
Abstract
Human immunodeficiency virus-tuberculosis (HIV-TB) co-infection poses a challenge to the immunologists in developing new diagnostic and therapeutic tools. Mechanisms behind the breakdown of the immune defense of the co-infected individual are poorly known. Numerous studies in HIV alone have revealed the role of PD1, TAP, and IL-10, but not in co-infection. The interaction of the 2 distinct bugs, which is resulting in domination over the host immune system, is still a lacuna. Hence, we aimed to portray functions of IL-10, TAP, and PD1 molecules in HIV-TB co-infection. Co-culture cells challenged with γ-irradiated M.Tb under various conditions resulted in high interleukin (IL)-10 secretion and high percentage of PD1 expression on CD8 T cells, which might be due to defective antigen presentation of TAP on dendritic cells and macrophages. Herein our observations provide an insight into the escape mechanisms by M.Tb in HIV-infected individuals from the host immune responses leading to TB co-infection.
Introduction
Human immunodeficiency virus-tuberculosis (HIV-TB) co-infection poses a challenge to the immunologists in developing new diagnostic and therapeutic tools. Mechanisms behind the breakdown of the immune defense of the co-infected individual are poorly known. In developing countries, TB is accounting for about 26% of Acquired Immune Deficiency Syndrome (AIDS)-related deaths (Collins and others 2002; Getahun and others 2010). Mechanisms of interaction of the 2 pathogens still have many gaps that need to be filled to develop preventive measures against the 2 diseases. Dysregulation of the immune system and alterations in the cytokine profile are common in HIV infection (Hertoghe and others 2000); on the other hand, activation of Th1 lymphocytes and macrophage is crucial in producing interferon-γ (IFN-γ) for defense against mycobacteria (Lienhardt and others 2002).
Transport of peptides from proteasome after proteolytic degradation is done by ATP-dependent family of transporters called TAP proteins. The heterodimer TAP translocates peptides from the cytosol across the membrane into the lumen of endoplasmic reticulum (ER), subsequently loaded on to class I major histocompatibility complex (MHC) molecules and presented to class I primary human B lympho-restricted CD8/cytotoxic T lymphocytes (CTLs) (Ljunggren and others 1990; Schumacher and others 1991). Interleukin-10 (IL-10) is produced mainly by TH2 cells; it has a unique broad immunomodulatory effect, especially in suppressing cell-mediated immunity through downregulating proinflammatory cytokines, co-stimulatory molecules, as well as MHC class II proteins. IL-10 gene has been implicated in HIV-1 infection and pathogenesis in numerous studies (Erikstrup and others 2007; Chatterjee and others 2009; Naicker and others 2009; Oleksyk and others 2009). A study by Zeidler and others reported that IL-10 is capable of modulating the expression of TAP1 levels in different B cells, but is not capable of abrogating TAP1 expression totally (Zeidler and others 1997).
Our previous studies demonstrated an association of Transporter associated with antigen processing-1 (TAP1) and IL-10 gene polymorphism with susceptibility for HIV-TB co-infection (Sunder and others 2011; Ramaseri Sunder and others 2012). IL-10 also impedes macrophage co-stimulatory activity, perhaps through restraint of the expression of MHC class II (de Waal Malefyt 1991). The molecular mechanism by which HIV infection may induce differential IL-10 production by T cells and monocytes is not well understood. High IL-10 levels can induce immunosuppression that either can affect transcriptional activation of the TAP1 gene or the turnover of rate of transcription (Zeidler and others 1997). Therefore, we hypothesize that high IL-10 levels could affect crucial TAP expression.
CD8+ T cells play a crucial role in control of retroviral infections. Programmed cell death-1 (PD1), an apoptotic marker on the cell surface, is a negative regulator of T cells. Recent studies revealed the critical role of PD1 in regulation of viral-specific T cells (Petrovas and others 2006). Upregulation of PD1 was also associated with reduced perforin and IFN-γ production, as well as decreased HIV-specific effector memory CD8+ T cell proliferation (Zhang and others 2007). A correlation between PD1 expression on effector CD8+ T cells and the frequency of IL-10+ suppressor CD8+ T cells in HIV infection (Elrefaei and others 2008) identified by a recent study is one of the supporting evidences to choose these markers. CD38/PD1 phenotype clearly distinguishes HIV-specific CD8 T cell responses between controllers and progressors (Vollbrecht and others 2010). Expression of PD1 is regulated not only by T cell activation status (Jurado and others 2008) but also by HIV peptides (Muthumani and others 2008). Besides, PD1 signaling inhibits T cell effector functions during TB (Jurado and others 2008) and HIV (Muthumani and others 2008). Jurado and others (2012) observed that higher basal levels of PD1 on T cells from HIV-positive subjects may induce dysfunctional M.tb-specific T cells, reflected in lower levels of IFN-γ production and proliferation in vitro against the antigen. These findings taken together, we aimed to check whether high CD38+/PD1+ and HLA co-expressions on CD8 cells leads to development of TB in HIV+ individuals as a co-infection.
Materials and Methods
Patient population
Immunophenotyping studies in peripheral blood
Sixteen HIV-seropositive patients with CD4 counts >350 cells/μL, of age group 32.45 ± 10.97 [mean ± standard deviation (SD)] without any clinical symptoms of TB and no previous history or family history of TB, 10 newly diagnosed TB patients who were seronegative for HIV test, of age group 34.57 ± 15.51 (mean ± SD), and 11 newly diagnosed HIV-TB co-infected patients, of age group 34.41 ± 8.43 (mean ± SD) attending the clinic at Blue Peter Public Health and Research Centre, LEPRA Society, Hyderabad, were enrolled. The control group comprised 10 HIV-seronegative, asymptomatic healthy volunteers, irrespective of their PPD status, without previous history or family history of TB of age group 28.74 ± 7.70 (mean ± SD) from different areas around the clinic.
Functional studies
Three HIV-seropositive, latent TB infection-positive (LTBI+), newly diagnosed patients whose CD4 counts were >350 cells/μL, of age group 28.33 ± 2.89 (mean ± SD) and without any clinical symptoms of TB with no previous history or family history of TB attending the clinic at Blue Peter Public Health and Research Centre, LEPRA Society, Hyderabad, were included in the study. QuantiFERON Gold In-Tube assay was performed to identify LTBI+. Furthermore, 3 HIV-seronegative, asymptomatic healthy volunteers without previous history or family history of TB and of age group 35.33 ± 9.45 (mean ± SD), from different areas around the clinic, were also included as controls.
Study was approved by institutional ethics committee (10/2010). Peripheral blood was collected after obtaining an informed consent from all of them.
Monocyte and T cell co-culture
Isolation of peripheral blood mononuclear cells (PBMC), CD14+ monocytes, and CD3+ T cells: PBMC were isolated by differential centrifugation over Histopaque (Sigma, Ayrshire, UK) in 1:3 ratio and centrifuged at 1800 RPM for 30 min. White interlayer of cells were collected and washed twice with 1 × Hanks buffer salt solution (Sigma, Paiseley, UK) and purified CD14+ and CD3+ cells were isolated by positive selection with magnetic beads conjugated to anti-CD14 and anti-CD3, respectively (Miltenyi Biotech, Auburn, CA). The positive cells were 91% for CD14 and 94% for CD3 as measured by flow cytometry. Cells were resuspended in RPMI 1640 (Sigma, Ayrshire, UK) complete medium with 10% Human AB serum (Invitrogen, Carlsbad, CA), L-glutamine, and Penicillin/Streptomycin (Sigma, St. Louis, MO). Viability and cell count were evaluated with Trypan blue (Sigma, St. Louis, MO).
Cell culture conditions: CD14+ monocytes (0.5 × 106 cells/mL) and CD3+ T cells (1 × 106 cells/mL) (1:2 ratio) were plated in flat-bottomed 24-well plates (Corning Incorporated, NY) in 1000 μL of complete medium (Sigma, Ayrshire, UK) in the presence or absence of 10 μg/mL γ-irradiated M. TB or 10 μg/mL Ag85A (BEI resources). In some experiments, along with γ-irradiated M. TB recombinant IL-10 (BD Pharmingen, San Diego, CA), 10 μg/mL neutralizing antibodies to IL-10 (BD Pharmingen), 10 μg/mL neutralizing antibodies to IFN-γ (BD Pharmingen), 10 μg/mL neutralizing antibodies to PD1 (R&D Systems, Minneapolis, MN), or 10 μg/mL isotype control mouse IgG1 (R&D Systems) were also added to the cells and were cultured for 96 h.
TAP expression
Total RNA was extracted from cultured cells using Tri-Reagent (Sigma, St. Louis, MO). RNA content was measured in a spectrophotometer at 260 nm (Nanodrop-1000; Thermo, Wilmington, DE). Total RNA was reverse transcribed using Superscript-III first-strand synthesis system (Invitrogen). Five hundred nanograms of total RNA was transformed into cDNA in a final volume of 20 mL. The expression of TAP1 and TAP2 gene transcript was amplified on Mini Opticon Detector for real-time PCR (CFB-3120; Bio-Rad Laboratories, Inc.) using the SYBR Green PCR kit (Qiagen, Hilden, Germany). The forward and reverse primers for TAP 1 and 2 genes (Kaczmarek and others 2007) had amplicon size of 301 and 185 bp, respectively. Primers for HGPRT were forward, 5′-GCCATCAATGACCCCTTCATT-3′, and reverse, 5′-TTGACGGTGCCATGGAATTT-3′. Real-time PCR was performed in duplicates with the conditions 94°C for 10 min, and 45 cycles of 94°C for 15 s, 60°C for TAP1, and 62°C for TAP2, respectively, for 60 s and 72°C for 30 s, followed by a melting curve starting with 60°C rising to 94°C at 0.5 per 2 s. All samples were normalized to the amount of HGPRT transcript present in each sample. Relative expression was calculated as 2ΔCT stimulated/2ΔCT control, where ΔCT = CTTAP − CTHGPRT.
Cytokine assay
Measurement of IL-10 and IFN-γ concentrations: IL-10 and IFN-γ levels were measured by ELISA Ready-SET-Go kit (eBiosciences, San Diego, CA) following manufacturer's instructions. For measurement of IL-10 and IFN-γ, cell-free culture supernatants were collected after 96 h of culture. Supernatants were stored at −80°C until cytokine concentrations were measured.
Cytofluorometric analysis
Antibodies used for staining
PD1-FITC, CD8-PE, CD38-PerCP, CD4-APC, CD14-FITC, HLA-DR-PE, CD163-PerCP, and HLA-A, B,C-APC
Cells from peripheral blood were surface stained for PD1/CD8/CD38/CD4
The cultured cells were washed and surface staining was done for 3 different panels:
(a) PD1/CD8/CD38/CD4
(b) CD14/CD8/CD38/CD4
(c) CD14/HLA-DR/CD163/HLA-A, B,C
CD4 (Cat. No. 555349) and CD8 (Cat. No. 340046) from BD Biosciences, CA,; HLA-DR (Cat. No. 307605), HLA-A,B,C (Cat. No. 311409), PD1 (Cat. No. 329904), CD38 (Cat. No. 303520), CD14 (Cat. No. 325603), and CD163 (Cat. No. 333607) from Biolegend, San Diego, CA.
Samples were acquired on FACS Calibur (BD Biosciences) and data were analyzed using Cell Quest Pro Software (Version 5.2.1). PD1/CD8/CD38/CD4 cells were gated from lymphocytes with 25,000 event acquisition and cells of other 2 panels were gated from lymphocytes and monocytes with 25,000 event acquisition on dot plot.
Statistical analysis
Results are shown as mean ± standard error after using GraphPad Prism version 5.00 (San Diego, CA). Comparisons between groups were done by an unpaired t-test and 1-way analysis of variance with Tukey's multiple comparisons test as appropriate. P < 0.05 was considered statistically significant. Microplate Manager Software version 6 (MPM6; Bio-Rad Laboratories, Veenendaal, Netherlands) was used to convert optical density into concentrations.
Results
Influence of mycobacterial peptide-specific stimulation on TAP expression
To determine whether mycobacterial peptide stimulation regulates the expression of TAP protein, the CD3+/CD14+ co-culture of HIV and control groups was challenged with and without Ag85A, γ-irradiated M.tb, γ-irradiated M.tb + recombinant IL-10, γ-irradiated M.tb +neutralizing antibodies to IL-10, γ-irradiated M.tb +neutralizing antibodies to IFN-γ, and γ-irradiated M.tb + neutralizing antibodies to PD1 as outlined in materials and methods.
The regulation of TAP1 and TAP2 expression on stimulation with Ag85A, γ-irradiated M. tb, γ-irradiated M. tb + recombinant IL-10, and γ-irradiated M. tb +neutralizing antibodies to IFN-γ, was almost similar in HIV and control groups.
Blocking of IL-10 increased the expression of TAP1 mRNA in patient group only. The stimulation with γ-irradiated M. tb+ neutralizing antibodies of IL-10 regulated the expression of TAP1 in the HIV group. The TAP1 mRNA expression was high in the HIV group compared to the control group.
Blocking of PD1 increased the expression of TAP2 mRNA. The stimulation with γ-irradiated M. tb+ neutralizing antibodies of PD1 regulated the expression of TAP2 in the HIV group. The TAP2 mRNA expression was significantly high in the HIV group compared to the control group (Fig. 1 and Table 1).

Relative mRNA expression of TAP1 (healthy controls and patients) and TAP2 (healthy controls and patients) in stimulated cells compared with its corresponding unstimulated cell in different study groups. Each line represents the mean fold change. P < 0.05 is considered significant. *P < 0.05, **P < 0.001. TAP1, transporter associated with antigen processing-1.
Relative mRNA expression of TAP1 and TAP2
Significant P-values are highlighted in bold.
IFN-γ, interferon-γ; SEM, standard error of the mean; TAP1, transporter associated with antigen processing-1.
Activation of monocytes on mycobacterial peptide-specific stimulation
Γ-irradiated M.tb-specific stimulation presented significantly low (P < 0.05) percentage of CD14+/CD38+ cells in the HIV group compared to the control group. This observation might suggest the lack of activation of monocytes on stimulation with certain mycobacterial peptides (Supplementary Fig. S1a).
Expression of monocyte-derived macrophages (CD14+/CD163+) on mycobacterial peptide-specific stimulation
The percentage of CD14+/CD163+ cells was significantly low (P < 0.05) when stimulated with γ-irradiated M.tb+ rIL-10 in the HIV group compared to the control group. Demonstrating the high levels of IL-10 may interfere with the activation of macrophages (Supplementary Fig. 1b).
Antigen presentation (CD14+163+/HLA-DR+/HLA-A, B, C+) on mycobacterial peptide-specific stimulation
On stimulation with γ-irradiated M.tb+ nIFN-γ and γ-irradiated M.tb+ rIL-10, percent of HLA-DR+/HLA-A,B,C+ on monocyte-derived macrophages was significantly low (P < 0.05) in the HIV group compared to the control group These observations might demonstrate lack of antigen presentation in the absence of IFN-γ and presence of IL-10 (Supplementary Fig. 1c).
T cell responses to mycobacterial peptide specific stimulation
Our observations in peripheral blood presented the percentage of CD4+ cells was significantly low in HIV and HIV-TB groups compared to that of TB and control, while CD8+ cells had significantly high percentage in HIV and HIV-TB groups compared to that of TB and control groups (Table 2 and Fig. 2). Hence, we determined the response of CD8+ cells toward antigen presentation by monocyte-derived macrophages in CD3+CD14+ co-culture challenged with mycobacterial peptides.
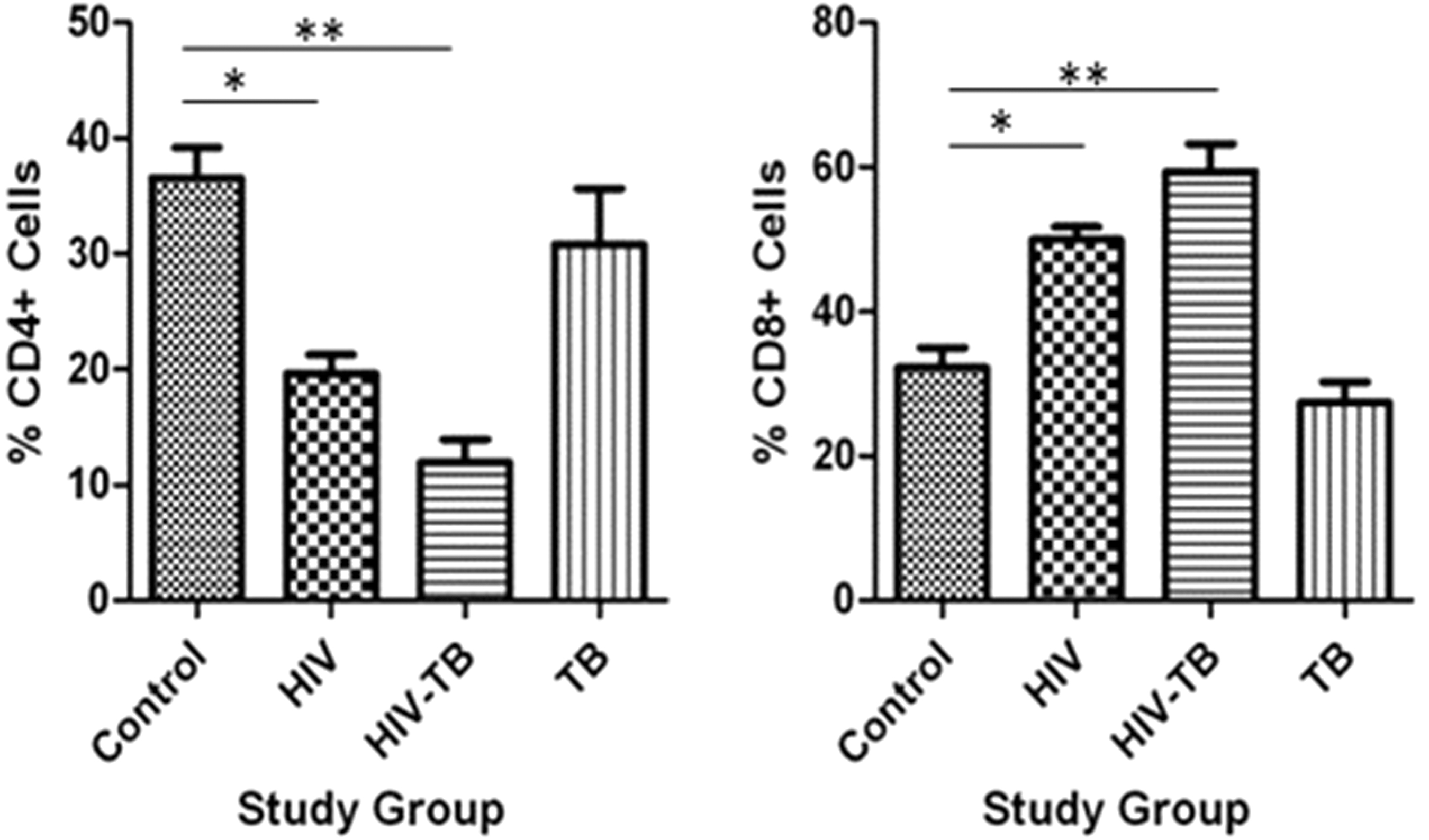
Percentage CD4+ and CD8+ expression in different study groups: mean ± SEM expression in control, HIV, HIV-TB, and TB study groups is shown. *P < 0.05, **P < 0.001 with mean difference and 95% class intervals. HIV-TB, human immunodeficiency virus-tuberculosis; SEM, standard error of the mean.
% Expression of CD4+ (TH) and CD8+ (TCTL) Cells in Different Study Groups
CTL, cytotoxic T lymphocyte; HIV-TB, human immunodeficiency virus-tuberculosis.
The significantly high percentage of CD8+ cells on stimulation with Ag85A and γ-irradiated M.Tb in HIV group demonstrated active CTL response toward mycobacterial antigen in HIV + individuals (Supplementary Fig. S2a), while HIV+TB group has similar trend as that of HIV group (data not shown).
Activation of CD8+ cells on mycobacterial peptide-specific stimulation
As we demonstrated, the percentage of CD8+ was significantly high in peripheral blood as well as after stimulation with specific mycobacterial antigens. We wanted to determine the activation of these cells by looking at the expression of CD38+ on CD8+ cells.
The percentage of CD38+ on lymphocytes in peripheral blood was significantly high in HIV-TB groups when compared to control and TB (Table 3 and Fig. 3). While the percentage of CD38+ on CD8+ cells in peripheral blood was significantly high in HIV and HIV-TB groups compared to that of control and TB groups (Table 3 and Fig. 4), a similar trend was observed on CD4+ cells as well.
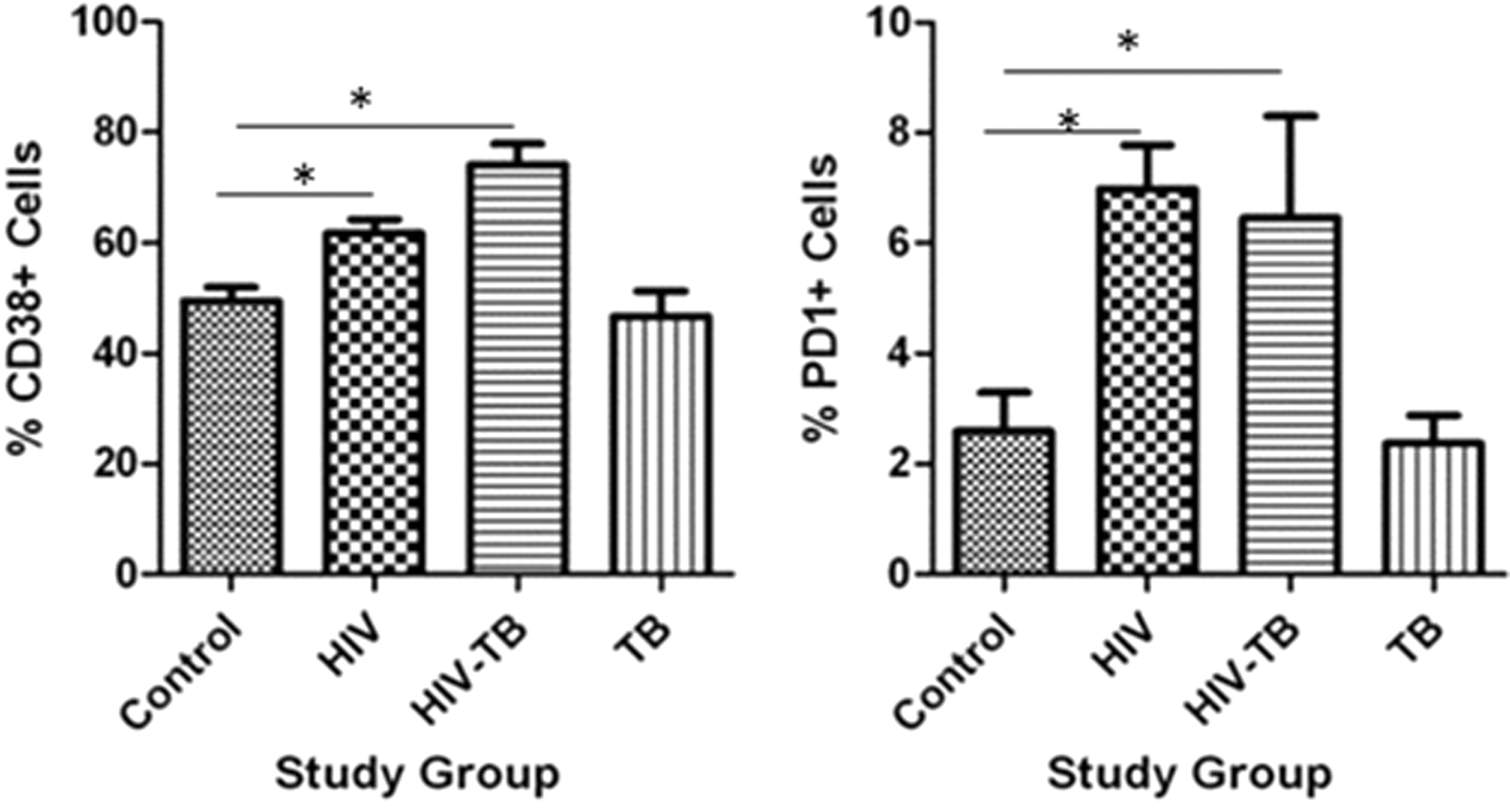
Percentage CD38+ and PD1+ expression on lymphocytes in different study groups: mean ± SEM expression in control, HIV, HIV-TB, and TB study groups is shown. *P < 0.05 with mean difference and 95% class intervals.
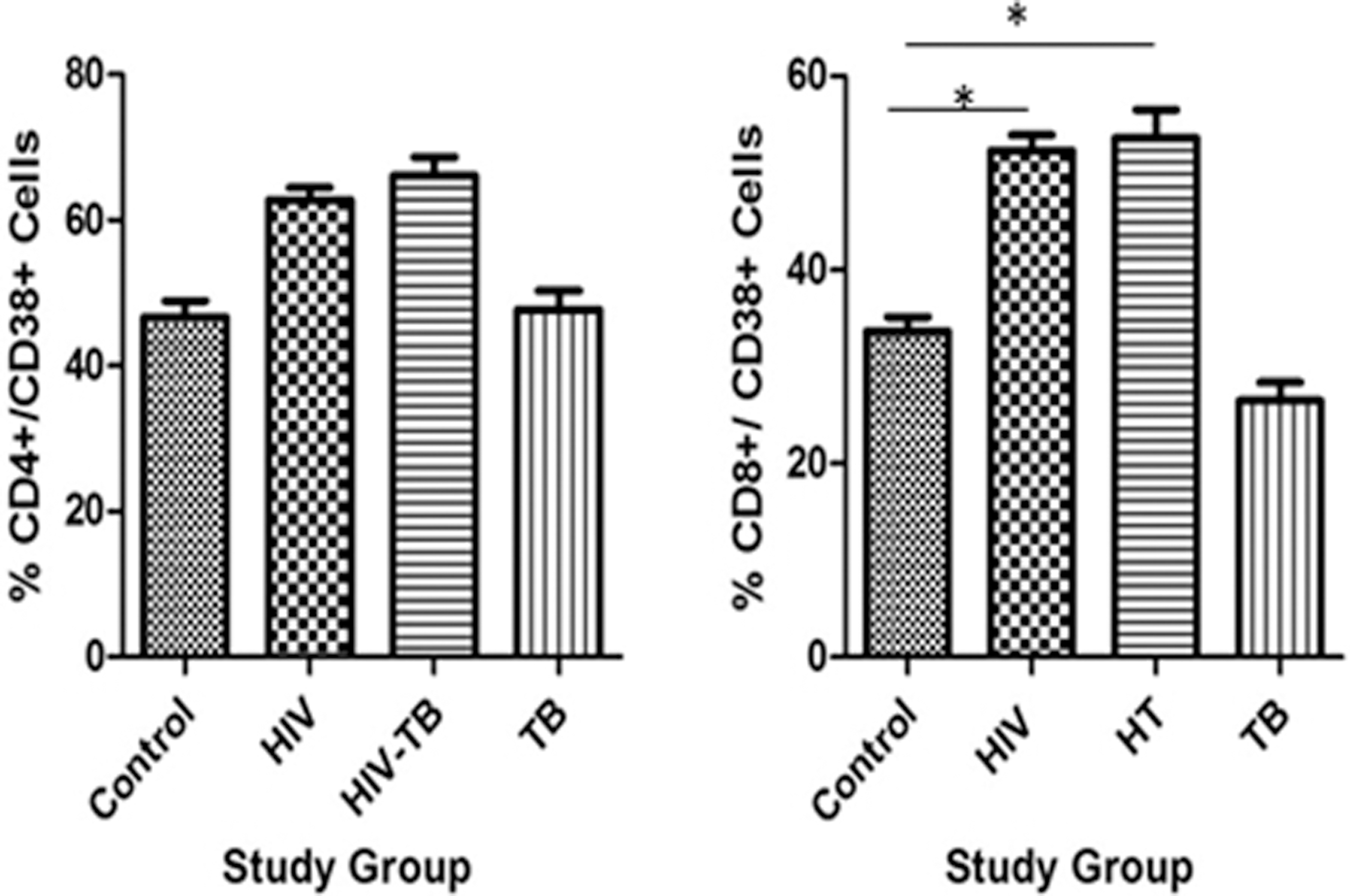
Percentage CD38+/CD4+ and CD38+/CD8+ expression in different study groups: mean ± SEM expression in control, HIV, HIV-TBc, and TB study groups is shown. *P < 0.05 with mean difference and 95% class intervals.
% Expression of 38+/CD4+and 38+/CD8+ Cells in Different Study Groups
On stimulation with γ-irradiated M.tb and γ-irradiated M.tb+ nPD1, the CD38+ levels decreased in patients (with mean ± standard error of the mean 5.445 ± 1.501; 6.823 ± 2.818) compared to controls (19.28 ± 2.978; 21.37 ± 1.120), while the percentage of CD38+ on CD8+ was similar in all stimulated conditions. Activation of cells under stimulated conditions with γ-irradiated M. tb and blocking with nPD1 might be poor.
Apoptosis of CD8+ cells on mycobacterial peptide-specific stimulation
To determine the apoptosis (cell death) of CD8+ cells, we checked for the expression of PD1 marker on lymphocytes as well as on CD8+ cells in peripheral blood and stimulated cultured cells.
In peripheral blood of HIV group, PD1 percentage on lymphocytes was significantly high compared to control and TB group (Table 4 and Fig. 3), while the percentage of PD1 on CD8+ cells was significantly high in HIV and HIV-TB groups compared to control and TB (Table 5 and Fig. 5).
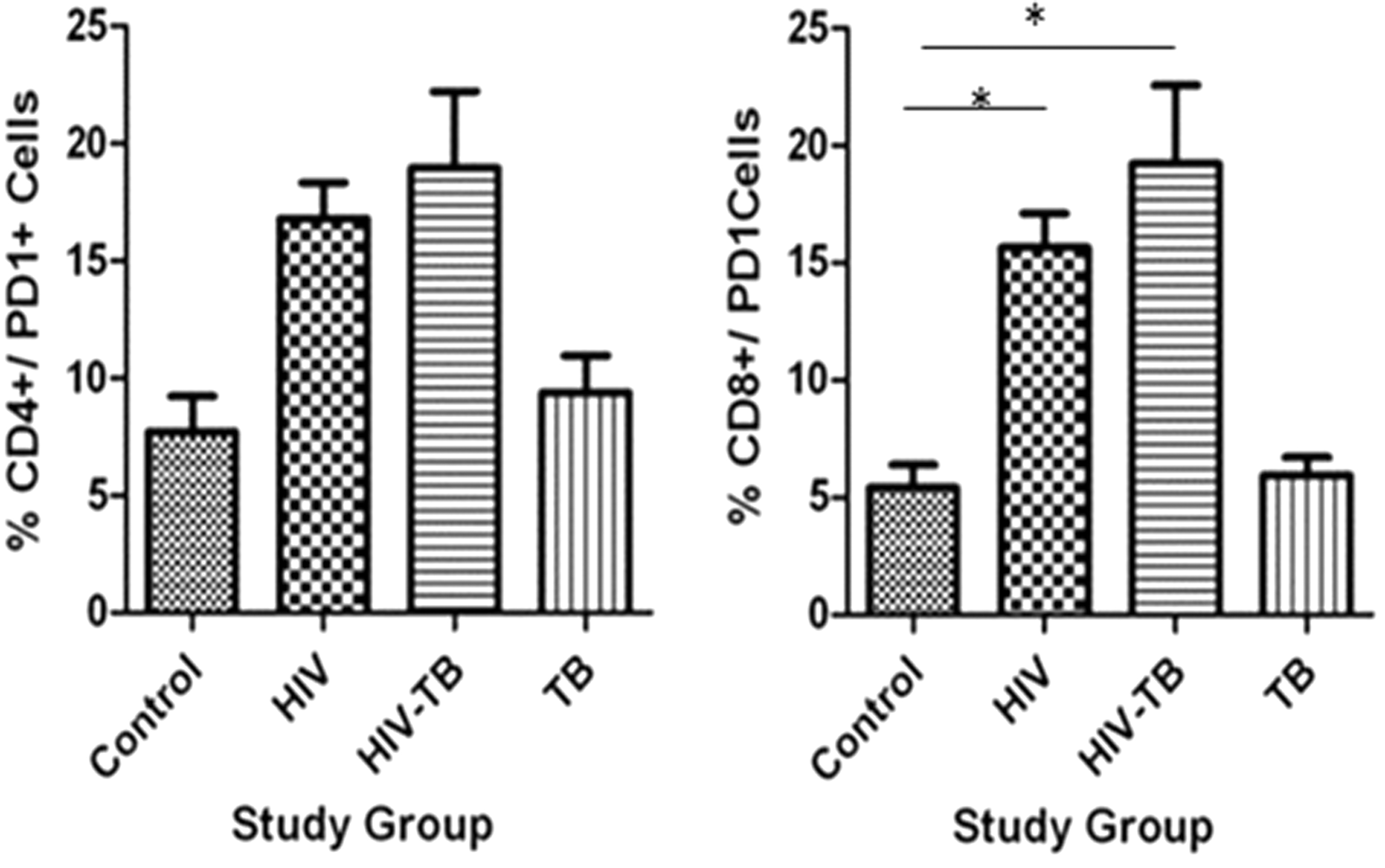
Percentage PD1+/CD4+ and PD1+/CD8+ expression in different study groups: mean ± SEM expression in control, HIV, HIV-TB, and TB study groups is shown. *P < 0.05 with mean difference and 95% class intervals.
% Expression of CD38+ and PD1+ on Lymphocytes in Different Study Groups
% Expression of PD1+/CD4+ and PD1+/CD8+ Cells in Different Study Groups
On stimulation with γ-irradiated M.tb+ nIFN-γ, γ-irradiated M.tb+ nIL-10, and γ-irradiated M.Tb+ rIL-10 the percentage of PD1 on CD8+ cells was significantly high in HIV (61.99 ± 7.484; 64.79 ± 10.9; and 70.51 ± 8.176) group compared to control (14.53 ± 5.061; 11.40 ± 1.648; and 26.54 ± 10.29).
Demonstrating that blocking of IFN-γ lead to increased cell death, the percentage of 38+/PD1+ on CD8+ cells in peripheral blood was significantly high in HIV and HIV-TB group compared to control group; also, the percentage of 38+/PD1+ on CD8+ cells was significantly high in HIV-TB group compared to HIV and TB groups. This might demonstrate the loss of CTL responses in HIV and HIV-TB groups (Table 6 and Fig. 6).

Percentage CD38+/PD1+/CD4+ and CD38+/PD1+/CD8+ expression in different study groups: mean ± SEM expression in control, HIV, HIV-TB, and TB study groups is shown. *P < 0.05, **P < 0.001 with mean difference and 95% class intervals.
% Expression of CD38+/PD1+/CD4+and CD38+/PD1+/CD8+ Cells in Different Study Groups
On stimulation with γ-irradiated M.tb, γ-irradiated M.tb+ nIFN-γ, γ-irradiated M.tb+ nIL-10, and γ-irradiated M.tb+ rIL-10, the percentage of CD8+/CD38+/PD1+ cells was significantly high (P < 0.05) in HIV group (42.67 ± 5.568; 41.02 ± 4.460; 45.33 ± 6.615; and 46.95 ± 3.455) compared to control group (18.62 ± 6.891; 9.633 ± 4.812; 7.633 ± 1.429; and 14.64 ± 3.819).
Expression of IFN-γ on mycobacterial peptide-specific stimulation
The In CD3+/CD14+ co-cultures stimulated with γ-irradiated M.tb+ neutralizing antibodies of IL-10 and γ-irradiated M.tb+ neutralizing antibodies of PD-stimulated cultures of HIV group have low level (P < 0.05) of IFN-γ compared to control group, show that the blocking of IL-10 and PD1 will not enhance the expression of proinflammatory cytokine IFN-γ (Fig. 7).
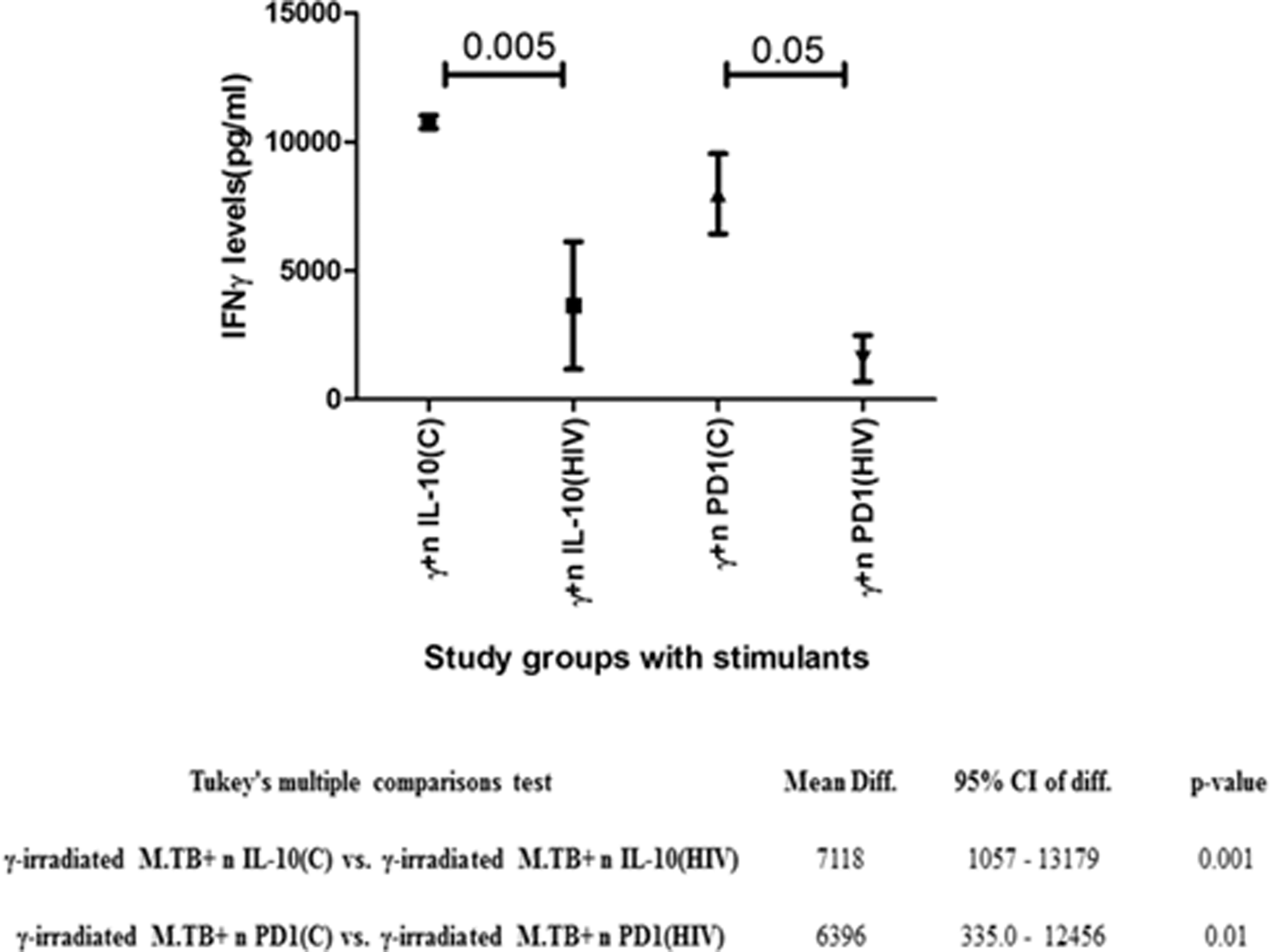
IFN-γ (pg/mL) levels in stimulated CD3+ CD14+ co-cultures: graph represents mean and error bar. Significance was calculated using t-test: P < 0.05 is considered significant. IFN-γ, interferon-γ.
Discussion
The CD8+ T cell priming resultant of MHC class I pathway is usually associated with presentation of endogenous antigens. The cytosolic antigens undergo degradation within proteasome, forming peptides, which are transferred from ER through TAP-mediated mechanism (Goldszmid and others 2007). In our previous studies, we reported that the polymorphism in TAP1 gene might cause susceptibility to TB in HIV-positive individuals (Ramaseri Sunder and others 2012). Defect in antigen-presenting machinery can cause cells deficient with peptide transport and further leads to lack of antigen presentation (Qin and others 2002; Urosevic and others 2003). We hypothesized that exploitation of antigen presentation pathway by M.tb might be one of the escape mechanisms against host immune responses in HIV-infected individuals.
In this study, the TAP expression in both HIV individuals and healthy controls with TB latency did not vary when challenged with mycobacterial peptides. We reported earlier that high IL-10 levels might hinder TAP expression allowing the survival of M.tb in HIV-positive individuals leading to co-infection (Ramaseri Sunder and others 2012). It is well established that IL-10 neutralizes IFN-γ signaling and the activity of antigen-presenting cells (APCs), allowing the proliferation of intracellular microbes (Kumar and Creery 2000; Urosevic and Dummer2002; Cheung and others 2005; Li and others 2005; Lee and others 2007; Chan and others 2009, 2010). HIV-positive individuals with TB at the latent stage are immune compromised with low levels of IFN-γ. Our observation that TAP expression is elevated in IL-10-suppressed conditions indicates that high IL-10 levels block TAP proteins.
Furthermore, there are also reports stating that IL-10 dampens APC and T cell interactions, resulting in less efficient immune response and pathogen clearance (Takeda and others 1999; Williams and others 2004; Chan and others 2010; Sharada and others 2012). Our study validated that in HIV-infected individuals, the presence of high IL-10 levels resulted in reduced activity of macrophages with low MHC expression and minimal activation of monocytes corresponding to low levels of IFN-γ indicated their ineptitude to load the mycobacterial peptides on to the MHC.
Our previous study reported that simultaneous activation and apoptosis of CD8+ cells might be the reason for lack of CTL responses in HIV-positive individuals with TB (Lang and others 2002). Evidences from previous studies state that CD8+ T cells play a role in the control of latent TB (van Pinxteren 2000; Lewinsohn and others 2003, 2007; Chen and others 2009). Another important contributor to the increased risk of reactivation of latent TB and susceptibility to new M. TB infection in HIV-positive individuals is depletion of CD4+ T cells along with other mechanisms such as manipulation of macrophage bactericidal pathways and a tipped Th1/Th2 balance (Spear and others 1990; Havlir and Barnes 1999). Trautmann and others reported that upregulation of PD1 on HIV-specific CD8+ T cells was proportional to viral load and low production of cytokine along with proliferation of HIV-specific CD8 (+) T cells (Trautmann and others 2006). Our findings were similar to that of Trautmann and others, wherein increased expression of PD1 on activated CD8+ cells may cause CTL dysfunction along with decreased IFN-γ levels. There are reports that expression of PD1 is marked to be a highly proliferative pathogen-specific T cell precursor that replenishes protective cytokine-producing effector CD4+ cells during M. TB infection (Lazar-Molnar and others 2010; Reiley and others 2010; Pollock and others 2016; Day and others 2018). Surprisingly, our observations revealed that activation of CD8 cells was hindered on blocking PD1; this needs elucidation. Also, the increased expression of TAP2 on inhibition of PD1 validated the role of PD1 in defective functioning of TAP.
Finally, our observations provide an insight into the escape mechanisms by M.tb in HIV-infected individuals from the host immune responses leading to TB co-infection. The consequence of high IL-10 and PD1 expression is due to defective antigen presentation on macrophage due to perturbations in TAP mRNA expression.. Also, high expression of PD1 on CD8+ and low IFN-γ levels cause cytotoxic T cell dysfunction, Pathways involved in these escape mechanisms need to be elucidated further.
Authors' Contributions
S.R.S. and N.C.S. contributed to analysis and article writing. S.R.S., N.C.S. S.S.P., and V.S.K.N. contributed in experimentation.
Footnotes
Author Disclosure Statement
No competing financial interests exist.
Funding Information
Indian Council of Medical Research (Project. No: Indous/69/9/2010-ECD-II).
Supplementary Material
Supplementary Figure S1
Supplementary Figure S2