
Abstract
Rheumatoid arthritis (RA) is a systemic immune disease. Rho family GTPase 3 (RND3) has been reported to play an important role in inflammatory diseases. In this study, the expression of RND3 in RA was analyzed by gene chips. After RND3 was overexpressed, cell counting kit-8 assay was to detect the viability of fibroblast-like synovial cells (RA-FLSs). Transwell assays were to appraise the migratory and invasive capacities of RA-FLSs. Enzyme-linked immunosorbent assay (ELISA) and Western blot analysis were to estimate inflammatory response. In addition, MMP3 and MMP9 levels were also tested by ELISA analysis. After forkhead box M1 (FOXM1) was overexpressed, RND3 expression was detected by Western blot. The transcriptional relationship between FOXM1 and RND3 was predicted by HumanTFDB and JASPAR databases. Luciferase reporter and chromatin immunoprecipitation assays verified the binding ability of FOXM1 and RND3. The role of FOXM1/RND3 axis in RA was detected again by functional experiments. Western blot detected the expression of Rho/ROCK pathway-related proteins. RND3 expression was downregulated in RA. Overexpression of RND3 reduced the proliferation, migration, invasion, and inflammation of RA-FLSs. RND3 was inhibited by FOXM1 transcription, and upregulated FOXM1 reduced the inhibitory effect of RND3 overexpression on cell growth and inflammation, which might be associated with the Rho/ROCK pathway. RND3 transcriptionally regulated by FOXM1 inhibited the migration and inflammation of RA-FLSs in RA through the Rho/ROCK pathway.
Introduction
Rheumatoid arthritis (RA) is a systemic autoimmune disease characterized by a persistent inflammatory response of the synovial membrane, resulting in synovial hyperplasia and eventual destruction of articular cartilage (Littlejohn and Monrad 2018). RA has been confirmed as a common disease with an average incidence of 1% worldwide (Alamanos and others 2006). It is widely considered that joint deformities and dysfunction in RA are becoming more severe and affect the patient's quality of life over time.
Fibroblast-like synovial cells (RA-FLSs) play an important role in the development of RA. RA-FLSs are cells that proliferate similar to tumor cells, and their enhanced activity is the key cause of abnormal synovial hyperplasia in RA patients (Yang and others 2017). In addition, during the process of RA, RA-FLSs are capable of releasing a large number of inflammatory factors, including tumor necrosis factor alpha (TNF-α), interleukin (IL)-1β and IL-6, and trigger inflammatory response, which may further activate the proliferation of RA-FLSs and aggravate cartilage damage (Kun and others 2020). Therefore, inhibiting the inflammatory response and abnormal proliferation and migration of RA-FLSs may be an important means for the treatment of RA.
Rho family GTPase 3 (RND3, also known as RhoE) is a member of the GTPase protein superfamily, which is involved in the metastasis and development of a variety of tumors (Zhang and others 2018; Garg and others 2020; Jiang and others 2020). RND3 is considered as an antagonist of Rho activity and downregulation of RND3 in lung fibroblasts can enhance RhoA activity and drive the fibrosis phenotype (Monaghan-Benson and others 2018). Knockdown of RND3 can promote the proliferation of Tenon cystic fibroblasts and inhibit the apoptosis of fibroblasts (Tong and others 2014; Sun and others 2015).
In addition, previous report has shown that RND3 can inhibit nuclear factor kappa B (NF-κB) activity and inflammation while improving cardiac function and survival rate (Dai and others 2019). Inhibition of NF-κB activation and production of pro-inflammatory cytokines by blocking Rho kinase can ultimately inhibit RA synovitis (He and others 2008). Also, through reducing the level of RhoA, the invasion ability of RA-FLS can be inhibited, thus providing help for the treatment of RA (Laragione and others 2019). Therefore, it is speculated that RND3 may be involved in the development of RA.
HumanTFDB database (
In this article, we examined the role of RND3/FOXM1 in the inflammatory response and abnormal migration of RA-FLSs and discussed the latent regulatory mechanism, which might provide a theoretical basis for the treatment of RA.
Materials and Methods
Microchips
The expression level of RND3 was analyzed by GSE55235, GSE1919, and GSE55457 microchips.
Database
HumanTFDB (
Cell culture
Synovial fibroblasts from RA RA-FLSs were obtained from the American Type Culture Collection (Manassas, VA) and Human fibroblast-like synoviocytes were obtained from Shanghai Cell Bank of the Chinese Academy of Sciences (Shanghai, China) and grown in Dulbecco's modified Eagle's medium (DMEM) with 10% fetal bovine serum (FBS, all from Gibco) plus 1% antibiotics (penicillin/streptomycin; Invitrogen) in a humidified incubator of 95% CO2 at 37°C.
Cell transfection
RA-FLSs were plated into 6-well plates at a density of 1 × 106 cell/well. RND3 and FOXM1 overexpression plasmid (Ov-RND3 and Ov-FOXM1) and their empty vector (Ov-NC) were obtained from Shanghai GeneChem Co. Ltd., and were then transfected into cells at a concentration of 20 nM using Lipofectamine® 3000 (Invitrogen; Thermo Fisher Scientific, Inc.) according to the manufacturer's instructions. The transfection efficiency was verified by reverse transcription-quantitative polymerase chain reaction (RT-qPCR) and Western blot analyses after 48 h.
Cell counting kit-8 assay
RA-FLSs (5 × 103 cells/well) were put into 96-well plates under the condition of 37°C. After incubation for 24, 48, and 72 h, 10 μL cell counting kit-8 (CCK-8) solution (Abcam, Cambridge, UK) was supplemented to cells. Cell proliferation was measured by detection of OD values at 450 nm under a microplate reader (Bio-Rad, Hercules, CA).
Transwell
Cell migration and invasion assays were performed in the Boyden chamber method using a filter with pore size of 8.0 μm (Transwell, Corning Labware Products). For cell migration, transfected RA-FLSs (8 × 104 cells/mL) were resuspended in serum-free medium in the upper well. Additional 600 μL DMEM with 10% FBS were placed as a chemoattractant in the lower well. After incubation for 12 h, a cotton swab was used to remove the nonmigrating cells on the filter's upper surface. The migrated cells to the lower side of the filter were fixed in 4% paraformaldehyde (Boster, China) for 20 min and stained with crystal violet (KeyGEN BioTECH) for another 20 min. Chemotaxis was quantified using an optical microscope to count the stained cells.
For the cell invasion assay, the upper chamber wells were coated individually with 30 μL diluted BD Matrigel Basement Membrane Matrix (BD Biosciences). The matrigel was diluted to 1:10 with DMEM. After air-drying at 4°C and concretion of matrigel, transfected RA-FLSs resuspended in serum-free medium (1 × 105 cells/mL) were seeded in the upper well. Six hundred microliters DMEM with 10% FBS was added to the lower well. After 24 h of culture under the condition of 37°C and 5% CO2, the mean number of migrated and invaded cells was recorded in 5 randomly selected fields at 50 × magnification for each assay.
Enzyme-linked immunosorbent assay
MMP9 (
Reverse transcription-quantitative polymerase chain reaction
Total RNA were isolated using TRIzol reagent (Invitrogen). Reverse transcription was performed using RevertAidTM First Strand cDNA Synthesis Kit (Fermentas, Ottawa Canada). The random RT primer 50–(dN) 9-30 was used for the RND3, FOXM1, and glyceraldehyde 3-phosphate dehydrogenase (GAPDH) genes. The PCR primers were designed using a master mix (SABI SYBR Green Master Mix; Invitrogen). All the steps followed the manufacturer's instructions. All mRNA quantification data were normalized to GAPDH. The data were processed using 2−ΔΔCt methods (Livak and Schmittgen 2001).
Western blot
Ice-cold lysis buffer (0.5 mL/well; Beyotime) was added to the cells and then the cells were rinsed after incubation for 30 min on ice. Protein concentrations were determined by BCA kit (Pierce, Rockford, IL). Thirty micrograms protein was separated by 12% sodium dodecyl sulfate (SDS)-polyacrylamide gel electrophoresis and transferred to polyvinylidene fluoride membranes. The membranes were probed at 4°C overnight with primary antibodies diluted to 1:1,000 after blocked with 5% nonfat milk. Next day, the membranes were incubated with the appropriate secondary antibodies (1:5,000; Abcam) for 1 h at room temperature. Immunoreactive bands were visualized by enhanced chemiluminescence reaction (Amersham Pharmacia Biotech, Little Chalfont, UK) and a semiquantitative analysis was conducted using ImageJ software (version 1.8.0; National Institutes of Health).
Dual-luciferase reporter assay
With the aid of Lipofectamine 2000 reagent (Invitrogen), RA-FLSs were co-transfected with the wild-type (WT) or mutant (MUT) region of RND3 (WT or MUT-RND3, respectively) promoter and Ov-FOXM1 or Ov-NC for 48 h. These fusion plasmids were severally transfected into cells. After transfection, the relative luciferase activity was determined after 48 h using the Dual-Luciferase Reporter Assay kit (Promega Corporation), according to the manufacturer's instructions. Renilla luciferase activity was used to normalize the firefly luciferase activity.
Chromatin immunoprecipitation assay
After the cells were scraped and lysed in 100 μL SDS lysis buffer (Beyotime Institute of Biotechnology), cells were fixed with 1% formaldehyde for 15 min at 37°C. Extracted proteins were immunoprecipitated using 5 μL FOXM1 antibody (1:1,000; Abcam) or control rabbit IgG (1:1,000; Abcam) for 12 h at 4°C after purified by soluble material. Subsequently, 20 μL protein A + G agarose beads (Beyotime Institute of Biotechnology) was added and incubated for another 4 h at 4°C. Then the collected agarose beads were washed 5 times with 100 μL phosphate-buffered saline and denatured by boiling for 5 min. The relative expression levels were determined through RT-qPCR according to the manufacturer's instructions.
Statistical analyses
Student's t-test and one-way analysis of variance (ANOVA) test were used for data analysis. Statistical analyses were performed using GraphPad Prism 6.0. P values <0.05 were considered as statistically significant.
Results
RND3 expression is downregulated in RA synovium
First, the expression level of RDN3 in RA was analyzed by GSE55235, GSE1919, and GSE55457 microchips. It was found that the expression of RND3 in RA synovium was significantly decreased compared with normal controls (Fig. 1A–C).
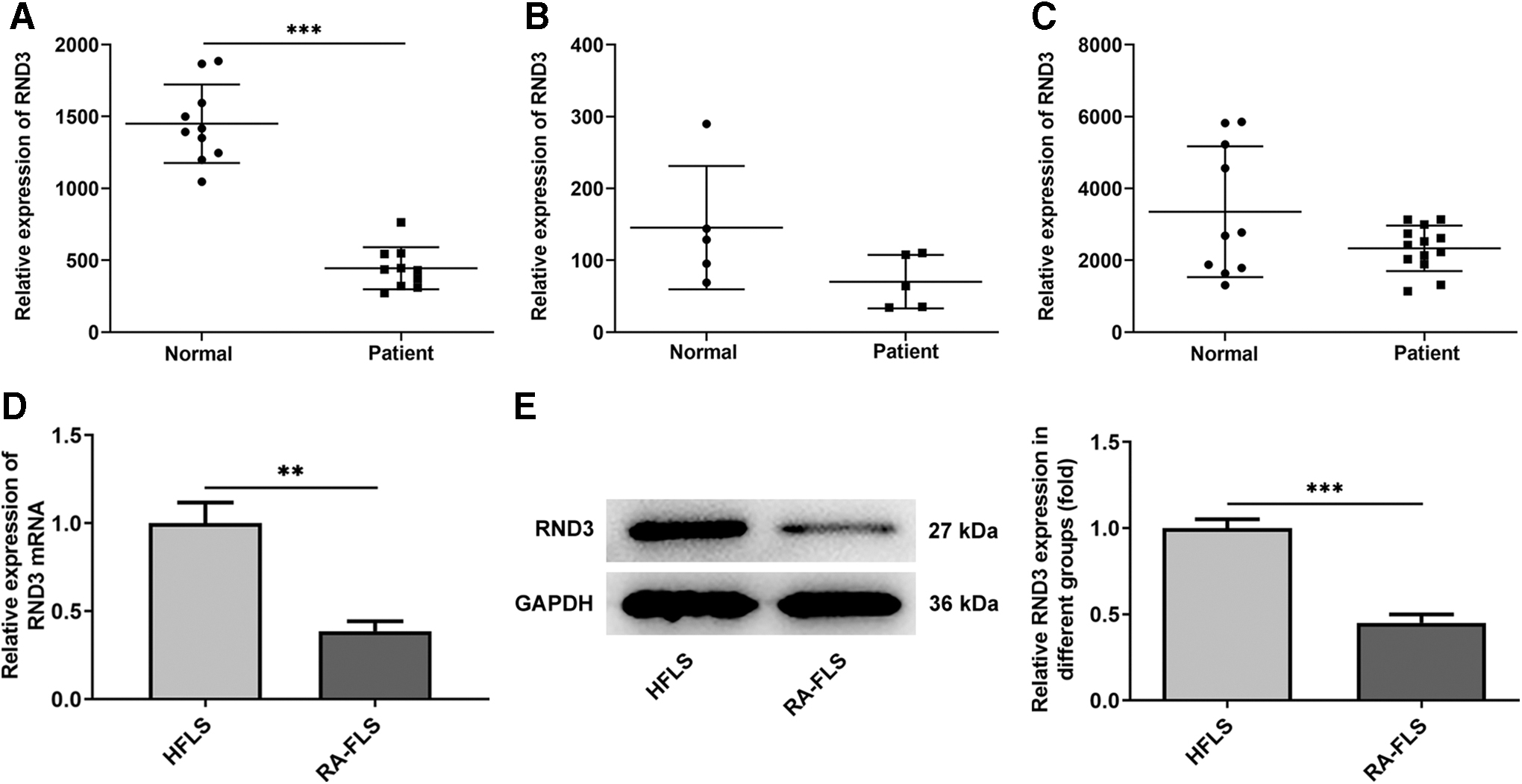
RND3 expression is downregulated in RA synovium. The expression level of RND3 was analyzed by GSE55235
Overexpression of RND3 reduces the viability, migration, and invasion of RA-FLSs
Subsequently, overexpression vector of RND3 was constructed and RT-qPCR and Western blot were used to detect the overexpression efficiency. The results showed that the expression of RND3 was significantly increased in the Oe-RND3 group compared with the Oe-NC group (Fig. 2A, B). CCK8 results showed that the cell viability of Oe-RND3 group was significantly decreased compared with Oe-NC group (Fig. 2C). Transwell results showed that the migration (Fig. 3A, B) and invasion (Fig. 3C, D) of RA-FLSs were significantly decreased after overexpression of RND3 compared with those of RA-FLSs transfected with Oe-NC. ELISA was used to detect the expression of MMP9 and MMP3 associated with migration and invasion, and the results showed that the expression of MMP9 and MMP3 was significantly decreased after overexpression of RND3 (Fig. 3E, F).
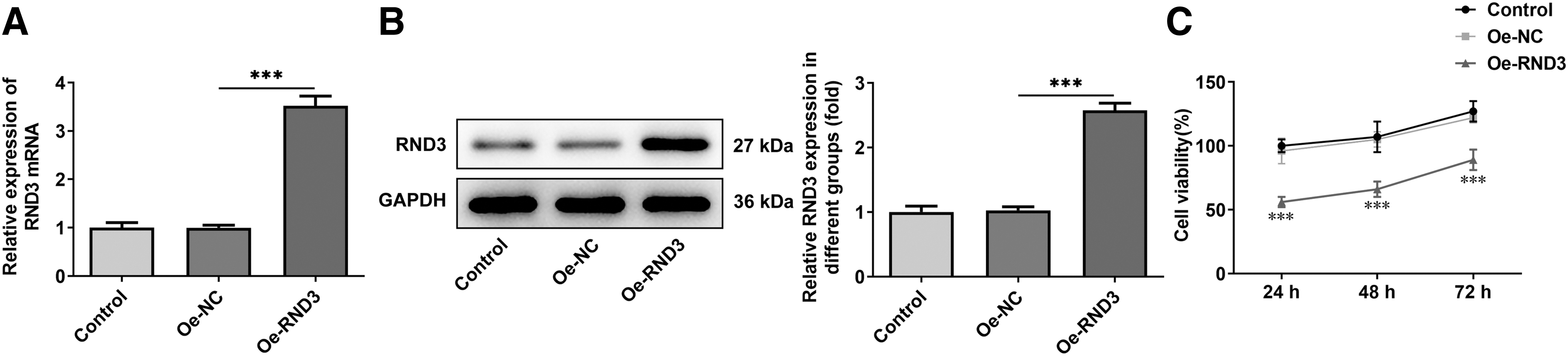
Overexpression of RND3 reduces the viability of RA-FLSs.
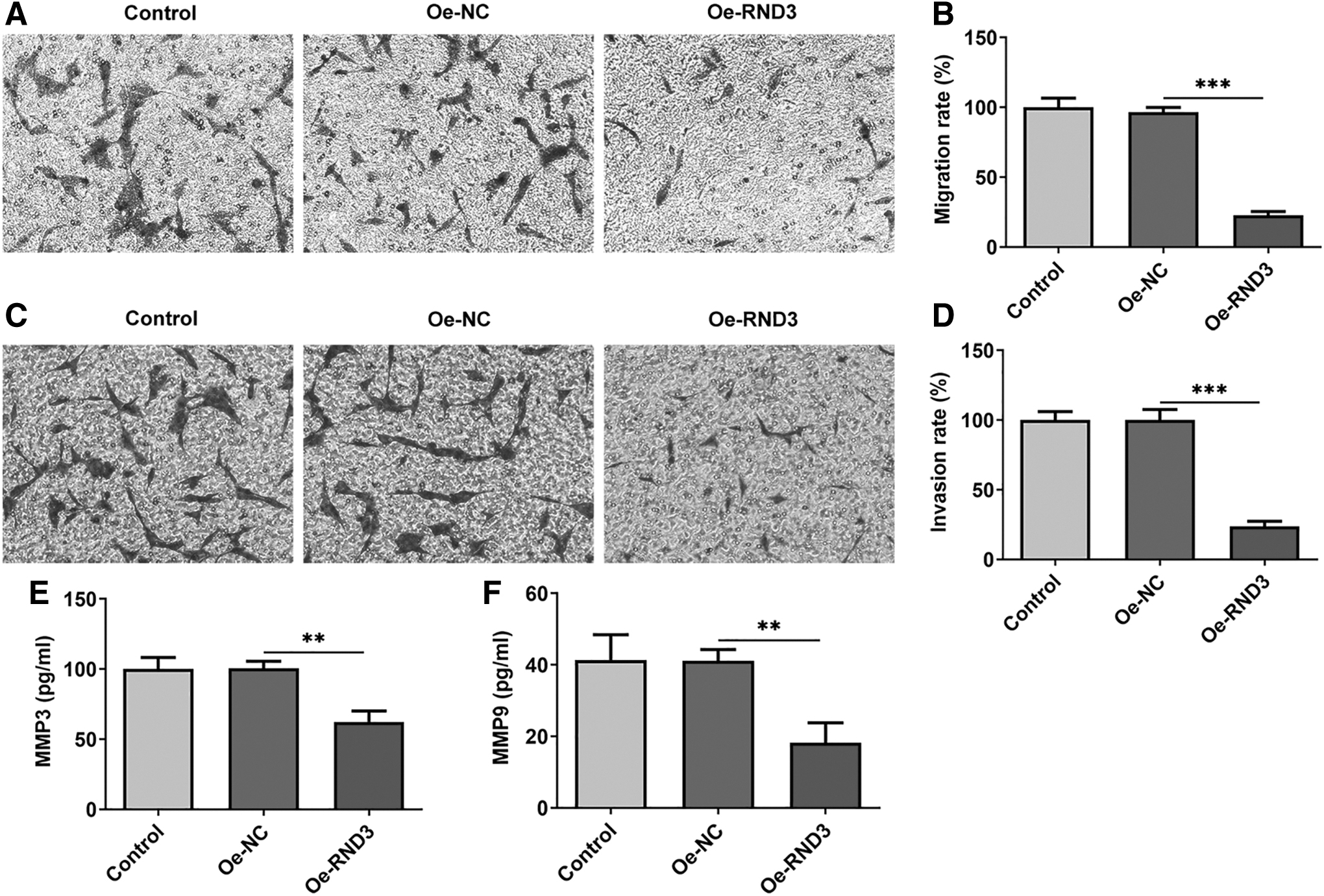
Overexpression of RND3 reduces the migration and invasion levels of RA-FLSs.
Overexpression of RND3 reduces the inflammatory response of RA-FLSs
To assess the impacts of RND3 on inflammation of RA-FLSs, ELISA assay and Western blot were to detect the levels and expression of inflammatory factors. The results of ELISA assay showed that the levels of IL-6, TNF-α, and IL-1β were significantly decreased after overexpression of RND3 compared with Oe-NC (Fig. 4A), and the expression of inflammation-related proteins iNOS, COX-2, and p-NF-κB was also significantly decreased after overexpression of RND3 (Fig. 4B).
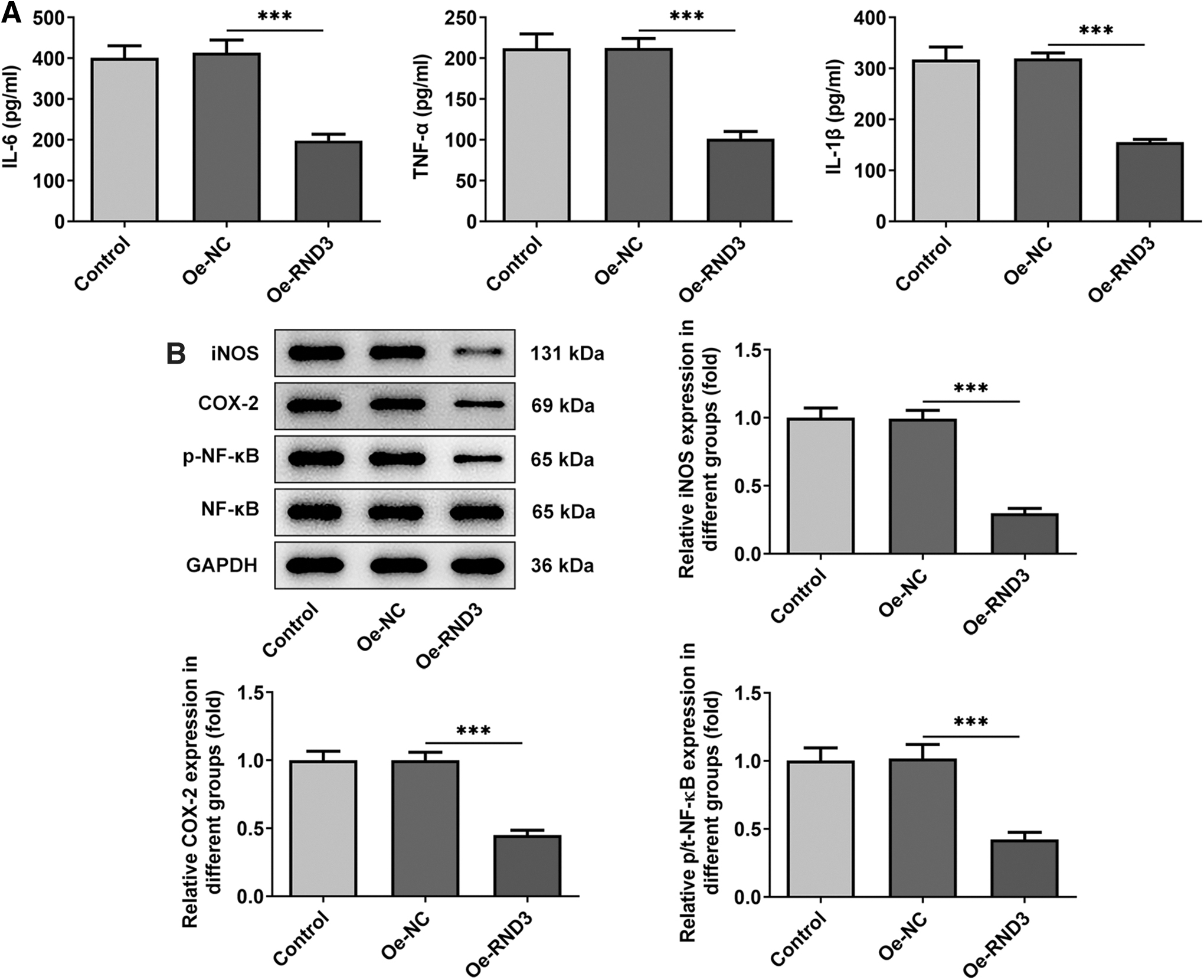
Overexpression of RND3 reduces the inflammatory response of RA-FLSs.
RND3 is transcriptionally inhibited by FOXM1 in RA-FLSs
After FOXM1 overexpression vector was constructed, the expression level of FOXM1 in cells was detected by RT-qPCR and Western blot. The results showed that FOXM1 was successfully overexpressed (Fig. 5A, B). Besides, after FOXM1 was overexpressed, the expression of RND3 was decreased significantly (Fig. 5C). Subsequently, FOXM1 was predicted to be a transcriptional regulator of RND3 (Fig. 5D) by means of JASPAR database. The binding affinity between FOXM1 and RND3 promoter was verified by luciferase reporter and chromatin immunoprecipitation assays (Fig. 5E, F). These results indicated that RND3 was transcriptionally inhibited by FOXM1 in RA-FLSs.

RND3 is inhibited by FOXM1 transcription in RA-FLSs.
Upregulation of FOXM1 reduces the inhibitory effect of overexpression of RND3 on the growth and inflammation of RA-FLSs
Then the cells were divided into 5 groups: control, Oe-NC, Oe-RND3, Oe-RND3+Oe-NC, and Oe-RND3+Oe-FOXM1. CCK8 results showed that cell viability in Oe-RND3+Oe-FOXM1 group was significantly increased compared with that in Oe-RND3+Oe-NC group (Fig. 6A). Transwell results showed that overexpression of FOXM1 could reverse the inhibitory effect of overexpression of RDN3 on the invasion and migration of RA-FLSs (Fig. 6B–E). ELISA results showed that the expression of MMP9 and MMP3 in Oe-RND3+Oe-FOXM1 group was significantly increased compared with that in Oe-RND3+Oe-NC group (Fig. 6F).

Upregulation of FOXM1 reduces the inhibitory effect of overexpression of RND3 on the growth of RA-FLSs.
Subsequently, levels of cellular inflammation were detected. Compared with Oe-RND3+Oe-NC group, the expression levels of inflammatory cytokines IL-6, TNF-α, and IL-1β, and inflammatory proteins iNOS, COX-2, and p-NF-κB were significantly increased in Oe-RND3+Oe-FOXM1 group (Fig. 7A, B). These results suggested that upregulation of FOXM1 could reduce the inhibitory effect of RND3 overexpression on RA-FLS inflammation.
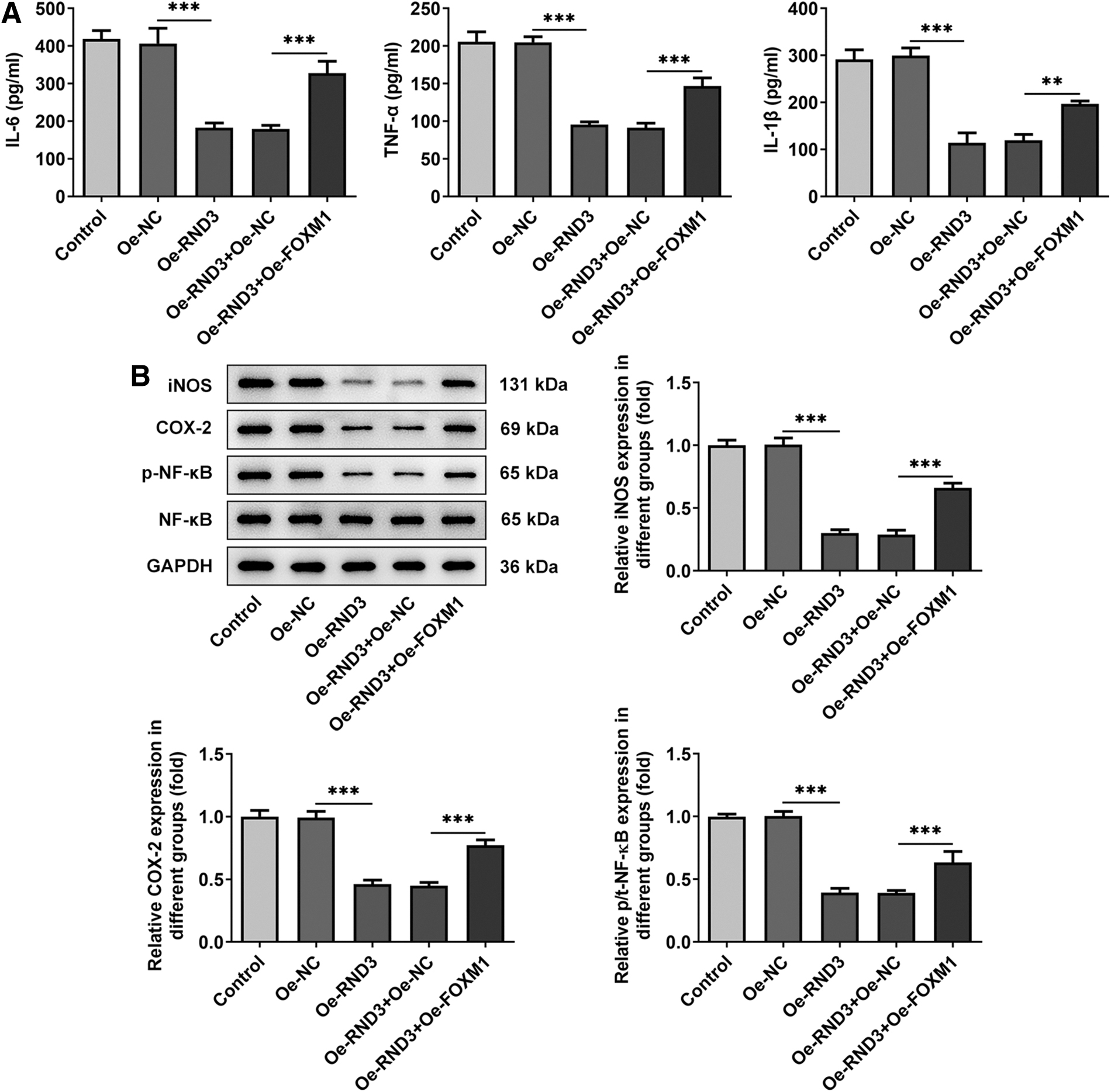
Upregulation of FOXM1 reduces the inhibitory effect of overexpression of RND3 on the inflammation of RA-FLSs.
Upregulation of FOXM1 reduces the inhibitory effect of overexpression of RND3 on Rho/ROCK pathway
More importantly, the Rho/ROCK signaling pathway was discovered to be dysregulated in RA-FLSs. Compared with Oe-NC group, the protein levels of Rho, ROCK1, and ROCK2 in Rho/ROCK signaling pathway was significantly decreased after overexpression of RND3. Compared with the Oe-RND3+Oe-NC group, the expression of Rho, ROCK1, and ROCK2 at protein level in the Oe-RND3+Oe-FOXM1 group were significantly increased (Fig. 8). It could be preliminarily concluded that upregulating FOXM1 reduced the inhibitory effect of overexpression of RND3 on Rho/ROCK pathway.

Upregulation of FOXM1 reduces the inhibitory effect of overexpression of RND3 on Rho/ROCK pathway. Western blot detected the expression of Rho/ROCK pathway-related proteins, n = 3. ***P < 0.001.
Discussion
It is well documented that the activation of FLSs play a leading role in the course of RA (Nygaard and Firestein 2020). Once the immune response is activated, a series of cytokines produced by RA-FLSs. As a consequence, the interaction between inflammation and endothelial cells is mediated, which may result in progressive destruction of bone and cartilage (Du and others 2019). A growing body of evidence has reported that RA-FLSs are frequently used in in vitro studies of RA (Bottini and others 2019; Li and others 2019; Najm and others 2020). Therefore, in this article, RA-FLS of RA patients were selected for in vitro exploration.
Tumor-like proliferative properties of the activated RA-FLSs lead to a large increase in the number of cells, which leads to the thickening of synovial tissues. The migration and invasion of thickened RA-FLSs are deemed as one of the most important biological characteristics in RA (Calmon-Hamaty and others 2015; Du and others 2019). In addition, immune inflammatory injury exists throughout the pathogenesis of RA. Owing to the disruption of the body's immune homeostasis, the synovial immune system of the joint is activated, and a large number of lymphocytes infiltrate, resulting in the imbalance of the environmental homeostasis in the joint cavity and accelerating the vicious cycle of local joint immunity and inflammation (Yang and others 2016; Nerurkar and others 2019). Therefore, in this article, the invasion, migration, and inflammatory injury of RA-FLSs were discussed so as to explore the regulation mechanism behind the pathogenesis of RA in vitro.
It was found that the expression of RND3 was significantly downregulated in RA patients by gene chip analysis. The role of RND3 in tumors has been widely studied, whereas the function of RND3 on RA has not been covered so far. It has been reported that RND3 is downregulated in fibrocytes and can regulate the proliferation of fibrocytes (Tong and others 2014; Monaghan-Benson and others 2018). In addition, RND3 is an active Rho antagonist.
Rho can lead to abnormal migration, invasion, and inflammatory response of RA-FLSs in RA (Arafa and others 2020; Singh and others 2021). Accordingly, it was speculated that RND3 played a regulatory role in the migration and inflammatory response of RA-FLSs in RA. Our study found that after overexpression of RND3 in RA-FLSs, the viability of RA-FLSs was significantly decreased, the invasion and migration abilities were significantly weakened, and the inflammatory response in RA-FLSs was also inhibited. These results indicated that overexpression of RND3 could inhibit the occurrence and development of RA.
It was predicted by HumanTFDB and JASPAR databases that FOXM1 could act as a transcription regulator of RND3, and it was also verified that RND3 was inhibited by FOXM1 transcription factor through experiments. A previous study has shown that inhibition of FOXM1 not only reduces joint bone destruction, but also reduces inflammatory response (Hasegawa and others 2019). Therefore, we made a conjecture that FOXM1 might play a role in RA through transcriptional regulation of RND3. Study has reported that FOXM1 expression in RA-FLSs was significantly increased compared with normal FLS (Fornes and others 2020). Therefore, in the experiment, RND3 and FOXM1 were overexpressed simultaneously to explore the mechanism. It was noticed that FOXM1 overexpression significantly reversed the inhibitory effect of RND3 overexpression on the viability, invasion, migration, and inflammatory response of RA-FLSs.
In the experiment, it was found that the Rho/ROCK signaling pathway was abnormally expressed. Study has shown that the expression of Rho/ROCK signaling pathway in arthritis is abnormally activated (Zhu and others 2016). Moreover, RND3 is able to reduce the activity of RhoA, and its phosphorylation status directly affects its ability to block Rho/ROCK signaling pathway (Madigan and others 2009). Our results suggested that overexpression of RND3 inhibited the activity of the Rho/ROCK signaling pathway, whereas further overexpression of FOXM1 reversed this inhibition. These results suggested that RND3 might play a role in RA by regulating the Rho/ROCK signaling pathway. In the following experiments, we will further verify our conclusions by adding pathway inhibitors.
There are some limitations in our article. Such as, one limitation of this paper is that we did not detect the RND3 gene knockout experiment, which will be further verified in future experiments. In our article, the relationship between cell viability, cell migration, and cell inflammation was not demonstrated. In addition, in this study, it was found that overexpression of RND3 inhibited NF-κB mediated inflammatory response, and literature review found that inhibition of NF-κB could effectively inhibit cell proliferation and migration (Liu and others 2018). So this is a complex process. We will further explore the relationship among cell viability, cell migration, and cell inflammation after overexpression of RND3 in future articles.
Conclusion
In this study, we concluded that RND3 transcriptionally regulated by FOXM1 inhibited the abnormal migration and inflammatory injury of synovial fibroblasts in RA through the Rho/ROCK pathway. Our article provided a theoretical basis for the treatment of RA.
Footnotes
Authors' Contribution
All authors read and approved the final version of the article.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported by grants from the Basic Research Project of Shenzhen Science and Technology Innovation Committee (No. JCYJ20210324120800001), the name of project is Study on the mechanism of thymic medullary fibroblasts in immune Tolerance Disorder of Rheumatoid Arthritis.