
Abstract
Graft-versus-host disease (GVHD) is a physiological response of the graft to allogeneic hosts. However, the effector cells, affected organ(s), and cytokines in the GVHD remain controversially discussed, without having determined a particular cytotoxic activity of the graft against the host. After i.v. injection of C57BL/6 (H-2b) spleen cells into irradiated BDF1 (H-2b/d) mice, the hosts developed interferon-gamma (IFN-γ)-dependent bone marrow (BM) GVHD on days 5–17. When H-2DdKd transgenic H-2b lymphoma cells were i.p. inoculated into irradiated, H-2b splenocyte-transplanted H-2b/d mice, the infiltration of macrophages cytotoxic against H-2DdKd transgenic H-2b mouse skin epithelia (a GVHD activity) into the peritoneal cavity preceded several days the infiltration of interleukin (IL)-2-dependent cytotoxic T lymphocytes (CTLs) to achieve a graft-versus-leukemia (GVL) effect. In contrast, allogeneic BM transplanted alone into the irradiated mice did not induce GVHD for 44 days, whereas i.v. injection of graft anti-host macrophages or graft anti-host CTLs along with allogeneic BM, respectively, induced GVHD or promoted the GVL effect in the absence of GVHD. These results revealed that macrophage-induced GVHD and the CTL-mediated GVL effect were a set (Th1: IFN-γ/IL-2) response of the graft to allogeneic hosts and leukemia cells, respectively, and that graft T cell activation rather than inhibition skipped GVHD after BM transplantation.
Introduction
In the 1970s, the critical role of T cells in discriminating between “self” and “non-self” was established by the failure of neonatally thymectomized and congenitally athymic nude rodents to reject skin or organ allografts (Pennycuik 1971). In the 1990s, on the one hand, studies using CD8 or CD4 knockout (KO) mice revealed that CD4+ helper T cells, but not CD8+ cytotoxic T lymphocytes (CTLs), are essential for the rejection of organ or skin allografts (Dalloul and others 1996; Krieger and others 1996; Van Buskirk and others 1996). On the other hand, we had earlier demonstrated that allograft-induced macrophages are cytotoxic against CTL-resistant, Meth A cells or skin epithelia (Yoshida and others 1988, 1991; Yamamoto and others 1998). In the 2010s, we reported that interferon-gamma (IFN-γ)-dependent monocyte/macrophage MHC receptor (MMR) expression on macrophage is essential for rejection of allografted skin in mice (Tashiro-Yamaji and others 2013; Yoshida 2014; Yoshida and others 2020).
Bone marrow (BM) transplantation has demonstrated a curative potential in patients with BM failures or hematological malignancies (Appelbaum and Thomas 1983; Eibl and others 1996). However, the cells transplanted into irradiated hosts become cytotoxic against allogeneic hosts [graft-versus-host disease (GVHD)] and leukemia cells [graft-versus-leukemia (GVL) effect], both of which are known to be mediated by CTLs (Ferrara and others 1996). Therefore, on the one hand, preventing GVHD, while preserving the GVL effect, has been a challenge in BM transplantation (Spoerl and others 2014; Messmann and others 2015; Ueha and others 2017). On the other hand, the most common GVHD prophylaxis has been based on immunosuppressive agents to attenuate T cell activation, but 20% to 80% of recipients still develop GVHD (Zeiser and Blazar 2017). Recently, there have been conflicting reports as to the effector cell type (eg, CTL or macrophage), the affected organ (eg, skin or BM), and the role of cytokines (eg, Th1 and Th2) in the pathobiology of GVHD in mice and humans (Henden and Hill 2015; Divito and others 2020; Jardine and others 2020).
In this study, we found that IFN-γ-activated macrophage-induced BM GVHD and the interleukin (IL)-2-dependent CTL-mediated GVL effect were a set (Th1: IFN-γ/IL-2) reaction and that the i.v. injection of graft anti-host CTLs simultaneously with allogeneic BM skipped macrophage-induced GVHD. This is a novel approach to prevent GVHD after BM transplantation by graft T cell activation rather than inhibition.
Materials and Methods
Antibodies
Fluorescein isothiocyanate (FITC)-labeled anti-mouse CD8 (53–6.7) and Ly-6G (1A8) antibodies (Abs); phycoerythrin (PE)-labeled anti-mouse CD4 (RM4–4), Mac-1 (M1/70), and NK-1.1 (PK136) Abs; and unlabeled anti-mouse TCRβ (H57–597), TCRγδ (GL3), Thy-1.2 (53–2.1), CD8 (53–6.7), CD4 (RM4–4), and CD3 (145–2C11) Abs from PharMingen (San Diego, CA).
Mice
Specific pathogen-free male (or female) A/J, C3H/He, C57BL/6, CBA/N, BALB/c, DBA/2, and BDF1 mice (7 weeks old) were purchased from Japan SLC (Hamamatsu, Japan). IFN-γ KO C57BL/6 mice were kindly donated by Dr. Y. Iwakura, Institute of Medical Science, University of Tokyo (Tagawa and others 1997). H-2DdKd transgenic or MMR1/2 KO C57BL/6 mice were generated in our laboratory (Inoue and others 2011; Tashiro-Yamaji and others 2013). All experiments were carried out in accordance with The Japanese Government Notification on Feeding and Safekeeping of Animals (Notification No.6 of the Prime Minister's Office); and the experimental protocol was approved by The Review Committee for Animal Experiments of Osaka Medical and Pharmaceutical University.
Tumor cells
EL-4 (C57BL/6 origin) T lymphoma cells were purchased from American Type Culture Collection (Rockville, MD). H-2DdKd transgenic EL-4 cells were generated in our laboratory (Inoue and others 2011).
Flow cytometric analysis
Cells (106 cells) were stained with FITC- or PE-labeled Abs for 20 min at 4°C, washed, analyzed, and sorted by fluorescence-activated cell sorter (FACS; FACStar®, Becton Dickinson, Mountain View, CA).
Cell preparations
(1) Peritoneal exudate cells (PECs): on days 0–20 after an i.p. transplantation of H-2DdKd transgenic EL-4 tumor cells (3 × 106 cells/mouse) into various strains of mice, PECs were obtained as described (Yoshida and others 1991). (2) T cell- or CD8+ or CD4+ T cell-depleted populations were obtained after treatment of PECs or spleen cells with anti-Thy-1.2 or anti-CD8 or anti-CD4 Ab and complement. (3) Macrophages (or granulocytes) were obtained as Ly-6G−/Mac-1+ (or Ly-6G+) cells by FACS. (4) Concanavalin A (Con A) blasts were prepared as described previously (Yoshida and others 1997b). (5) C57BL/6 anti-DBA/2 CTLs were induced by mixed lymphocyte culture (MLC), as described earlier (Yoshida and others 2020). (6) Mouse skin components were obtained, as described previously (Yamamoto and others 1998). Among them, hair matrix cells and epidermal keratinocytes appeared to be labeled with 51Cr (Taylor and others 2000; Hirota and others 2001).
BM transplantation
Male (or female) C57BL/6 (or BDF1) mice, 6–8 weeks old, were lethally (9 Gy) or sublethally (4 Gy) irradiated with a Cs source HW-150 (HITEX; Osaka, Japan) at a rate of 36.7 rad/min with a 0.25-mm Cu filter, as described previously (Ushio-Umeda and Yoshida 1997). Some of the lethally irradiated C57BL/6 mice were reconstituted on day 2 by i.v. injection of BALB/c BM cells, spleen, and/or exudate cells (each 2.5 × 106 cells). Some of the sublethally irradiated BDF1 mice were reconstituted on day 2 by i.v. infusion of C57BL/6 BM cells+MLC-induced C57BL/6 anti-DBA/2 CTLs (each 3 × 107 cells).
Spleen cell transplantation
Bulk (3 × 107 cells) or CD4+ or CD8+ T cell-depleted spleen cells from wild-type or IFN-γ (or MMR1/2) KO C57BL/6 mice were i.v. transplanted on day 2 into sublethally irradiated BDF1 (or H-2DdKd transgenic C57BL/6) mice.
GVHD clinical score
Mice were monitored daily for survival after transplantation and assessed clinically by a standard scoring system comprising individual scores for weight loss, posture, activity, fur texture, and skin integrity (Cooke and others 1998).
Hematology
Red blood cell (RBC) counts, platelet counts, and hemoglobin (Hb) concentration were evaluated by the sheath flow direct current detection method (for RBC and platelet) or by the sodium lauryl sulfate method (for Hb). Hematocrit (Hct; %), mean corpuscular volume (MCV; fL), mean corpuscular Hb (MCH; pg), and mean corpuscular Hb concentration (MCHC; g/dL) were calculated.
Cytotoxic activity assay
Effector cells were mixed with 51Cr-labeled targets in U-shaped microtiter wells, as described previously (Yamamoto and others 1998).
Cold target inhibition assays
To determine whether the same H-2 antigen could be responsible for the lysis of 51Cr-labeled target cells, we used C57BL/6, C3H/He, CBA/N, A/J, DBA/2 and BALB/c Con A blasts as cold (unlabeled) targets.
Test for endocytosis
An in vitro test for endocytic activity was performed on adherent cell populations, as described previously (Yoshida and others 1991).
Hematoxylin-eosin staining
The excised mouse tissues were fixed in 10% formalin solution and stained with hematoxylin-eosin, as described previously (Yoshida and others 2020).
Results
GVHD induction by i.v. transplantation of C57BL/6 spleen cells into sublethally irradiated BDF1 mice
When C57BL/6 (H-2b) spleen cells were i.v. infused into sublethally irradiated BDF1 (H-2b/d) mice, the BDF1 mice showed a time-dependent decrease in body weight starting around day 5 as the clinical, but not skin (eg, ruffled facial fur and alopecia), GVHD scores reciprocally increased; and the mice died within 2–3 weeks (Fig. 1a). In contrast, after i.v. injection of spleen cells from syngeneic BDF1 donors into irradiated BDF1 recipients, the BDF1 mice exhibited a time-dependent increase in the body weights, and the clinical GVHD scores were retained at the base line (Fig. 1a).
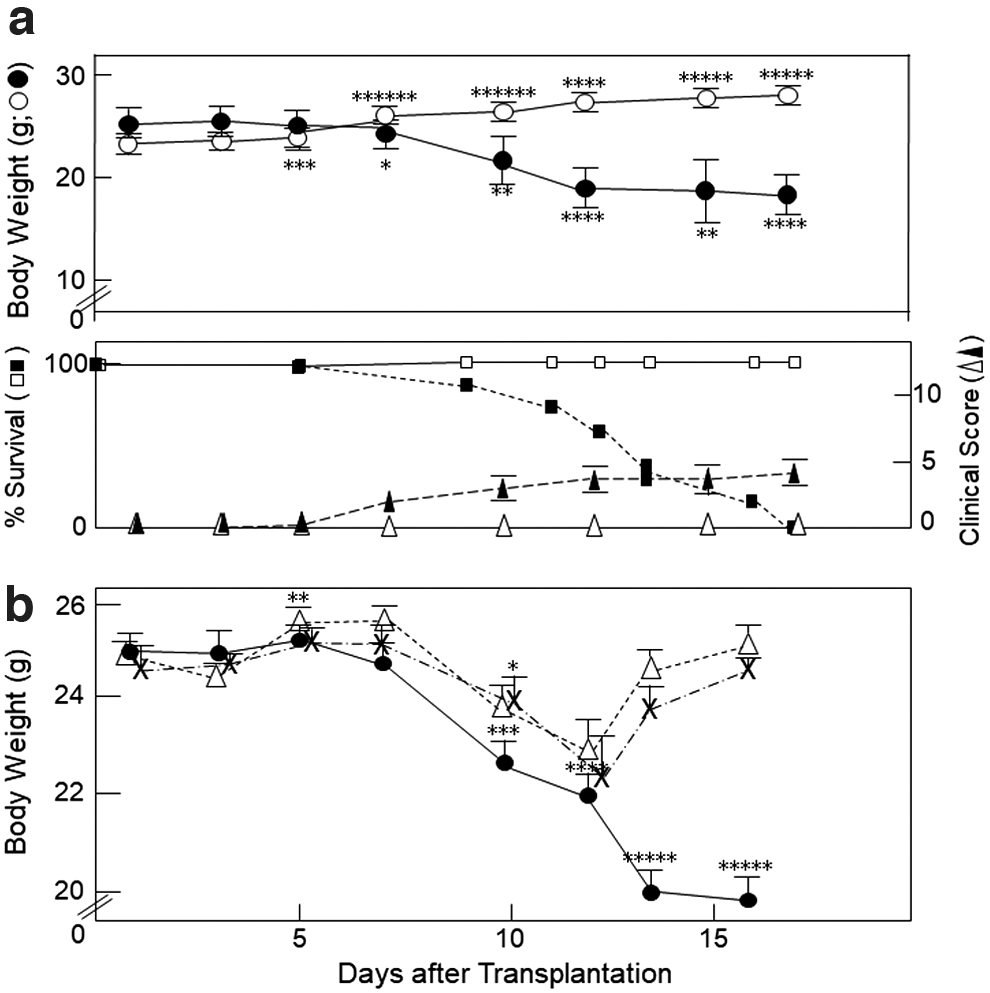
GVHD induction after i.v. transplantation of C57BL/6 spleen cells into sublethally irradiated BDF1 mice.
To determine which type of cells in the spleen population was involved in the induction of GVHD, we i.v. infused bulk (as a control) or CD4+ or CD8+ T cell-depleted spleen cells from C57BL/6 mice into irradiated BDF1 mice on day 2 (Fig. 1b). CD4+ or CD8+ T cell-depleted splenocyte-transplanted BDF1 mice commenced losing their body weight on day 7, which loss in turn began to increase on day 12, suggesting that both CD4+ and CD8+ T cells in the spleen might have been involved in the induction of GVHD on days 5–12.
Induction of GVH and GVL activities after i.p. injection of H-2DdKd transgenic EL-4 cells into irradiated, C57BL/6 splenocyte-transplanted BDF1 mice
When we i.p. inoculated H-2DdKd transgenic EL-4 (H-2b) T lymphoma cells into irradiated BDF1 (H-2b/d) mice, the tumor cells grew time dependently in the transplantation site and then died on days 18–20 (Fig. 2a, inset). At all intervals after the injection, there was no cytotoxic activity against 51Cr-labeled lymphoma cell targets in the cells infiltrating into the peritoneal cavity. In contrast, when H-2DdKd transgenic EL-4 (H-2b) T lymphoma cells were i.p. injected into irradiated, H-2b splenocyte-transplanted BDF1 mice, the growth rate was remarkably decreased (Fig. 2a, inset) with increasing cytotoxic activities of PEC against the lymphoma cells (Fig. 2a), suggesting a GVL effect of PECs on the host-type T lymphoma cells.
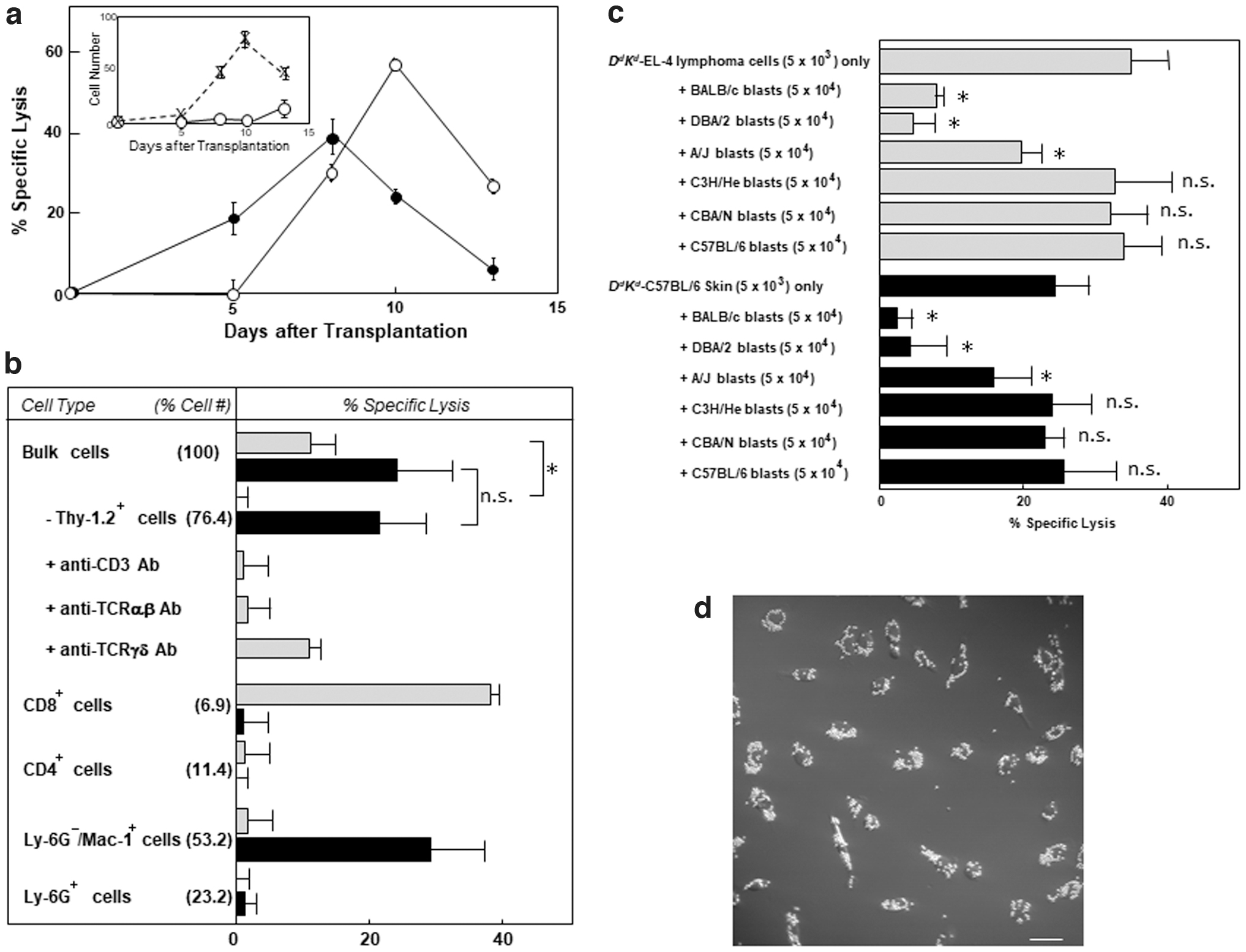
GVH and GVL activities of PEC after i.p. injection of H-2DdKd
transgenic EL-4 cells (3 × 106 cells/mouse) into irradiated, C57BL/6 splenocyte-transplanted BDF1 mice.
Most of the cytotoxic activity of PECs (day 10) against H-2DdKd transgenic EL-4 lymphoma cells disappeared after specific removal of T cells by complement-dependent cell lysis with anti-Thy-1.2 Ab; and the cytotoxic activity of PECs was largely suppressed by the addition of anti-CD3 or anti-TCRβ, but not by that of anti-TCRγδ, Ab (Fig. 2b). Figure 2b also shows that FACS-isolated CD8+, but not CD4+, Ly-6G+, or Ly-6G−/Mac-1+, cells were cytotoxic against the lymphoma cells. The cytotoxic activity against 51Cr-labeled H-2DdKd transgenic EL-4 cells was largely suppressed by the addition of unlabeled H-2d or H-2a, in that order, but not by that of unlabeled H-2k or H-2b, Con A blasts (Fig. 2c), indicating that the major effector cells responsible for the GVL effect were H-2d-specific, Thy-1.2+/CD3+/CD8+/TCRαβ + cells.
Unexpectedly, after i.p. injection of CTL-susceptible, H-2DdKd transgenic EL-4 lymphoma cells into irradiated, H-2b splenocyte-transplanted H-2b/d mice, the bulk infiltrates also exhibited time-dependent changes in cytotoxic activities against CTL-resistant, H-2DdKd transgenic C57BL/6 mouse skin epithelia (Fig. 2a). The infiltration of cells cytotoxic against skin epithelia (a GVH activity) into the peritoneal cavity preceded by several days the infiltration of CTLs cytotoxic against the lymphoma cells (a GVL effect).
Most of the cytotoxic activity of bulk PECs (day 8) against 51Cr-labeled H-2DdKd transgenic H-2b skin epithelia was retained after specific removal of T cells and was recovered by the Ly-6G−/Mac-1+ (53.2% of PECs), but not by the CD8+ (6.9%) or CD4+ (11.4%), cells; and Ly-6G+ granulocytes (23.2%) were inactive toward both targets (Fig. 2b). In contrast, virtually no NK-1.1+ cell had infiltrated into the peritoneal cavity. The cytotoxic activity against 51Cr-labeled H-2DdKd transgenic H-2b skin epithelia was largely suppressed by the addition of unlabeled H-2d or H-2a, in that order, but not by that of unlabeled H-2k or H-2b, Con A blasts (Fig. 2c). The Ly-6G−/Mac-1+ cells phagocytosed large numbers of the particles (Fig. 2d), suggesting that the major effector cells responsible for the GVH activity might have been H-2d-specific, Ly-6G−/Mac-1+ macrophages.
Induction of BM GVHD by i.v. inoculation of C57BL/6 spleen cells into irradiated BDF1 mice
Sublethally irradiated BDF1 mice showed normal histology of skin, lung, or intestine on day 12 (data not shown). The minimal histological criteria for skin, gastrointestinal tract, and lung GVHD require apoptosis within the basilar or lower spinosum layers of the epidermis; enterocyte apoptosis, crypt or basilar gland destruction, and mucosal denudation; and constrictive bronchiolitis obliterans, respectively (Schulman and others 2015). In the case of (C57BL/6→BDF1) GVHD mice, there were a few cells infiltrating into the skin or lung; but no tissue degeneration or apoptotic damage (eg, reduced cellularity, apoptotic bodies, and vacuolization) was seen in these tissues (Fig. 3). On the other hand, on day 9 after i.v. injection of bulk H-2b spleen cells into irradiated H-2b/d mice, the numbers of BM cells, spleen cells, H-2Dd+ peripheral white blood cells (WBCs), H-2Dd+ BM cells, and H-2Dd+ spleen cells decreased markedly (Table 1). Table 1 also shows that the numbers of total spleen cells and peripheral blood WBCs were almost normalized after i.v. infusion of CD4+ or CD8+ T cell-depleted spleen cells.
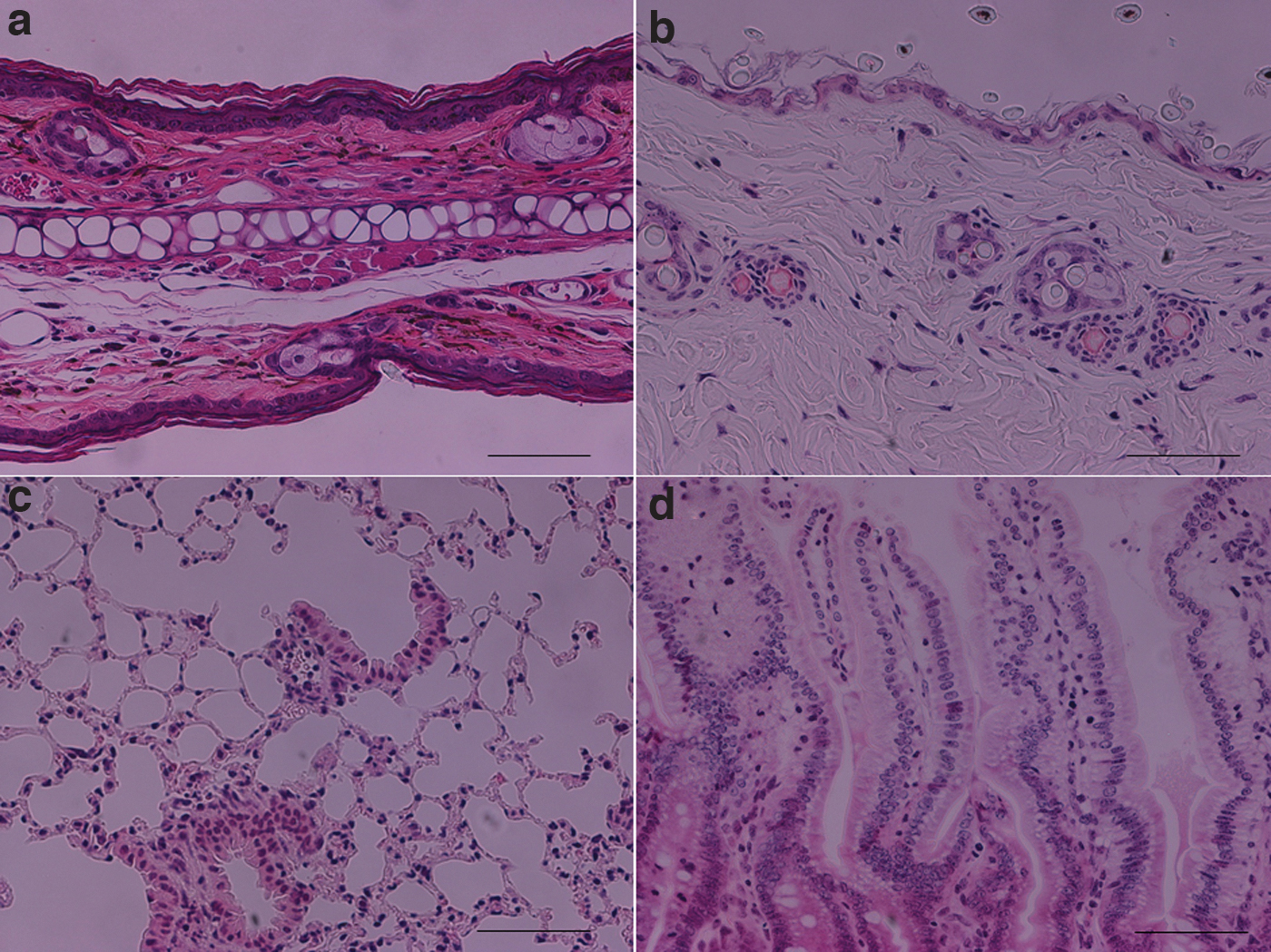
Representative HE staining of
Numbers of Leukocytes in Spleen, BM, and Peripheral Blood of Irradiated BDF1 Mice on Day 9 After i.v. Transplantation of C57BL/6 Mouse Spleen Cells
As a control, BDF1 mouse spleen cells were i.v. transplanted into irradiated BDF1 mice.
Each value represents the mean ± SD of 3 different experiments.
Each value represents the mean of 3 different experiments.
The difference between sample and the control (BDF1→BDF1) was significant (* P < 0.05; ** P < 0.01; *** P < 0.005) according to Student's t-test. n.s., not significant.
BM, bone marrow; SD, standard deviation; WBC, white blood cell.
Next, we assessed the numbers of RBCs, Hb, Hct, MCV, MCH, MCHC, platelets, and WBCs in the peripheral blood on day 9 after i.v. transplantation of bulk or CD4+ or CD8+ T cell-depleted H-2b spleen cells into irradiated H-2b/d mice. In the case of i.v. injection of bulk spleen cells, the numbers of RBCs, Hb, Hct, platelets, and WBCs remarkably decreased by 56%, 57%, 58%, 77%, and 37%, respectively, whereas the MCV, MCH, and MCHC values were almost unchanged (Table 2), suggesting that a type of normocytic, normochromic anemia with severe pancytopenia might have been induced by GVHD. Table 2 also shows that the BM GVHD was partly normalized by depletion of CD4+ or CD8+ T cells from the spleen cell population.
Numbers of RBCs, Hb, Hct, MCV, MCH, MCHC, Platelets, and WBCs in Peripheral Blood of Irradiated BDF1 Mice on Day 9 After i.v. Transplantation of C57BL/6 Mouse Spleen Cells
As a control, BDF1 mouse spleen cells were i.v. transplanted into irradiated BDF1 mice.
Each value represents the mean ± SD of 4 mice.
The difference between sample and the control (BDF1→BDF1) was significant (* P < 0.05; ** P < 0.02; *** P < 0.01; **** P < 0.005; ***** P < 0.001) according to Student's t-test. n.s., not significant.
Hb, hemoglobin; IFN-γ, interferon-gamma; KO, knockout; MCH, mean corpuscular Hb; MCV, mean corpuscular volume; MCHC, mean corpuscular Hb concentration; RBC, red blood cell.
Absence of BM GVHD after i.v. transplantation of IFN-γ KO C57BL/6 (H-2b) mouse spleen cells into irradiated BDF1 (H-2b/d) mice
IFN-γ-dependent MMR expression on macrophage is essential for H-2 haplotype-specific cytolysis of allogeneic skin epithelia (Tashiro-Yamaji and others 2013; Yoshida and others 2020). When IFN-γ KO C57BL/6 (H-2b) mouse spleen cells were i.v. injected into irradiated BDF1 (H-2b/d) mice, the numbers of RBCs, Hb, Hct, platelets, and WBCs on day 9 were at the normal levels (Table 2). Similarly, the BM GVHD was not seen after i.v. injection of MMR1/2 KO C57BL/6 mouse spleen cells into irradiated H-2DdKd transgenic C57BL/6 mice (data not shown). In contrast, on day 9 after i.p. injection of H-2DdKd transgenic EL-4 cells into irradiated, IFN-γ (or MMR1/2) KO C57BL/6 mouse splenocyte-transplanted BDF1 (or H-2DdKd transgenic C57BL/6) mice, CD8+ cells among the PECs exhibited the GVL effect [40.2 ± 0.4 (IFN-γ KO) or 29.9 ± 1.0 (MMR1/2 KO) % specific lysis at an E/T ratio of 5 in a 4-h assay]. Therefore, the terms GVHD macrophage and GVL lymphocyte were considered to be an appropriate designation for these types of macrophage and lymphocyte, respectively.
Effects of i.v. injection of GVHD macrophages or GVL lymphocytes along with allogeneic BM into irradiated hosts on GVHD
Lethal (9 Gy) irradiation alone caused a time-dependent weight loss, resulting in the death of all C57BL/6 mice within 10–11 days (Fig. 4a). When T cell-depleted, CD8+ T cell-depleted, CD4+ T cell-depleted, or bulk anti-host infiltrates or bulk spleen cells along with BM or BM alone were i.v. injected into lethally irradiated C57BL/6 mice, the 50% survival time was 13, 17, 20, 26, >40, or >40 days, respectively (Fig. 4a): Unexpectedly, after i.v. inoculation of BM alone or BM+nonsensitized spleen cells into irradiated C57BL/6 mice, the mice survived for 44 days; and their body weights were, respectively, retained within a line of −10% weight loss or remained at approximately the original level (day 1) until day 44 (Fig. 4a). However, [BM+donor anti-host macrophages ( = bulk infiltrates–T cells)] induced GVHD and [BM+bulk or CD4+ (or CD8+) donor anti-host lymphocytes] strongly or partially ameliorated the macrophage-induced GVHD (Fig. 4a).
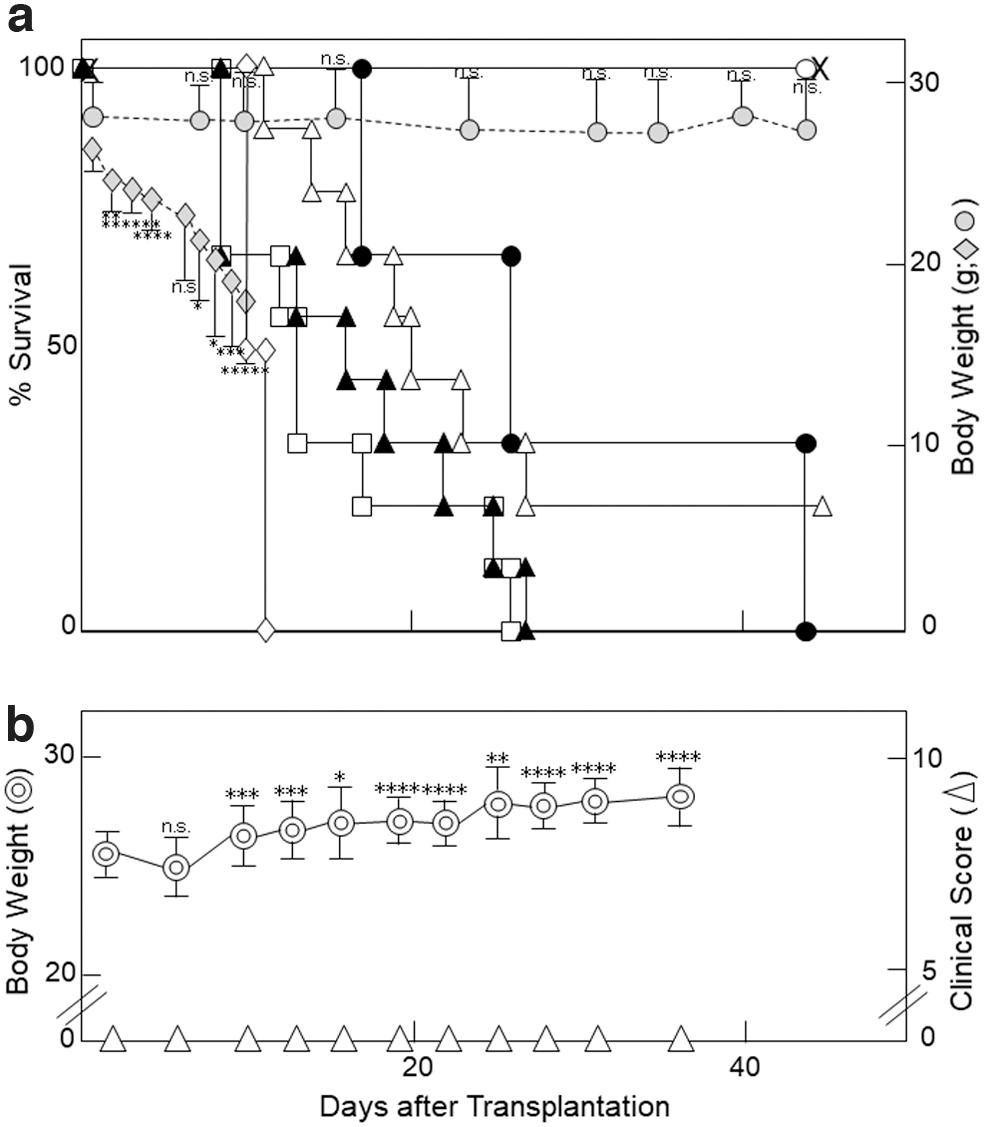
Control of Th1 [IFN-γ/IL-2 (or GVHD macrophage/GVL lymphocyte)] response.
Next, to skip the GVHD on the GVL effect after BM transplantation, we i.v. injected MLC-induced C57BL/6 anti-DBA/2 CTLs ( = bulk infiltrates–macrophages) along with C57BL/6 BM into sublethally irradiated BDF1 mice. Of particular interest, the BDF1 mice showed a time-dependent normal increase in body weight without any increase in GVHD scores on days 1–35 (Fig. 4b), as had been observed with i.v. injection of syngeneic BDF1 spleen cells into irradiated BDF1 mice (Fig. 1a).
Discussion
In animal models, donor T lymphocytes play an essential role in immune-mediated damage to host epithelium (Ferrara and others 1996). In human GVHD, some reports highlight a correlation between the level of macrophage infiltration and clinical severity and outcome (Nishiwaki and others 2014; Terakura and others 2015); and monocyte-derived macrophages are known to mediate pathological damage to skin explants independent of T cell involvement (Jardine and others 2020). In this study, we found that donor macrophages, but not CTLs, both of which had been induced by host-type lymphoma cells in the host undergoing GVHD, exhibited a GVHD activity against host skin epithelia in mice (Fig. 2), demonstrating the effector cells for GVHD to be macrophage (Fig. 5).

Possible mechanisms of BM GVHD and the GVL effect.
In human GVHD, the principal organs affected by GVHD are the skin (81%: not 100%), gastrointestinal tract (54%), and liver (50%; Martin and others 1991), implying BM GVHD as the base. Moreover, Divito and others (2020) demonstrated that T cells in the host skin induce GVHD-like dermatitis independent of donor T cells in human skin-grafted mice. Hematopoietic stem cell transplantation protocols include a conditioning regimen that is myelosuppressive, to deplete host stem cells and make space for donor stem cell engraftment, suggesting fixation of i.v. injected donor cells in the host BM to induce BM GVHD. Consistently, our mice undergoing BM GVHD (Tables 1 and 2) did not develop obvious skin GVHD (Fig. 1); and no apoptotic damage was seen in the epithelia of their skin, lung, or intestine (Fig. 3). In murine models of GVHD, Shono and others (2010) previously reported the early destruction of BM osteoblasts.
Three distinct phases (ie, tissue damage, donor T cell activation, and target cell apoptosis) of GVHD provide complex cellular interactions and inflammatory cascades (Ferrara and Deeg 1991). Therefore, there have been conflicting reports as to the role of cytokines (eg, Th1 and Th2) in the pathobiology of GVHD (Henden and Hill 2015). In this study, we demonstrated that IFN-γ was essential for the BM GVHD (Table 2) and that both CD4+ and CD8+ T cells were involved not only in the GVHD macrophage induction as IFN-γ producers (Yoneda and Yoshida 1998) but also in the GVL lymphocyte induction as IL-2-producing CD4+ T cells or CD8+ effector cells (Fig. 5). Consistently, allograft rejection is IL-2 independent (Steiger and others 1995; Dai and others 1998), but the allo-CTL induction in the primary response is IL-2 dependent (Suzuki and others 1992).
Ruxolitinib, a selective JAK1/2 inhibitor, has recently gained favor as a second-line approach in patients with steroid-refractory GVHD (Spoerl and others 2014). The JAK/STAT pathway is central not only to IFN-γ/IFN-γ receptor signaling but also to T cell activation, differentiation, and survival. In contrast, we here demonstrated that graft T cell activation rather than inhibition skipped GVHD after BM transplantation. Similarly, despite advances in immune suppression, allograft loss due to rejection remains a formidable clinical problem (Eisen 2014). Allograft rejection is a physiological response of the host to allografts; and the infiltration of macrophage cytotoxic against CTL-resistant allografts (eg, Meth A cells and skin epithelia) into the rejection site precedes by several days the infiltration of CTLs cytotoxic against donor-type lymphoid cells (Yoshida and others 1997a; Yamamoto and others 1998). Therefore, it is anticipated that allografts might become tolerated by simultaneous i.v. injection of recipient anti-donor CTLs.
Conclusions
Macrophage-induced BM GVHD and CTL-mediated GVL effect was a set (Th1: IFN-γ/IL-2) response of the graft to hosts and leukemia cells, respectively; and i.v. infusion of graft anti-host CTLs along with allogeneic BM skipped macrophage-induced GVHD. This approach can be applicable to the clinical setting.
Footnotes
Acknowledgments
The authors thank Drs. D. Hashimoto and K. Ikeda for critical reading of this article and T. Ueno, R. Oide, and K. Minami for their skillful technical assistance.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported, in part, by the Mori Memorial Research fund from Osaka Medical College and by the program Grants-in-Aid for Scientific Research (C; Grant No. 20591538) from the Ministry of Education, Science, Culture, and Sports, Japan.