
Abstract
Interferons (IFNs) are divided into 3 types (type I, type II, and type III) on the basis of sequence homology and functional properties. Recombinant IFNs have been approved by regulatory agencies in many countries for clinical treatment of hepatitis B, hepatitis C, and other diseases; these IFNs are mainly produced in microorganisms and mammalian cell systems. However, there are serious obstacles to the production of recombinant IFNs in microorganism systems; for example, the recombinant IFN may have different glycosylation patterns from the native protein, be present in insoluble inclusion bodies, be contaminated with impurities such as endotoxins and nucleic acids, have a short half-life in human blood, and incur high production costs. Some medicinal proteins have been successfully expressed in plants and used in clinical applications, suggesting that plants may also be a good system for IFN expression. However, there are still many technical problems that need to be addressed before the clinical application of plant-expressed IFNs, such as increasing the amount of recombinant protein expression and ensuring that the IFN is modified with the correct type of glycosylation. In this article, we review the classification of IFNs, their roles in antiviral signal transduction pathways, their clinical applications, and their expression in plant systems.
Background
In 1957, Isaacs and Lindenmann discovered interferon (IFN) as a factor that protected cells from viral infection (Isaacs and Lindenmann 1957). IFNs have subsequently been shown to be proteins with antiviral activity that are secreted from cells in response to a variety of stimuli (Pestka and others 2004), and are only found in vertebrates (Takaoka and Yanai 2006). IFNs are part of the class II family of cytokines produced by monocytes and lymphocytes; members of this family are polypeptides 161–208 amino acids in length and share a conserved structure that comprised 6 α helices (Lazear and others 2019).
IFNs are a group of bioactive proteins with multiple functions, including activity against a broad spectrum of viruses and regulation of cell growth, differentiation, and immune function. In addition to antiviral activity, IFNs also have antitumor activity and immunomodulatory effects (Borden 2019; Fox and others 2020). Because of these activities, IFNs are widely used in the treatment of disease. For example, in the 1980s, recombinant IFN-α was approved by regulatory agencies in many countries for the clinical treatment of hepatitis B, hepatitis C, and other diseases (Radwanski and others 1987; Takaoka and Yanai 2006; Yeh and others 2019).
The IFN used in clinical applications is mainly obtained through genetic engineering of Escherichia coli and Chinese hamster ovary (CHO) cells. The IFN produced by microbial systems is not glycosylated, and there is a slight difference in biological activity from the natural protein (Tharmalingam and others 2008). A large dose is required to achieve a therapeutic effect, and there is a risk of developing neutralizing antibodies (Jakimovski and others 2018). Although high yields of IFNs produced by microorganisms and mammalian cells can be obtained, recombinant IFN has a short half-life in clinical applications, which may be caused by the lack of or incomplete glycosylation modification. At present, IFN is modified with polyethylene glycol (PEG) to increase its half-life, and patient sensitivity to PEG may still compromise the effectiveness and safety of the drug therapy (Ramon and others 2005; Lee and others 2007; Katla and others 2019; Yeh and others 2019).
The production of antibodies, vaccines, and other therapeutic proteins in plants (ie, biopharming) shows great promise because recombinant proteins offer crucial advantages, including rapid scalability, low cost, and significantly reduced risk of human contamination compared with other production systems (Moustafa and others 2016; Ghag and others 2021). Over the last few decades, significant progress has been made in overcoming several shortcomings of biopharming production systems and regulatory issues (biosafety and risk assessment), and biopharming reached a stage where it can compete with challenge from the established production technologies that use bacteria, yeast, and mammalian cells (Ghag and others 2021). Because some medicinal proteins expressed in plants have been successfully used in clinical applications (Yao and others 2015; Moustafa and others 2016), researchers have begun to express IFN in plants and have tried to increase the yield of plant-expressed IFN and extend its half-life in clinical applications (Castro and others 2021).
In this review, we first describe briefly the classification of IFNs and the antiviral signal transduction pathways in which IFNs are involved, and then focus on clinical applications of IFNs and expression of IFNs in plant expression systems.
IFN Family Members
IFNs are part of the class II cytokine family, members of which are 161–208 amino acid polypeptides that share a conserved structure composed of 6 α-helices (Lazear and others 2019). IFNs are divided into 3 types (type I, type II, and type III) on the basis of sequence homology and functional properties (Lazear and others 2019), and each type is divided into different subclasses (Table 1).
Interferon Family Members
Pseudogene.
Not found in humans.
IFN, interferon; IL, interleukin.
Type I IFNs include the IFN-α (alpha), IFN-β (beta), IFN-ɛ (epsilon), IFN-κ (kappa), IFN-ω (omega), IFN-ν (nu), IFN-τ (tau), IFN-δ (delta), IFN-ζ (zeta), and IFN-χ (chi) subtypes. All type I IFNs are principally expressed by innate immune cells such as monocytes, macrophages, and T cells (Negishi and others 2018), and induced in virally infected cells to confer an antiviral state in uninfected cells (Mesev and others 2019).
There are differences in type I IFN subtypes among different species. In humans, there are 13 subtypes of IFN-α, with IFN-α1 and IFN-α13 having identical amino acid sequences (Accession No. NP_076918.1) (Wu and others 2020). Human IFN-β exists as single protein subtype and is predominantly expressed by fibroblasts (Lazear and others 2019). IFN-ω is present in humans, mice, cats, and cattle, but genomic analysis shows that the IFN-ω gene is missing in dogs (Li and others 2017). Human IFN-ω genes include 4 pseudogenes and 1 full gene that is expressed in leukocytes, and human IFN-ω shares 62% amino acid sequence homology and similar functions with IFN-α, and 33% amino acid similarity with IFN-β (Li and others 2017). IFN-ν is a pseudogene located on human chromosome 9 (Krause and Pestka 2005). IFN-τ, IFN-δ, IFN-ζ, and IFN-χ have been identified in pigs, mice, cattle, and other ruminants, but are not found in humans and dogs (Khan and Nurjis 2012; Ealy and Wooldridge 2017; Guo and others 2020).
IFN-γ was originally discovered in a study of cell resistance to viral infections (Alspach and others 2019), and is now known to represent the only member of type II IFNs. IFN-γ plays critical roles not only in orchestrating both innate and adaptive immune responses against viruses, bacteria, and tumors, but also in promoting pathologic inflammatory processes (Lazear and others 2019). IFN-γ is mainly expressed in natural killer (NK) cells and T lymphocytes, and can eventually lead to an immune response composed of CD4+ T cells and CD8+ T cells (Negishi and others 2018; Stanifer and others 2019).
Type III IFN members include IFN-λ1 (lambda), IFN-λ2, IFN-λ3, and IFN-λ4 (Lazear and others 2019). In 2012, while designating the official names and gene symbols of IFN-λ4, the Human Genome Organization Gene Nomenclature Committee changed the official symbols of interleukin (IL)-29, IL-28A, and IL-28B to IFN-λ1, IFN-λ2, and IFN-λ3, respectively (Negishi and others 2018; Lazear and others 2019). These changes are consistent with the generally recognized fact that these cytokines mainly act as IFNs, rather than as ILs (Walter 2020). The proteins encoded by these 3 genes (IFN-λ1, IFN-λ2, and IFN-λ3) are highly similar (Lazear and others 2019). IFN-λ2 and IFN-λ3 share 96% amino acid similarity, and the noncoding sequences of these genes, including upstream sequences, are almost the same (O'Brien and others 2014).
IFN Signal Transduction
IFNs differ greatly in structure, receptor distribution, and tissue-specific biological activity, but all of them can induce antiviral signaling. The type I IFN signal in the human body can fight numerous viruses, such as the influenza virus (Wei and others 2019); the flaviviruses dengue virus, yellow fever virus, West Nile virus, and Zika virus (Ngono and Shresta 2018); the hepatitis viruses hepatitis B virus (HBV), hepatitis C virus (HCV), and hepatitis D virus (Shin and others 2016; Su and Kao 2017; Xia and others 2019; Zhang and Urban 2020); and severe acute respiratory syndrome coronavirus-2 (Sa and others 2020).
Each type of IFN has a distinct heterodimeric cell surface receptor. Although the structural similarity between type I IFN subtypes is limited, they all share the same heterodimeric receptor composed of the low-affinity receptor IFNAR1 and the high-affinity receptor IFNAR2 (Takaoka and Yanai 2006; Lazear and others 2019). Type II IFNs bind to the IFNGR receptor complex (composed of IFNGR1 and IFNGR2), and type III IFNs bind to the IFNLR receptor complex (composed of IFNLR1 and IL-10Rβ) (Lazear and others 2019; Mesev and others 2019).
Although type I and type III IFNs bind to different receptors, they activate similar signaling pathways and transcriptional responses (Fig. 1) (Mesev and others 2019). Both type I and type III IFNs can promote signaling in an autocrine (in the secreting cells) or paracrine (in bystander cells) manner to induce an IFN-mediated immune response (Stanifer and others 2019). Binding of type I and type III IFNs to their respective receptor complexes results in phosphorylation of Janus-activated kinase (JAK)1 and tyrosine kinase 2 (TYK2) by the cytoplasmic domains of the receptor subunits (Stanifer and others 2019). This triggers phosphorylation of the heterotrimeric transcription factor complex IFN-stimulated gene factor 3 (ISGF3), which is composed of phosphorylated STAT1 and STAT2 and IFN regulatory factor 9 (Teijaro 2016; Mesev and others 2019). Following phosphorylation, the signal transducer and activator of transcriptions (STATs) form various complexes that translocate into the nucleus, where they bind to IFN-stimulated response elements (ISREs) on the promoters of IFN-stimulated genes (ISGs), resulting in an immune response against the viral infection (Lazear and others 2019; Mesev and others 2019).
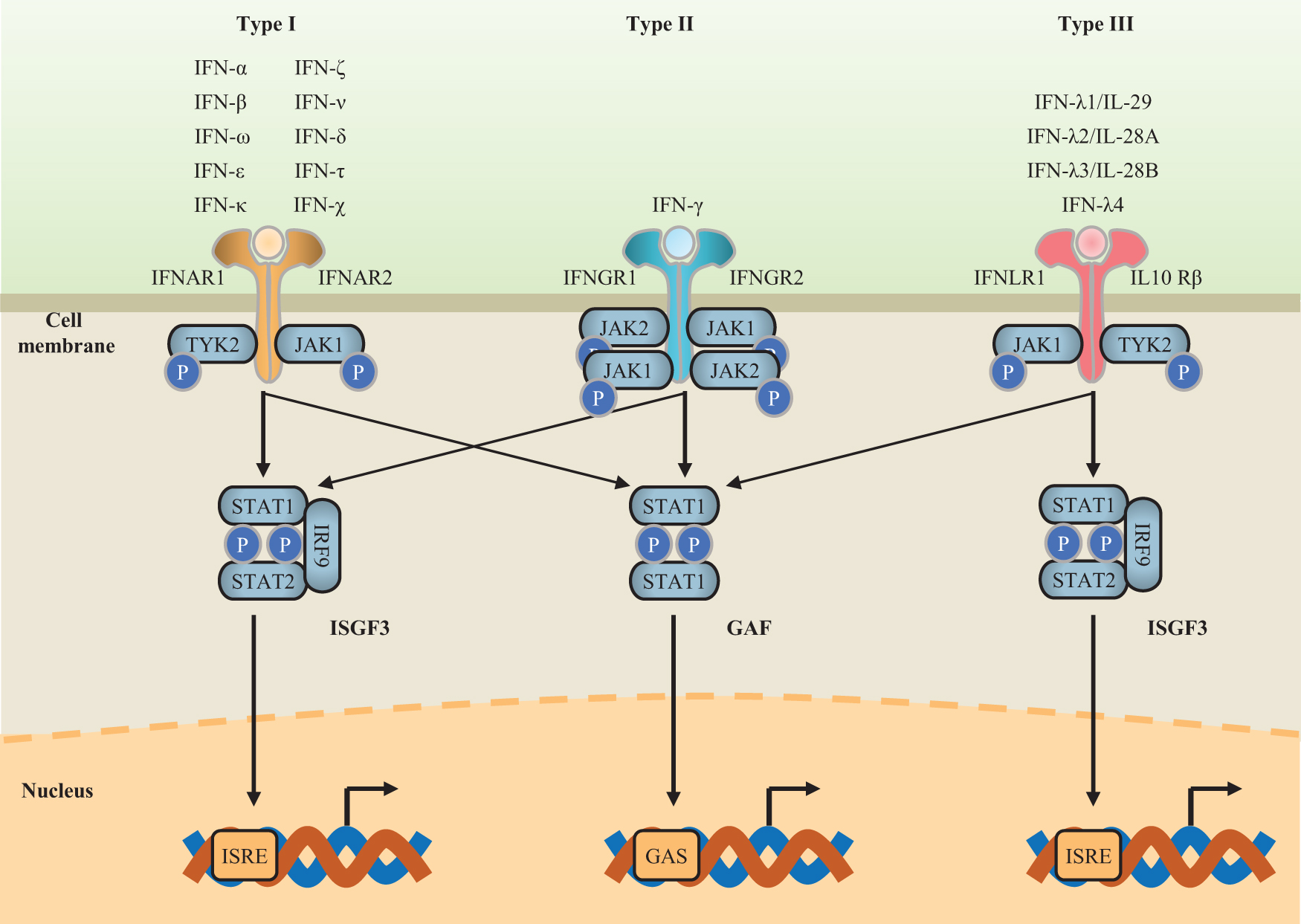
Schematic diagram of signal transduction pathways for the 3 types of IFNs. Type I IFNs bind to the IFNAR receptor complex (composed of IFNAR1 and IFNAR2); type II IFNs bind to the IFNGR receptor complex (composed of IFNGR1 and IFNGR2); type III IFNs bind to the IFNLR receptor complex (composed of IFNLR1 and IL-10Rβ) (Lazear and others 2019; Mesev and others 2019). Each type of IFN activates JAK or TYK phosphorylation after binding to its corresponding receptor. Then, the phosphorylation of the ISGF3 or GAF complex is triggered. Finally, the antiviral signal is transduced by activation of the transcription of ISGs through high-affinity binding of STAT complexes to ISRE- or GAS-containing DNA sequences in the nucleus (Negishi and others 2018). GAF, IFN-γ-activated factor; GAS, gamma-activated sequence; IFNs, interferons; IL, interleukin; IRF9, IFN regulatory factor 9; IRSE, IFN-stimulated response element; ISGF3, IFN-stimulated gene transcription factor 3; ISGs, IFN-stimulated genes; JAK, Janus-activated kinase; P, phosphate; STAT, signal transducer and activator of transcription; TYK2, tyrosine kinase 2. Color images are available online.
Type II IFN (IFN-γ) is best known as a key cytokine secreted during activated NK and T cell responses (Lee and Ashkar 2018). In the case of inflammation, IFN-γ induces a rapid response through JAK/STAT pathways (Negishi and others 2018). First, IFN-γ binds to a cell surface receptor (different from those bound by type I and type III IFNs) composed of IFNGR1 and IFNGR2 subunits, which in turn associates with JAK1 and JAK2 (Fig. 1). Then, the activation of these kinases leads to STAT1 homodimerization, nuclear translocation, and binding to gamma-activated sequences on DNA (Platanias 2005; Negishi and others 2018).
ISGF3 is also activated by IFN-γ signaling, although the activation is weak (Negishi and others 2018). It has also been shown that an effective IFN-γ signal depends on a weak type I IFN signal, which is caused by the low constitutive production of type I IFN (Green and others 2017).
IFNs Used in Clinical Treatment
The antiviral and immunomodulatory properties of type I and type II IFNs have been used clinically to control viral infection, enhance antigen presentation, and promote antitumor response (Table 2). Although preclinical experiments (Clinical trial registration, NCT01204762) have shown that type III IFNs may provide therapeutic benefits with reduced side effects during chronic viral infection, IFN-λ drugs have not been approved for use in humans (Phillips and others 2017; Lazear and others 2019).
Interferon Drugs Used in Clinical Treatment
IFN drugs were retrieved from the official websites of the FDA (
CHO, Chinese hamster ovary; FDA, Food and Drug Administration; HBV, hepatitis B virus; HCL, hairy cell leukemia; HCV, hepatitis C virus; HPV, human papilloma virus; MS, multiple sclerosis; NANB, non-A, non-B hepatitis; PEG, polyethylene glycol.
The Food and Drug Administration (FDA)-approved drugs Roferon-A (IFNα-2a) produced by Roche Pharmaceuticals and Intron A (IFNα-2b) produced by Merck Sharp & Dohme Corp. are both used clinically for treatment infections caused by viruses such as HBV and HCV. At present, long-acting IFNs (PEG-Intron and Pegasys) are mainly modified with PEG, which is nontoxic, nonimmunogenic, and nonpyrogenic; protein conjugation is achieved through chemical reaction of PEG with specific amino groups or sulfhydryl groups in IFNs (Hu and others 2018; Zhang and others 2020b). IFN-β-based products, including Avonex (IFN-β1a; Biogen), Rebif (IFN-β1a; EMD Serono), Betaseron (IFN-β1b; Bayer), Betaferon (IFN-β1b; Bayer), and Plegridy (PEG-IFN-β1a; Biogen), were the first major therapeutic class of drugs developed for use in multiple sclerosis (Arababadi and others 2010). The type II interferon IFN-γ is used to treat chronic granulomatous disease and liver fibrosis (Leiding and Holland 1993; Razaghi and others 2016; Xiang and others 2020; Zhang and others 2020a).
Most of these IFN drugs approved by the FDA are recombinant proteins expressed in E. coli or proteins modified with PEG. In China, the IFN drugs approved by the National Medical Products Administration include Anfulong (IFN-α2b; Tianjin Sinobioway) and Liyamei (IFN-α2b; Harbin Pharmaceutical Group), which are expressed in Pseudomonas putida, and Wanfuluo (IFN-α2b; Shanghai Tenry), which is expressed in yeast (Table 2).
In the 1980s, IFN was produced by exposing human T lymphocytes to mitogenic stimuli, or by translating mRNA in oocytes, which yielded lowly expressed proteins with low activity (Nagata and others 1980; Pestka 1986). With the development of molecular biology technology, E. coli has become one of the most commonly used systems for heterologous expression of foreign proteins, and several studies have reported the expression of IFN in E. coli. For example, Ahmed and others (2015) used the E. coli strain BL21DE3Plys to obtain a yield of 100 mg/L of recombinant human consensus interferon (rh-cIFN), and El-Baky and others (2015) obtained a yield of 270 mg/L for rh-cIFN-α using auto-induced expression in shake flask E. coli cultures.
However, there are serious obstacles to the production of recombinant IFN in microorganism systems, including the lack of or incorrect glycosylation, the production of insoluble inclusion bodies, contamination with impurities such as endotoxins and nucleic acids, and high production costs; furthermore, the resulting recombinant protein may have a short half-life in human blood (Hooker and James 1998; Razaghi and others 2016). Researchers have thus used different expression systems, such as plant expression systems, in an attempt to solve this problem.
IFN Expression in Plant Systems
Modern biotechnology exploits to overcome certain precincts normally seen during the commercial productions and pharmaceutical proteins from microbial system to mammalian cell culture in terms of cost, scalability, safety, and authenticity (Kopertekh and Schiemann 2019). In 1986, human growth hormone produced in tobacco and sunflower was the first plant-derived recombinant therapeutic protein (Barta and others 1986). Subsequently, several plant-derived pharmaceuticals were placed in preclinical or clinical trials. Taliglucerase alfa, a carrot cell-expressed recombinant protein used as a glucocerebrosidase replacement therapy for Gaucher disease, was approved by the FDA in 2012 after Phase III clinical trials were completed (Clinical trial registration, NCT00376168) (Zimran and others 2011). Recombinant glucocerebrosidase (T. alfa) produced in carrot cells has entered clinical applications, and the production of recombinant drug proteins in plants has entered a new stage (Hefferon 2013; Twyman and others 2013). ZMapp, which is a humanized monoclonal antibody produced in tobacco by the Mapp Biopharmaceutical Company (Joffe 2014; Kanapathipillai 2014; Davey and others 2016), has been used in a clinical trial for the treatment of Ebola virus infection (Clinical trial registration, NCT02363322). Due to the absence of fucose on N-glycans attached to the Fc domains, the plant-produced anti-Ebola mAbs elicited a significantly stronger antibody-dependent cellular cytotoxicity than the analogous anti-Ebola mAbs with fucosylated N-glycans, which were produced in a mammalian CHO cell line (Budzianowski 2015; Davidson and others 2015).
There are also some commercialized plant-derived proteins used as reagents in scientific research, such as bovine aprotinin (Cat. No. A6103) and tissue factor proteinase inhibitor-II (Cat. No. T5077) produced by Sigma-Aldrich, which are expressed in tobacco (
There are a variety of plants with glycosylation systems that have been modified to be as similar as possible to mammalian glycosylation systems, thereby reducing the risk of allergic reactions (Chen and others 2005; Arntzen 2015; Yao and others 2015). It has been reported that the lack of carbohydrate chains in IFN-β leads to reduced solubility and protein aggregation (Spearman and others 2005). Native hIFN-γ has 2 N-glycosylated sites, asparagine N25 (modified with fucosylated complex-type oligosaccharides) and N97 (modified with hybrid and high-mannose structures), which are known to be essential for resistance to cathepsin G, granulocyte proteases, plasmin, and purified elastase (Razaghi and others 2016; Wang and others 2020a). Unlike bacteria, plants do not require any other adaptations for heterologous expression of eukaryotic genes, especially intron excision (Yuan and Grotewold 2015). Also, plants, which are in cultivable in various platforms (open field, greenhouses, or bioreactors), have the potential to produce different types of therapeutic proteins with reduced risks of contamination with human and animal pathogens, and so, they are a relatively safe source of recombinant proteins for medical and veterinary purposes (Fischer and others 2012; Krenek and others 2015).
The current strategy for the expression of medicinal proteins including IFN in plant systems is stable genetic transformation and transient expression using Agrobacterium (Heidari-Japelaghi and others 2020b). The plants used to express IFN are mainly tobacco, potatoes, and rice (Table 3). A commonly used strategy to improve the expression of IFN in plants is to add elements that facilitate gene transcription or expression, such as an untranslated region, Kozak sequence, signal peptide, strong promoter, and fusion tag, in the plant expression vector (Ahangarzadeh and others 2012; Khan and Nurjis 2012; Heidari-Japelaghi and others 2020a, 2020b). High-level expression of IFNs is a key step toward human clinical trials and commercialization. In the current plant expression systems, the amount of IFN expressed ranges from 0.42% to 22.48% of the total soluble protein (TSP) (Table 3). Arlen and others (2007) expressed recombinant human IFN-α2b in tobacco chloroplasts with a yield of 3 mg/g fresh weight or 20% TSP, and transgenic lines were grown in the field after obtaining approval from the US Department of Agriculture Animal and Plant Health Inspection Service. Human IFN-γ fused with the elastin-like polypeptide sequence was transiently expressed in tobacco using Agrobacterium infiltration, with a yield of 22.48% TSP (Heidari-Japelaghi and others 2019).
Expression of Human Recombinant Interferons in Plants
FW, fresh weight; NC, not calculated; TSP, total soluble protein; UTR, untranslated region.
Modification of IFN with PEG prolongs its half-life in the human body, but at the expense of greatly decreased bioactivity (Ramon and others 2005; Lee and others 2007; Yeh and others 2019). Thus, researchers have focused on the improvement of glycosylation to improve the half-life of recombinant proteins. Xu and others (2007) found that expression of a chimeric protein composed of human IFN-α2b and arabinogalactan protein (AGP) (IFN-α2b-AGP) in tobacco cells increased the in vivo serum half-life of recombinant IFN-α2b up to 13-fold, while the biological activity remained similar to that of native human IFN-α2b. IFN-α2b-AGP was not immunogenic when injected into mice (Xu and others 2007). Plant-expressed IFN has many advantages; however, there are still many technical problems that need to be overcome to facilitate its use in clinical applications; these include the low amount of recombinant protein expression and incorrect glycosylation (Table 4).
Differences Between Interferons Expressed in Different Systems
Plant virus expression vectors have been designed to express vaccine epitopes as well as full therapeutic proteins such as antibodies, therapeutic proteins, and other biopharmaceutical products in plant tissue both safely and effectively (Hefferon 2012). There are few reports of IFN expression using viral-based plant expression systems, which may be due to the antiviral activity of IFN. For example, after tobacco mosaic virus (TMV) infection, chicken IFN-α transgenic tobacco showed delayed onset and reduction of symptoms, indicating that transgenic tobacco had antiviral activity, and that avian IFN can inhibit the proliferation of the plant virus TMV (Song and others 2010). The engineered vector zucchini yellow mosaic virus was used to express hIFN-γ in pumpkin plants, ∼1–1.2 mg hIFN-γ per 100 g of tissue was obtained from the leaves 2 weeks after inoculation (Hosseini and others 2013). When an exogenous IFN gene is introduced into the plant, it also affects the growth and development of the plant. After treating tobacco and wheat with human IFN, Kulaeva and others (1992) found that the activity of plant root mitogens increased, and there was evidence that IFN promoted root cell division and plant growth and development.
Conclusions
With the development of the pharmaceutical market, the clinical requirements for IFNs are becoming more and more refined, and recombinant IFN with glycosylation modification is needed to increase its half-life and reduce side effects. There are many challenges to the use of plant expression systems, such as low yield, the differences between plant and animal glycosylation, and purification and downstream processing barriers, but plant expression of human pharmaceutical IFN proteins still represents a promising approach for low-cost and large-scale production. The large-scale production and purification of IFNs in Good Manufacturing Practice facilities should lead to more human clinical trials and further advances in this field.
Footnotes
Authors' Contributions
L.C. analyzed the literature and wrote the article. The other authors read and approved the final article.
Acknowledgment
The authors gratefully acknowledge the financial support from the Guizhou Tobacco Company Project (Grant No. 2020XM04).
Consent for Publication
All authors approve the article for publication.
Availability of Data and Materials
Data and materials related to this work are available upon request.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported by the Guizhou Tobacco Company Project (Grant No. 2020XM04) and Guizhou province Technology R&D Project (Grant No. [2019]2401).