promoting phosphorylation of AMPK. Studies have shown that the LKB1/AMPK pathway played a role in some life processes such as vasoconstriction, systemic ischemic response, and cell cycle (Fogarty and others 2016; Kim and others 2018; Majd and others 2018). In nonsmall cell lung cancer and in cervical cancer, research suggests that LKB1 mutation is correlated with oxidative stress-inducing therapies (Zulato and others 2018). But the relationship between LKB1 and BM-MSCs has not been reported. We found that knocking down LKB1 significantly attenuated the conversion of BM-MSCs to adipocytes under oxidative stress, and we demonstrated that the regulation was AMPK dependent. More interestingly, oxidative stress can simultaneously promote the expression of LKB1 and AMPK. This phenomenon increases the regulation of LKB1 on transformation of BM-MSCs.
Conclusions
In summary, the study showed that LKB1-siRNA significantly inhibits the transformation of BM-MSCs into adipocytes under oxidative stress, and the regulation still relies on the classic LKB1/AMPK pathway. In addition, we also demonstrated that β3 receptor agonists could reverse the effect of knockdown of LKB1 on transformation of BM-MSCs. The proposed LKB1/AMPK/BM-MSCs pathway is significant for the understanding of transformation of BM-MSCs and provides the basis for the utilization of BM-MSCs.
Footnotes
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This study was funded by the Discipline Construction Project of Pudong New Area Commission of Health and Family Planning (Grant No. PWYts2018-03).