
Abstract
Lycopene (lyc) has an effect on preventing cancer, yet its effects on hypoxia/reoxygenation (H/R) injury remained obscure. The study aimed at discovering its role in preventing hepatic cells against H/R injury. Hepatic cells were incubated in hypoxia incubator to simulate ischemia/reperfusion injury in vitro. Cell viability was detected using 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) assay after Lycopene treatment with or without ML385 (nuclear factor erythroid 2-related factor 2 [Nrf2] inhibitor). Lactate dehydrogenase (LDH) and malondialdehyde (MDA) content were detected. Cellular cytokine (tumor necrosis factor-α, TNF-α; interleukin-6, IL-6) levels were measured using enzyme-linked immunosorbent assay (ELISA). Hepatic cell apoptosis and cellular reactive oxygen species (ROS) content was detected by flow cytometry. Nrf2 transfer was observed using immunofluorescence staining. Nrf2 and heme oxygenase-1 (HO-1) expressions were detected with quantitative real-time polymerase chain reaction and western blot as needed. In hepatic cells, after H/R, the viability was dropped, TNF-α and IL-6 levels and LDH and MDA content were increased, with high apoptosis rate and ROS content. Lycopene led to a reversed effect, with promotion on Nrf2 transfer from cytoplasm into nucleus and Nrf2/HO-1 pathway activation. Further experiments showed that ML385 could reverse the effects of Lycopene. Lycopene could activate Nrf2/HO-1 pathway to protect hepatic cells against H/R injury.
Introduction
Ischemic heart disease, a critical condition, which is caused by blood supply interruption, is recognized as the leading cause of death around the world (Zhu and others 2019). Restoration on coronary blood flow is seen as an effective intervention to alleviate myocardial ischemic injury, but reperfusion itself may induce more sustained and irreversible myocardial damages and remodeling on pathological remodeling. This is defined as myocardial ischemia/reperfusion (I/R) injury (Dang and others 2019).
Various factors may be implicated in myocardial I/R injury pathogenesis, which include oxidative stress, overload on intracellular calcium, inflammation, as well as apoptosis (Wu and others 2020), yet the detailed mechanism of myocardial I/R injury is complicated. Consequently, better and full understanding on the molecular pathogenesis underlying the dysfunction is urgently needed for finding a potential therapeutic method for myocardial protection.
Lycopene is an acyclic hydrocarbon, non-provitamin A carotenoid and is a potent antioxidant with properties on anticancer (Burke and others 2019). As Kim and Kim (2015) found, Lycopene has the potential anticancer effects on gastric carcinogenesis, which is mediated by oxidative stress. Also, in the study of Ye and others (2016), Lycopene could inhibit the proliferation and invasion of human head and neck squamous cell carcinoma through the inhibition on protein kinase B and mitogen-activated protein kinase pathway.
As for hypoxia/reoxygenation (H/R) injury, as illustrated in the study of Xu and others (2015), Lycopene could protect neonatal cardiomyocytes in mice by alleviating apoptosis induced by endoplasmic reticulum. However, its implication with nuclear factor erythroid 2-related factor 2 (Nrf2) and its binding gene heme oxygenase-1 (HO-1) in hepatic cancer is hardly discovered and discussed.
Nrf2 is a member belonging to the basic leucine transcription factor family, and has been recognized as a key regulator of cellular antioxidant response (Schmidlin and others 2019). It starts the transcription of corresponding downstream antioxidant molecules, which include Phase II detoxifying enzymes through binding to antioxidant reactive element (ARE) of the promoter region of genes, including HO-1 (Liu and others 2017). In addition, activation of Nrf2 has been found to enhance the levels of antitoxic enzymes as well as phase-2-detoxifying enzymes, which may be chosen as one of the ways on reduction of oxidative stress and chronic inflammation (Kurutas 2016).
More importantly, Nrf2 is a potential and possible target for cardioprotection and it can help cardiomyocytes endure oxidative damage during myocardial I/R injury and maintains redox homeostasis (Wang and others 2015; Shen and others 2019). However, its regulation with HO-1 in I/R-induced hepatic cells, which have been treated with Lycopene is rarely discussed. Since H/R cellular model has been used to simulate I/R injury in vitro (Chen and others 2017), our study, based on a H/R cellular model, aimed to discovering the possible effects and mechanisms of Lycopene on protecting hepatic cells from I/R-induced injury.
Materials and Methods
Cell culture and H/R model construction
Mouse hepatic cell AML12 used for the experiment was obtained from American Type Culture Collection (ATCC, catalog number: CRL-2254, Manassas, VA) and was subsequently incubated in Dulbecco's modified Eagle's medium (DMEM, 11965092; Thermo Fisher Scientific, Waltham, MA), which was supplemented with 10% fetal bovine serum (FBS; Invitrogen, Carlsbad, CA), 100 U/mL penicillin (Invitrogen), and 0.1 mg/mL streptomycin (Invitrogen).
For H/R model construction to simulate I/R injury in the cells, we pretreated cells with different concentrations of Lycopene (0.5, 1, 2, 5, 10, and 20 μM) for 4 h. Then, cells were collected and incubated in Galaxy® 48R hypoxia incubator (Eppendorf AG, Hamburg, Germany) with 5% CO2, 94% N2, and 1% O2 for 12 h, and subsequently cultured in oxygen chamber with 5% CO2 and 95% atmosphere for 4 h. Temperature for cell culture was 37°C for whole experiment.
3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide assay
Hepatic cells (1 × 104 cells/well) were seeded in 48-well plates, then they received H/R treatment (hypoxia for 12 h, reoxygenation for 4 h) and treated with different concentrations of Lycopene (0.5, 1, 2, 5, 10, and 20 μM; treated for 4 h). For cell viability detection, 10 μL 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) reagent (5 mg/mL; catalog number: K017; Fine Biotech Co., Ltd., Wuhan, China) was added into the cells. After being incubated at 37°C for 4 h, the supernatant of cell culture was dumped, and the visible formazan crystals was dissolved by 100 μL dimethyl sulfoxide (DMSO; 472301, Sigma-Aldrich Corp., St. Louis, MO). Then the values at an absorbance of 560 nm was measured and recorded using a SpectraMax Plus 384 microplate reader (Molecular Devices, LLC., San Jose, CA).
Lactate dehydrogenase leakage assay
After the cells were treated with H/R or Lycopene, 120 μL culture supernatant was seeded in 96-well plates and 60 μL of the Lactate Dehydrogenase (LDH) Detection Kit (catalog number: MK401; TaKaRa Bio, Tokyo, Japan) was added for further incubation at 37°C in the dark. About 30 min later, mass leakage of LDH was measured at an absorbance at 490 nm with the LDH Detection Kit following the manuals offered by the producer.
Detection of malondialdehyde content
Cells, which have undergone the treatments, were lysed using radioimmunoprecipitation assay (RIPA) buffer (P0013C; Beyotime, Shanghai, China) and then were centrifuged at 1600 g for 10 min. The supernatant obtained was subsequently mixed with 200 μL malondialdehyde (MDA) reaction solution (catalog number: M-012; Sigma-Aldrich Corp.) at 100°C for 15 min. Samples were then cold and centrifuged. The absorbance at 532 nm was captured to detect MDA content in the cells.
Enzyme-linked immunosorbent assay
After the hepatic cells were treated with H/R or Lycopene, enzyme-linked immunosorbent assay (ELISA) was used to measure the level of inflammatory factors in the supernatant of hepatic cells. After hepatic cells were separated following the centrifugation at 2,000 g for 15 min at 4°C, cell supernatant was available and then stored in aliquots at −80°C. Inflammatory factors, including tumor necrosis factor-α (TNF-α) and interleukin-6 (IL-6) levels in supernatant, were quantified with their corresponding ELISA Kits (catalog number: BMS607-3 for TNF-α; BMS603-2 for IL-6; Invitrogen) following the manufacturer's manuals. OD values were detected with a Crocodile 4-in-1 assay miniWorkstation (Titertek-Berthold Detection Systems GmbH, Pforzheim, Germany).
Flow cytometry
After the hepatic cells were treated with H/R or Lycopene, the apoptosis was detected by using the Annexin V-FITC/Propidium Iodide (PI) Apoptosis Kit (catalog number: K101; BioVision, Milpitas, CA), according to the producer's instructions. In short, cells were harvested and then washed with cold phosphate-buffered saline (PBS) twice. Then cells were treated with Annexin V and PI together for 15 min in the dark at room temperature. Cell apoptosis was then analyzed using flow cytometry assay (APO006; FLISP Serine Protease Detection Kits, Bio-Rad).
Reactive oxygen species assay
For reactive oxygen species (ROS) production detection, cells were suspended in solution of 2′, 7′-dichlorodihydrofluorescein diacetate (DCFH-DA; catalog number: D6883, Sigma-Aldrich Corp.). In brief, AML12 cells were washed with PBS after treatment and subsequently incubated with 10 μM DCFH-DA in a nonserum medium at 37°C for 30 min in the dark. Then cells were under observation with a fluorescence microscope (catalog number: BX51; Olympus Corporation, Tokyo, Japan), collected, and resuspended in the flushing fluid at a density of around 1 × 106 cells/mL. After being incubated with DCFH-DA, cells were analyzed using CytoFLEX flow cytometer (catalog number: B53008; Beckman Coulter Life Sciences, Indianapolis, IN) with a 525 nm emission filter.
Immunofluorescence staining and analysis
Immunofluorescence staining was used to detect the expressions of cellular Nrf2 in AML12 cells. In brief, AML12 cells were permeabilized with 1% Triton X-100 (Sigma-Aldrich, Inc.) in PBS for 20 min at room temperature. Subsequently, cells were blocked in 5% bovine serum albumin for 30 min, and were incubated with anti-Nrf2 antibody (rabbit, 1:3000, ab137550; Abcam, Cambridge, UK) at 4°C overnight. Then, the cells were incubated with secondary antibody (goat, ab205718, 1:2000; Abcam) for 1 h at room temperature after being washed 3 times with PBS.
Finally, the nuclei were stained using 4′, 6-diamidino-2-phenylindole (DAPI, D1306; Thermo Fisher Scientific) for 10 min and then the cells were washed 3 times with PBS in the dark. Immunofluorescence in the cells was later analyzed with a laser confocal microscope (LSM980; Carl Zeiss Microscopy, Oberkochen, Germany) in accordance with the manufacturer's manuals.
RNA isolation and quantitative real-time polymerase chain reaction
TRIzol reagent (Invitrogen, Madison, MI) was used to extract total RNA following the protocols of the producer. Then the extracted RNA was preserved in a 4°C or −80°C refrigerator. Total RNA concentration was later detected and quantified with a biological spectrometer (NanoDrop 2000; Thermo Fisher, Waltham, MA). The cDNA was synthesized from 1 μg of total RNA with a First-Strand cDNA Synthesis Kit (E6300L; New England Biolabs, Ipswich, MA) in accordance with the producer's instructions. Quantitative real-time polymerase chain reaction (PCR) experiment was conducted with the SYBR PremixEx Taq II Kit (RR820L; TaKaRa) in AriaMx real-time PCR System (G8830A; Agilent, Santa Clara, CA).
Primer sequences for this experiment were shown in Table 1. Glyceraldehyde 3-phosphate dehydrogenase (GAPDH) was used as an internal reference. Relative gene expressions were quantified with 2−ΔΔCT calculation method (Livak and Schmittgen 2001).
Primers for Quantitative Real-Time Polymerase Chain Reaction
GAPDH, glyceraldehyde 3-phosphate dehydrogenase; HO-1, heme oxygenase-1.
Western blot
Western blot was used to detect and measure Nrf2 (both nucleus and cytosolic) and HO-1 protein expressions in our study as previously described (Chen and others 2019). Shortly after collecting the cells, protein was first extracted from cytosolic and nucleus of cells with the Cytoplasmic and Nuclear Protein Extraction Kit (catalog number: AR0106; Boster Bio, Pleasanton, CA), and then lysed using RIPA buffer (P0013C; Beyotime), and protein concentration was then measured by the Bicinchoninic Acid (BCA) Protein Kit (P0012S; Beyotime). Thirty micrograms of lysates of the protein sample was subsequently electrophoresed by 12% sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE; P0012A; Beyotime), followed by transferring into polyvinylidene fluoride membrane (PVDF; FFP28; Beyotime), which was then blocked with 5% nonfat milk for 2 h.
The membrane was incubated in the primary antibodies, which consist of anti-Nrf2 antibody (rabbit, 1:3,000, ab137550; Abcam), anti-Lamin B1 antibody (rabbit, 1:1,500, ab220797; Abcam), anti-HO-1 antibody (rabbit, 1:2,000, ab13243; Abcam), and anti-GAPDH antibody (rabbit, 1:10,000, ab181602; Abcam) at 4°C overnight. Lamin B1 (for Nucleus Nrf2) and GAPDH (for Cytosolic Nrf2 and HO-1) were used as internal reference. The membrane was subsequently incubated in the secondary horseradish peroxidase (HRP)-combined antibody: goat anti-rabbit IgG H&L (HRP) (goat, 1:2,000, ab205718; Abcam) at room temperature for 1 h and washed with tris-buffer saline Tween (TBST) for 3 times.
Protein band was collected from the samples and was analyzed in an Enhanced Chemiluminescence (ECL) Kit (SW2020; Solarbio, Beijing, China). The gray values of the strips were further gathered and were under the calculation of ImageJ (version 5.0; Bio-Rad, Hercules, CA).
Statistical analysis
All experiments in our study have been conducted more than 3 times independently. Data collected from experiments were expressed as mean ± standard deviation (SD). Statistical analysis was performed with SPSS 20.0 software (IBM Corporation, Armonk, NY). Statistical significance was presented by one-way analysis of variance and Student's t-test followed by Dunnett's post hoc test. P value <0.05 was considered to address significant differences.
Results
Lycopene could improve the viability, ameliorate the injury, and relieve the oxidative stress in H/R-treated AML12 hepatic cells
To discover the effects of Lycopene on H/R-induced AML12 hepatic cells, we treated AML12 cells with different concentrations of Lycopene (0.5, 1, 2, 5, 10, and 20 μM) before the cells were treated with H/R. We then found that cell viability dropped significantly after H/R treatment (Fig. 1A, P < 0.001, versus Control). However, an improved cell viability was found in a dose-dependent manner after cells were treated with different concentrations (0.5, 1, 2, 5, 10, and 20 μM) of Lycopene, with the best effect found on cell viability after we treated cells with 10 μM of Lycopene (Fig. 1A, P < 0.001, versus H/R). Therefore, we chose 10 μM of Lycopene for subsequent study.
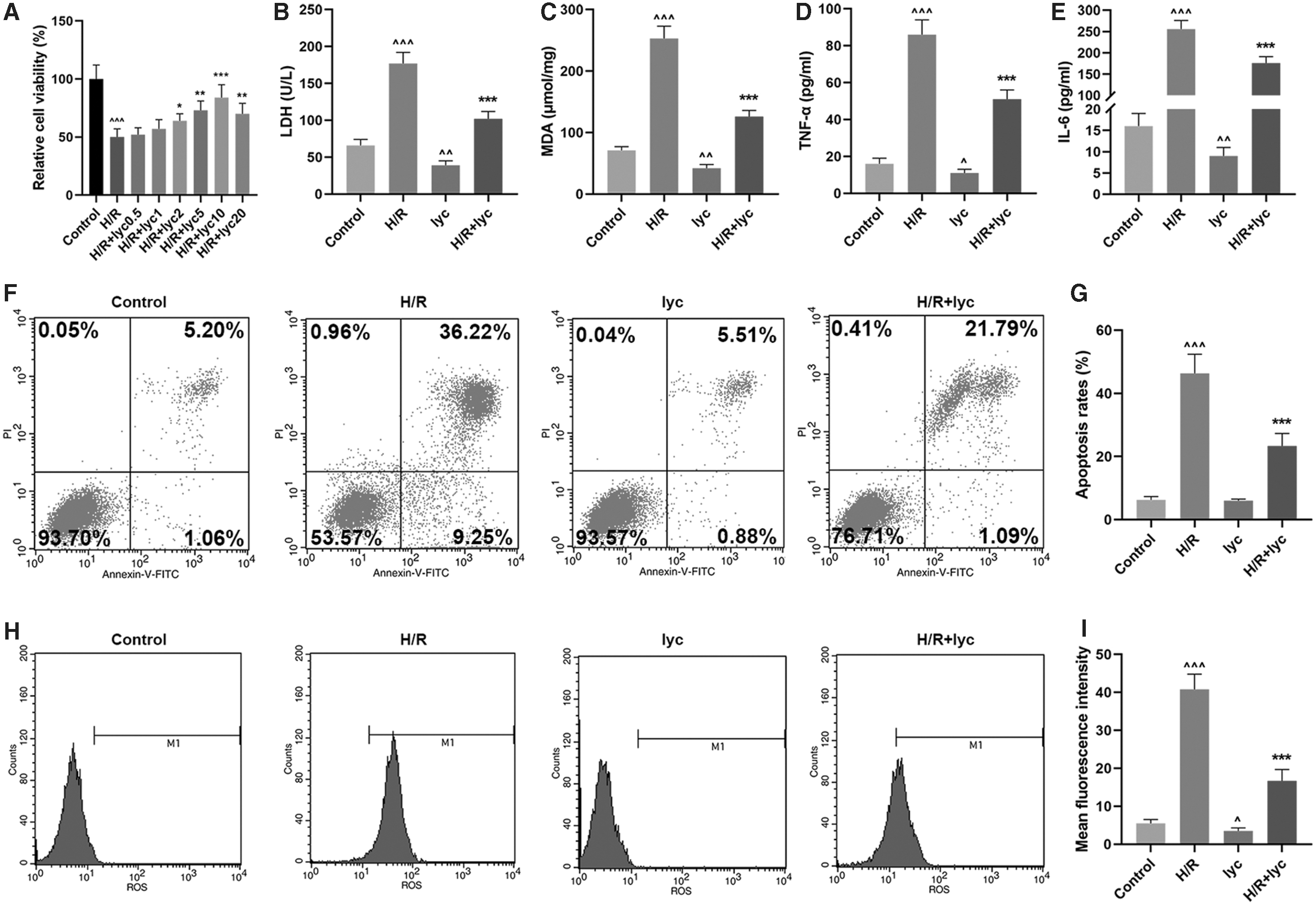
Effects of Lycopene on H/R-induced AML12 hepatic cells.
The leakage of LDH has been seen as a cellular marker for cell injury for disruption on plasma membrane (Li and others 2017). As Fig. 1B presented, LDH content in AML12 cells was significantly upregulated after H/R injury (P < 0.001, versus Control), whereas reduced content of LDH was found in cells, which have only received Lycopene treatment (P < 0.01, versus Control). Furthermore, an improved content of LDH was found in cells after treating H/R-induced cells with Lycopene (Fig. 1B, P < 0.001, versus H/R), suggesting that Lycopene could ameliorate injuries in AML12 hepatic cells.
In addition, to discover the potential effects of Lycopene on oxidative stress in the cells, we measured MDA content before or after Lycopene treatment in H/R-induced cells. It was found that MDA content in AML12 cells was evidently upregulated (Fig. 1C, P < 0.001, versus Control), whereas in cells treated with Lycopene alone led to an opposite result (Fig. 1C, P < 0.01, versus Control). In addition, after being treated with both H/R and Lycopene, MDA content in the cells was downregulated compared with H/R group (Fig. 1C, P < 0.001), showing that Lycopene could relieve the oxidative stress in AML12 hepatic cells.
Lycopene could reduce the levels of H/R-related cytokines and protect cells from H/R-induced apoptosis
To discover the potential effects of Lycopene on production and unleash of H/R-related cytokines, we measured the level of TNF-α and IL-6 in cell supernatants with ELISA. It was found that in cells with only H/R treatment, both TNF-α and IL-6 levels greatly upregulated (Fig. 1D, E, P < 0.001, versus Control), whereas cells with Lycopene treatment alone resulted in a different result (Fig. 1D, E, P < 0.01, versus Control). Besides, both levels of TNF-α and IL-6 were reduced after H/R and Lycopene treatment in comparison with H/R group (Fig. 1D, E, P < 0.001), showing that Lycopene could reduce the levels of H/R-related cytokines in AML12 hepatic cells.
The inhibitory effects of Lycopene on AML12 hepatic cell apoptosis were detected with flow cytometry. As illustrated in Fig. 1F and G, H/R treatment led to a high apoptosis rate in the cells (P < 0.001, versus Control), and after Lycopene treatment, the apoptosis rate in the cells was downregulated compared with H/R group (Fig. 1F, G, P < 0.001), suggesting that Lycopene could protect AML12 hepatic cells from apoptosis.
Lycopene could reduce H/R-induced ROS generation
Both the DCFH-DA fluorescence assay and flow cytometry assay were performed to examine cellular ROS level. In the experiments, we found that H/R treatment raised ROS level in the cells significantly, whereas ROS level in cells treated with only Lycopene was reduced (Fig. 1H, I, P < 0.001, versus Control). Also, we discovered that Lycopene treatment could reduce the generation of H/R-induced ROS in AML12 cells compared with H/R group (Fig. 1H, I, P < 0.001), which uncovered the role of Lycopene in reducing H/R-induced ROS generation of AML12 cells.
Lycopene could activate Nrf2/HO-1 pathway in AML12 hepatic cells
Nrf2 as well as its target gene HO-1 were found as significant regulators, which were responsive to several stresses (Loboda and others 2016). In our previous study, we showed that Lycopene could reduce H/Rq-induced ROS generation, but its detailed molecular mechanisms remained unclear. We subsequently investigated whether Lycopene could reduce H/R-induced ROS generation through modulating the Nrf2/HO-1 pathway. In the immunofluorescence assay, it was found that Lycopene treatment led to the translocation of Nrf2 into the nuclei of AML12 cells (Fig. 2A). Lycopene treatment upregulated the protein expression of Nucleus Nrf2 (Fig. 2B, C, ^^^P < 0.001, versus Control; ***P < 0.001, versus H/R). H/R treatment downregulated the protein expression of Cytosolic Nrf2 and promoted the HO-1 mRNA and protein expressions in hepatic cells (Fig. 2D–F, ^^^P < 0.001, versus Control; ***P < 0.001, versus H/R), and Lycopene treatment further enhanced the effects.

Lycopene could activate Nrf2/HO-1 pathway in AML12 hepatic cells.
In addition, H/R treatment inhibited the total Nrf2 mRNA and protein expression, while Lycopene treatment promoted the total Nrf2 mRNA and protein expression (Fig. 2G–I). These results indicated that Lycopene could activate Nrf2/HO-1 pathway in AML12 cells.
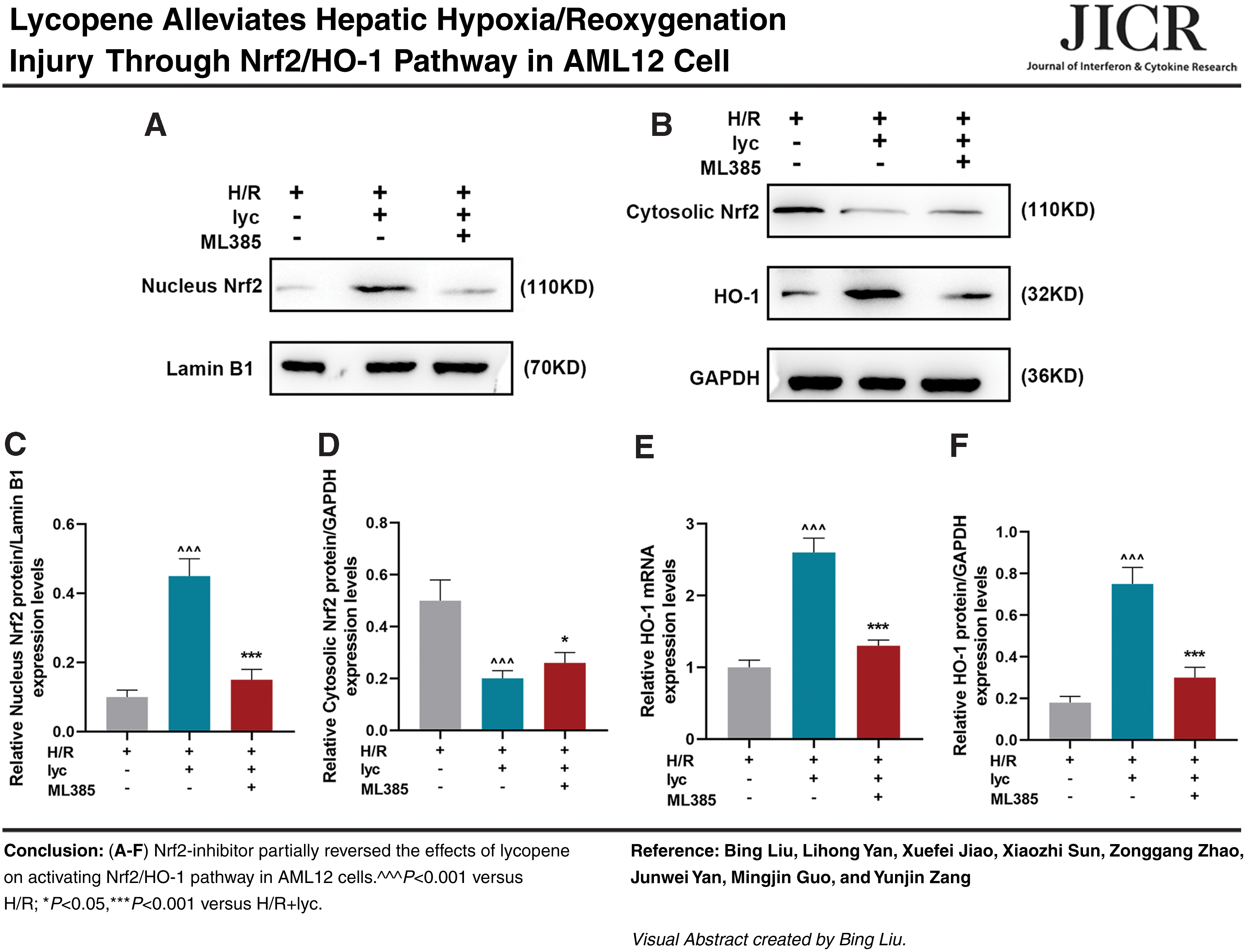
Nrf2-inhibitor partially reversed the protective effects of Lycopene in AML12 cells
To evaluate whether Nrf2 plays a vital role in Lycopene defense against oxidative damage, AML12 cells were pretreated with 1 μM ML385 (Nrf2 inhibitor) for 2 h before Lycopene treatment. In MTT assay, as Fig. 3A shows, Lycopene treatment could upregulate the H/R-induced cell viability compared with H/R group (Fig. 3A, P < 0.001). However, cell viability was dropped after cells were treated with ML385 in comparison with H/R+lyc group (Fig. 3A, P < 0.001), suggesting that Nrf2-inhibitor could partially reverse the promoting effects of Lycopene on the viability of AML12 cells.
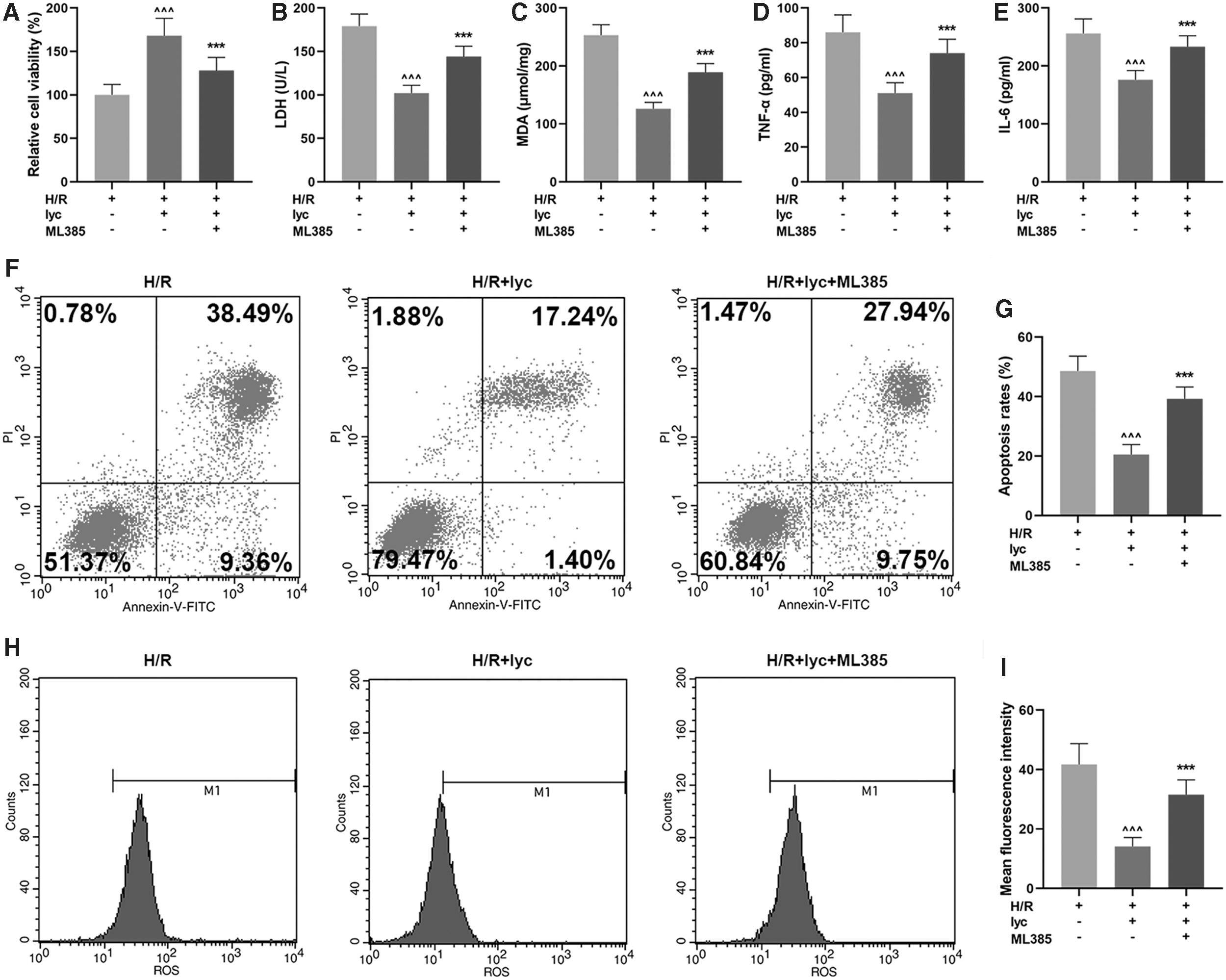
ML385 (Nrf2-inhibitor) partially reversed the protective effects of Lycopene in AML12 hepatic cells.
As for LDH and MDA content in AML12 cells, it is shown in Fig. 3B and C that both contents of LDH and MDA in H/R-induced AML12 cells with Lycopene treatment were downregulated compared with H/R group (Fig. 3B, C, P < 0.001), but after ML385 treatment, their contents in H/R-induced AML12 cells were increased compared with H/R+lyc group (Fig. 3B, C, P < 0.001), suggesting that Nrf2-inhibitor could partially reverse the effects of Lycopene on ameliorating the injury and relieving the oxidative stress in H/R-treated AML12 hepatic cells.
In Fig. 3D and E, it was found in ELISA that Lycopene could reduce H/R-related cytokines TNF-α and IL-6 levels compared with H/R (Fig. 3D, E, P < 0.001), yet a partially reversed result was found after ML385 treatment in comparison with H/R+lyc (P < 0.001), indicating that Nrf2-inhibitor was able to partially reverse the effects of Lycopene on reducing the levels of H/R-related cytokines.
In flow cytometry assay, we discovered a reduced apoptosis rate of H/R-induced AML12 cells after Lycopene treatment compared with H/R group (Fig. 3F, G, P < 0.001). However, a partially reversed effect on cell apoptosis was found after we treated cells with ML385 compared with H/R+lyc group (Fig. 3F, G, P < 0.001), which therefore suggested that Nrf2-inhibitor had the capability to reverse the protective effects on Lycopene against cell apoptosis.
Finally, we detected the content of ROS in H/R-induced AML12 hepatic cells with both the DCFH-DA fluorescence assay and flow cytometry assay. It was found in the DCFH-DA fluorescence assay and flow cytometry assay that the content of ROS was reduced after Lycopene treatment in comparison with H/R group (Fig. 3H, I, P < 0.001), whereas ML385 treatment led to a partially reversed effect compared with H/R+lyc (Fig. 3H, I, P < 0.001), showing that Nrf2-inhibitor reversed the effects of Lycopene on reducing ROS production in AML12 hepatic cells.
Nrf2-inhibitor partially reversed the effects of Lycopene on activating Nrf2/HO-1 pathway in AML12 cells
To discover the effects of Nrf2-inhibitor on Nrf2/HO-1 pathway, we measured Nrf2 (both Nucleus and Cytosolic) and HO-1 expressions in H/R-induced AML12 hepatic cells. It was discovered that both Nucleus Nrf2 and HO-1 expressions in H/R-induced AML12 hepatic cells after Lycopene treatment were upregulated, yet those of Cytosolic Nrf2 were downregulated compared with H/R (Fig. 4A–F, P < 0.001), indicating that Lycopene could promote the activation of Nrf2/HO-1 pathway. However, after we treated cells with ML385, a partially reversed result was discovered compared with H/R+lyc (Fig. 4A–F, P < 0.001), which therefore suggests that Nrf2-inhibitor could partially reverse the effects of Lycopene on activating Nrf2/HO-1 pathway in AML12 cells.

ML385 partially reversed the effects of Lycopene on activating Nrf2/HO-1 pathway in H/R-induced AML12 hepatic cells.
Discussion
Hepatic I/R injury has been found to occur in several clinical conditions, like trauma, hypovolemic shock, hepatic resection, as well as transplantation of liver (Wang and Yan 2017). Previous study suggested that the activation of Akt may be used as a potential target for hepatic I/R injury (Covington and others 2017), but detailed mechanisms in preventing the development and progression of I/R injury are rarely discovered and discussed. In our study, to the best of our knowledge, we first reported that Lycopene may have the potential effects on hepatic I/R injury treatment, with the regulation on Nrf2/HO-1 pathway.
LDH and MDA have been found as 2 biomarkers of oxidative stress (Sifi and Soltani 2019). LDH, a marker for cellular injury, could be released from cells when cells were injured, which was caused by several factors (Duan and others 2019). As Wang and others (2017) pointed out, the LDH activity indirectly indicated the degree of cell injury. MDA, however, has been recognized as one of the most popular and reliable markers for oxidative stress determination in clinical situations (Cwynar and others 2018). In our study, we discovered an increase of both LDH and MDA content in hepatic cells, showing that I/R injury could result in the onset of oxidative stress.
In recent years, cytokines have come up as biomarkers for local and systematic inflammation in several cardiovascular diseases (Dubnika and others 2018). Some proinflammatory cytokines, such as TNF-α and IL-6, have been depicted as independent coronary heart diseases risk factors and atherogenesis promoters (Kosmas and others 2019). In our study, likewise, we found both TNF-α and IL-6 levels in the cells were upregulated after H/R cell model was established, which therefore suggests that downregulating these cytokines may be of great help to alleviate cardiovascular injuries.
ROS are those short-lived and highly reactive molecules and excess ROS levels under H/R in the cells have been found to be associated with activating several processes related to cell death, such as apoptosis (Redza-Dutordoir and Averill-Bates 2016). In our study, after the establishment of cellular I/R model, we discovered a significant increase on ROS content in hepatic cells, with an evidently upregulated cell apoptosis rate. Consequently, ROS elimination using antioxidants may be selected as a critical way on restraining cell injury induced by ROS (Li and others 2017).
Lycopene a non-pro-vitamin A carotenoid and mainly synthesized by microorganisms and plants, especially by tomatoes, has become well known for its antioxidant properties (Francieli da Silva and others 2015). It was discovered and discussed in the research of Liu and others (2018) that Lycopene could ameliorate oxidative stress in ovary of aging chicken through activating Nrf2/HO-1 pathway. As for treatment on I/R-injury, in the study of Yue and others (2015), Lycopene has been found to have a cardioprotective effect in treating myocardial I/R injury . In our study, increased hepatic cell viability, yet reduced cell apoptosis, was found after Lycopene treatment, with the effects on cell injury amelioration, which shows the protective role of Lycopene on hepatic cells from H/R-induced injury.
Nrf2 has been suggested to regulate the cellular cycle of hepatocyte (Muller and others 2018) and accumulating evidences have unveiled that Nrf2 activation plays a vital role in kidney protection against several disorders (Lv and others 2018). Nrf2 is generally known to enhance the cellular defense system to counteract oxidative injury and inflammation (Hassanein and others 2020). Under normal conditions, Nrf2 exists in the cytoplasm sequestered by Keap1 (Itoh and others 1999). When exposed to reactive species, like ROS, Nrf2 is found to be translocated from cytoplasm into the nucleus and subsequently binding to the cis-acting ARE with gene promoters, including HO-1, which could attenuate the kidney function and morphology during I/R injury (Ge and others 2017; Barakat and others 2018).
It has been reported that Nrf2 activation promotes the expression of HO-1, which protects the cells against apoptosis (Yonar 2013). HO-1 and their products exert beneficial effects through the protection against oxidative injury, regulation of apoptosis, and modulation of inflammation (Loboda and others 2016). Lycopene attenuates colistin-induced nephrotoxicity in mice through activation of the Nrf2/HO-1 pathway (Dai and others 2015). Lycopene ameliorates atrazine-induced oxidative damage in the adrenal cortex of male rats by activation of the Nrf2/HO-1 pathway (Abass and others 2016).
In conditions with oxidative stimuli, the dissociation of Keap1 from the CUL-E3 ligase, which modifies the cysteine residues of Keap1, in particular Cys151, leads to accumulation of Nrf2 (Eggler and others 2005), as a result, Nrf2 liberates and translocates into the nucleus where it binds to the ARE and promotes the transcription of antioxidant genes (Venugopal and Jaiswal 1998).
Lycopene has been shown to be a great scavenger of free radicals and a potential antioxidant attributing to the 11 conjugated bonds within the molecule (Kelkel and others 2011; Petyaev 2016), Lycopene is widely used for protection against oxidative stress-mediated tissue injury (Liu and others 2018). Lycopene is capable of scavenging nitrogen dioxide, thionyl, and thionyl radicals (Mortensen and Skibsted 1997).
In our study, we found Lycopene treatment could promote the translocation of Nrf2 into cell nucleus and the activation of Nrf2/HO-1 pathway and therefore ameliorating H/R injury in hepatic cells, which was consistent with a previous study (Lei and others 2016), where it indicated that Lycopene may promote the dissociation of Keap1 and lead to the nuclear translocation of Nrf2. In addition, an opposite result was found after inhibiting Nrf2 expression in hepatic cells, which further showed the effects of Lycopene on activating Nrf2/HO-1 pathway and ameliorating cellular H/R injury. However, the effects of HO-1 inhibitor on the cell viability, apoptosis, and production of LDH, MDA, ROS, TNF-α, and IL-6 needs further analysis.
However, there are some limitations in our study. In our present study, we only discovered the role of Lycopene on ameliorating H/R injury in hepatic cells, but its efficacy on animals was unclear. Therefore, further researches are required. In addition, the effect of Lycopene on the proliferation of H/R-induced hepatic cell needs further study. The specific regulatory mechanism of Lycopene inducing Nrf2 nuclear translocation remains to be further studied.
Taken together, the results from our study provided a new evidence for Lycopene on ameliorating hepatic H/R injury. In our study, Lycopene treatment led to the increase on hepatic cell viability and activating Nrf2/HO-1 pathway, yet a decrease on the H/R-induced cytokine production and cellular LDH and MDA content, with low apoptosis rate and ROS content. These results showed the potential effects of Lycopene on hepatic H/R injury amelioration, with the potential therapeutic method for hepatic H/R injury amelioration.
Footnotes
Author Disclosure Statement
No competing financial interests exist.
Funding Information
No funding was received for this article.