
Abstract
Some progress has been made in immunotherapy with chimeric antigen receptor (CAR)-T cells targeting NKG2D-NKG2DL with the purpose of eradicating solid tumors. Non-small cell lung cancer (NSCLC) has been shown to express NKG2DL. This study hence evaluated the therapeutic effect of NKG2D CAR-T cells on NSCLC. Accordingly, NKG2D CAR-T cells were obtained from diverse human autologous T cell sources. T cells from peripheral blood T lymphocytes of healthy volunteers (without NKG2D CAR insertion) were used as NT-T cells. Coculture of effector cells (CAR-T cells or NT-T cells) with target cells (NSCLC cells such as PC-9 or NCL-H460 cells) was performed at different ratios. The cytotoxicity of CAR-T cells was examined using lactate dehydrogenase assay kits. Murine xenograft assay was conducted to investigate the in vivo antitumor effect of CAR-T cells. Cytokines secreted from CAR-T cells were assessed by enzyme-linked immunosorbent assay. CAR-T cell infiltration into xenografts was observed through immunochemical assay. Based on the results, NKG2DL was highly expressed in NSCLC cells. Compared with NT-T cells, NKG2D CAR-T cells from different sources of T cells delivered stronger toxicity, and secreted more effector and memory function-related cytokines to NSCLC cells, and those from the peripheral blood of healthy donors (H-T cells) exhibited the strongest effect. Furthermore, compared with NT-T cells, H-T cells and NKG2D CAR-T cells from NSCLC patients' peripheral blood diminished tumor, improved survival, increased body weight and tumor-infiltrating capacity, and upregulated serum IFN-γ level in NOG mice. Collectively speaking, NKG2D CAR-T cells exhibit a robust effect on eradicating NSCLC in a NKG2DL-dependent manner, thus making themselves a promising therapeutic candidate for NSCLC patients.
Introduction
Lung cancer is a malignant solid tumor responsible for an estimated 2 million new cases and 1.76 million mortalities per year worldwide (Thai et al., 2021). The dismal outcomes caused by lung cancer often result from delayed diagnosis, and the diagnosis of this disease can be made only at an advanced stage (Nooreldeen and Bach, 2021). Lung cancer can be classified into distinct subtypes, namely small cell lung cancer (SCLC) and non-small cell lung cancer (NSCLC) (Howlader et al., 2020), the latter of which accounts for the majority (85%) of all lung cancers (Schabath and Cote, 2019).
The 5-year survival rate of patients with NSCLC remains at 23% (Siegel et al., 2019). To make it worse, patients with advanced NSCLC who are not suitable for surgery only gain an absolute survival benefit of 5.4% at 5 years from their routine treatment method, chemotherapy (Alexander et al., 2020; Pignon et al., 2008).
Remarkable progress has been made in the management of nonsurgical NSCLC with the advent of cancer immunotherapies (Howlader et al., 2020), and the most promising advance in immunotherapy is the incorporation of chimeric antigen receptor (CAR)-T cells (Ma et al., 2019). CAR-T cells refer to the T cells with genetically modified receptor (CAR) (Stadtmauer et al., 2020). T cells have 2 major T cell subtypes (CD8+ T cells and CD4+ T cells), both of which are important to the cytotoxicity induced by CAR-T cells (Golubovskaya and Wu, 2016).
CD8+ T cells mediate the adaptive immune responses with recognition and efficient cytolytic functions (Raskov et al., 2021), while CD4+ T cells can aid in the development of CD8+ T cell memory functions (Shedlock and Shen, 2003). With the addition of CAR, T cells are endowed with the ability to recognize more target antigens than just human leukocyte antigen (HLA), thus being able to kill more extensive cells when contrasted with normal T cells (Sadelain et al., 2003).
Nevertheless, the present effect of CAR-T cells is primarily witnessed in hematological malignancies, and targeting solid tumors with CAR-T cells remains a great challenge (Ma et al., 2019). This is due to the fact that the CAR-T cells transporting to and infiltrating into solid tumors are often limited by the immunosuppressive microenvironment (Zhang et al., 2016).
The type II transmembrane glycoprotein NKG2D is a C-type lectin-like receptor from the CD94/NKG2 family, which plays a regulatory role in both innate and adoptive immune responses (Dhar and Wu, 2018). NKG2D can be normally expressed on the surface of CD8+ T cells as well as natural killer (NK) cells, natural killer T cells, and γδ T cells (Raulet, 2003; Zhang et al., 2015). Meanwhile, NKG2D can also bind with its ligand (NKG2DL), thereby participating in the activation of NK cells, cytokine production, and lysis of target cells (Lanier, 2015).
NKG2DL is often induced by tumorigenic conditions while being generally absent on the surface of normal cells (Baragaño Raneros et al., 2014). Tumor cells' surface expression of NKG2DL enables NKG2D to activate NK cells and costimulate effector T cells, thus sensitizing these tumor cells to immune cell-mediated destruction (Baragaño Raneros et al., 2014). Currently, NKG2D-NKG2DL signaling has been reported as a promising target for antitumor therapy with CAR-T cells (Zhang et al., 2020).
Previous research has underlined that NKG2DL expression is higher in NSCLC than in SCLC (Zhu et al., 2021), hinting a possibility that NKG2D CAR-T cells are presented by NSCLC cells to bind to NKG2DL-expressing cancer cells and subsequently perform its cell-killing function.
In our research, accordingly, T cells from different sources were modified into NKG2D CAR-T cells, and then their killing effect was tested on NSCLC cells.
Methods and Materials
Ethical statement
The study has obtained the approval of the ethics committee of Lishui People's Hospital, the Sixth Affiliated Hospital of Wenzhou Medical University (Clinical approval No. 001-01, 2019), and all participants provided written informed consents to any experimental work with human specimen.
Clinical sample collection and T cell isolation
Peripheral blood was collected from healthy volunteers (n = 2), and peripheral blood, together with lymphoma tissues or with solid tumors, was also obtained from patients with lymphoma (n = 8) or patients with NSCLC (n = 8). After the collection, the peripheral blood and tumors were instantly cryopreserved in liquid nitrogen and stored in a refrigerator at −80℃.
The lymphoma tissues and solid tumors were ground by a sterile stainless-steel mesh, and digested to prepare single-cell suspension using 1 mg/mL collagenase type IV (C4-BIOC; Sigma-Aldrich, St. Louis, MO), 2 U/mL DNase I (11284932001; Sigma-Aldrich), and 0.5 U/mL hyaluronidase type V (H6254; Sigma-Aldrich). Then, the obtained single-cell suspension was filtered, after which the remnant cells were washed by phosphate-buffered saline (PBS; P4417; Sigma-Aldrich) and adjusted by lymphocyte serum-free medium (KBM581; Corning, Corning, NY) to become cell suspension with a density of 1 × 106 cells/mL (Wang et al., 2020).
Within 12 h after being stored in liquid nitrogen, primary T cells were isolated from the peripheral blood and the cell suspension through density gradient centrifugation on RosetteSep human T Cell enrichment cocktail (15021; Stemcell Technologies, Vancouver, Canada) (Meng et al., 2019). The isolated T cells were stored in a refrigerator at −80℃ before being used for CAR-T cell preparation.
NSCLC cell culture
Human NSCLC cell lines (A549, NCI-H1650, PC-9, H1299, and NCL-H460) were purchased from American Type Culture Collection (ATCC; CRM-CCL-185, CRL-5883, CRL-5803, and CRL-5835; Manassas, VA), and PC-9 cells were obtained from BLUEFBIO (BFN60800699, Shanghai, China). All these cells were cultured in RPMI-1640 medium (30-2001; ThermoFisher, Waltham, MA) with 10% fetal bovine serum (FBS; 12106C, Sigma-Aldrich) and 1% Penicillin–Streptomycin (P/S; 15140148, ThermoFisher). All these cells were incubated in a humidified incubator with 5% CO2 at 37°C.
In the meantime, 293 T cells were procured from Procells (CL-0005, Wuhan, China), and maintained in DMEM (11965092; ThermoFisher) containing 10% FBS and 1% P/S under the same condition as NSCLC cells.
Generation and package of lentivirus
A codon-optimized NKG2D extracellular region, along with a CD8 hinge, an intracellular domain of 4-1BB and a CD3zeta activation domain, was fused together to generate NKG2D CAR per the previous description (Gomes-Silva et al., 2017). The entire encoding sequence of the NKG2D CAR expression molecule was subcloned into LentiGuide-Puro vectors (#52963; Addgene, Watertown, MA).
Two hundred ninety-three T cells were used as the tool cells in lentivirus packaging. In brief, after being cultured for 24 h, 293 T cells were transfected with the CAR lentivirus vector and 2 helper vectors psPAX2 and pMD2.G (#12260 and #12259; Addgene), using PEIpro transfection reagent (101000026; Polyplus, Illkirch, France). The supernatant of the cultures was collected and concentrated after 48 h through ultracentrifugation, as described previously (Benskey and Manfredsson, 2016).
NKG2D CAR-T preparation
The isolated T cells were stimulated in 6-well plates precoated by immobilized CD3 and CD28 antibodies (customized by Elabscience, Wuhan, China) for 24 h, after which the activated T cells were incubated with the lentivirus (at a multiplicity of infection [MOI] of 5) for 16 h to produce CAR-T cells. The CAR-T cells were treated with 200 IU/mL interleukin (IL)-2 (SRP3085; Sigma-Aldrich) for future expansion in lymphocyte serum-free medium (Zhang et al., 2020).
The CAR-T cells from different sources of T cells were named H-T cells (T lymphocytes from peripheral blood of healthy volunteers), L-T cells (T lymphocytes from peripheral blood of patients with lymphoma), L-C-T cells (T lymphocytes from the lymphoma of patients with lymphoma), N-T cells (T lymphocytes from peripheral blood of patients with NSCLC), and N-C-T cells (T lymphocytes made from the solid tumor of patients with NSCLC). NT-T cells were the T lymphocytes from the peripheral blood of healthy volunteers (without lentivirus infection).
Flow cytometry
After 10-day IL-2 treatment, the CAR-T cells were harvested. Subsequently, NSCLC cells, NT-T cells, and the harvested CAR-T cells were centrifuged at 300 g for 3 min, and suspended in PBS to prepare cell suspension at a density of 1 × 106 cells/mL. The expression of NKG2DL in NSCLC cells was identified by being stained with the following antibodies: phosphoethanolamine (PE)-conjugated MHC class I polypeptide-related sequence A (MICA) antibody (FAB1300P; R&D Systems, Minneapolis, MN), Alexa Fluor 488-conjugated MHC class I polypeptide-related sequence B (MICB) antibody (FAB1599G; R&D Systems), and allophycocyanin (APC)-conjugated UL16 binding protein 1 (ULBP-1) antibody (FAB1380A; R&D Systems) for 30 min.
For the detection of NKG2D CAR, the suspension of NT-T and CAR-T cells was stained with PE-conjugated NKG2D antibody (FAB139P; R&D Systems) for 30 min. The stained cells were loaded onto a flow cytometer (FACSCanto II; Becton, Dickinson and Company, Franklin, NJ), and the data were analyzed by FlowJo 10 software (Becton, Dickinson and Company).
Cell coculture assay
CAR-T cells or NT-T cells (effector cells) were directly cocultured with PC-9 and NCL-H460 cells (1 × 104 cells, target cells) for 16 h in RPMI-1640 medium supplemented with 10% FBS and 1% P/S at the ratios of 0.5:1, 1:1, 2:1, 4:1, and 8:1 for cytotoxicity assay or of 8:1 for enzyme-linked immunosorbent assay (ELISA) (Meng et al., 2019).
Cytotoxicity assay
The supernatant (50 μL) of the cocultured cells was collected through centrifugation at 1,000 g for 5 min, and the level of lactate dehydrogenase (LDH) within the supernatant was detected by the LDH assay kit (ab102526; Abcam, Cambridge). Concretely, the supernatant was adjusted to the solution with a volume of 50 μL with LDH Assay Buffer, and then reacted with reaction mix. The optical density at a wavelength of 450 nm was measured every 2–3 min for ∼30 min using a microplate reader (PHERAstar FSX; BMG LABTECH, Offenburg, Germany) in a kinetic mode.
Murine xenograft assay
Male NOG mice (n = 45, 6 weeks old) were reared under the pathogen-free conditions in microisolator cages (temperature: 22°C ± 2.0°C, humidity: 50% ± 5%, and a 12-h dark–light cycle), and given free access to water and standard rodent chow.
Thereafter, these mice were used in murine xenograft assay. All assay protocols were implemented according to the guidelines of the National Institutes of Health on Animal Care and Use, and approved by the Ethics Committee of Zhejiang Baiyue Biotech Co., Ltd. for Experimental Animals Welfare (approval number: ZJBYLA-IACUC-20221028). PC-9 cells (4 × 106) were subcutaneously injected into the right flank of NOG mice.
Fourteen days later, the mice received the intravenous injection with 2 × 106 NT-T, H-T, or N-T cells (every 15 mice were injected with one type of CAR-T cells). The volume of the tumors was measured with calipers, and the mouse body was weighed every 10 days, starting from the day when NT-T or CAR-T cells were injected. The tumor volume was calculated as follows: volume (mm3) = 0.5 × length (mm) × width (mm)2.
In addition, the peripheral blood was collected from retro-orbital bleeding every 10 days, starting from day 0 after the injection of CAR-T cells, and used in ELISA. All mice were sacrificed through cervical dislocation under anesthesia using 1% pentobarbital sodium (P010, 50 mg/kg; Sigma-Aldrich). At days 3, 5, 7, and 10 post-CART injection, 12 of the mice were sacrificed each day, and mouse tumors were harvested for immunohistochemistry assay. The remaining mice were sacrificed at day 40 post-CART injection.
Enzyme-linked immunosorbent assay
The supernatant of the cocultured effector and target cells (at a ratio of 8:1) was collected as mentioned above. The mouse peripheral blood was centrifuged at 2,000 g for 20 min to obtain the serum as needed. The levels of cytokines, including tumor necrosis factor-(TNF)-α, IL-10, IL-2, and interferon (IFN)-γ, which were secreted by the above CAR-T cells into the coculture supernatant or into the mouse serum, were determined using the corresponding ELISA kits (AD20104, AD20148, AD20139, and AD20155 (human)/AD40155(mouse); ADANTI, Wuhan, China).
Simply put, the supernatant and the antibody cocktail (50 μL for each) were added into each well of precoated 96-well plates in sequence, followed by the incubation at room temperature for 1 h. After the plates were washed with washing buffer, TMB development solution (100 μL) was added to each well. After 10 min, the reaction was terminated by adding 100 μL stop solution to each well. The optical density at a wavelength of 450 nm was measured by the microplate reader after shaking of the plates for 1 min.
Immunohistochemistry assay
The tumors of mice were fixed with 4% paraformaldehyde (CL10571; Chemegen, Shanghai, China) for 24 h, dehydrated by gradient ethanol, transparentized using xylene (CL13001; Chemegen), and paraffin-embedded (A606115; Sangon, Shanghai, China). The paraffinized tumors were cut into 5-μm–thick slices, followed by the deparaffinization by xylene and rehydration using gradient ethanol. After rinsing in PBS, the slices were treated with 3% H2O2 (88597; Sigma-Aldrich) for 20 min.
After washing in PBS again, the slices were boiled in antigen-retrieval solution (P0088; Beyotime, Shanghai, China) for 5 min. PBS was employed once again for the rinse. Ten percent bovine serum albumin (A602449; Sangon) was used to block the slices for 30 min at room temperature. Subsequently, the slices were incubated with the primary antibody against CD3 (ab16669; Abcam) at 4℃ overnight. Secondary antibody goat antirabbit IgG-HRP (ab205718; Abcam) was employed to probe the slices.
Color development was performed utilizing Pierce DAB substrate kit (34002; ThermoFisher), and the counterstaining of the nuclei was conducted using hematoxylin (H8070; Solarbio, Beijing, China). Infiltration of CAR-T cells was observed by an inverted microscope (CKX53; Olympus, ToKyo, Japan) under × 100 magnification.
Statistical analysis
Statistical analysis was implemented with Graphpad prism (version 8.0; GraphPad Software, Inc., San Diego, CA). Data of results obtained from experiments conducted at least thrice were presented as mean ± standard deviation (SD). The survival periods of mice from different groups were presented in the Kaplan–Meier curves and compared by log-rank tests. Independent samples t-tests were implicated in comparison between the data of 2 groups, whereas comparison among multiple groups was carried out using one-way analysis of variance. A statistically significant difference was accepted when P < 0.05.
Results
NKG2DL was highly expressed in NSCLC cells, and NKG2D CAR-T cells from peripheral blood of healthy donors were more toxic to NSCLC cells than those from other T cell sources
The binding of NKG2D to NKG2DL expressed on cells to induce cell lysis is an impressive mechanism in cancer immunotherapy (Zhang et al., 2020). A relatively elevated expression of NKG2DL can be seen in NSCLC cells (Zhu et al., 2021). We therefore determined a potential immunotherapy designated to lyse NSCLC cells by activating NKG2D-NKG2DL signaling.
In the beginning, based on the data of flow cytometry, A549, NCI-H1650, PC-9, H1299, and NCL-H460 cells were all confirmed to specifically express NKG2DLs, MICA, MICB, and ULBP-1 on the cell surface (Fig. 1A). Then, we randomly selected out PC-9 and NCL-H460 cells as the subjects for the subsequent experiments. NKG2D CAR-T cells were designed, which contain the NKG2D extracellular domain, CD8α hinge and transmembrane domain, 4–1BB and CD3 zeta intracellular region, as illustrated in Fig. 1B.

NKG2DL was highly expressed in NSCLC cells, and NKG2D CAR-T cells from NSCLC solid tumors were more toxic to NSCLC cells than those from other T cell sources.
After separation from peripheral blood of healthy donors or peripheral blood/solid tumors of cancer patients, T cells were activated by immobilized CD3/CD28 antibody, and then underwent the infection with NKG2D CAR-lentivirus to generate CAR-T cells.
After 10-day IL-2 stimulation, surface expression of NKG2D was shown through detection on all these CAR-T cells (H-T, L-T, L-C-T, N-T, and N-C-T cells) using flow cytometry (Fig. 1C). Thereafter, H-T, L-T, L-C-T, N-T, and N-C-T cells as well as NT-T cells were cocultured with PC-9 and NCL-H460 cells at the ratio of 0.5:1, 1:1, 2:1, 4:1, and 8:1. All these CAR-T cells (H-T, L-T, L-C-T, N-T, and N-C-T), in comparison with NT-T cells, promoted the release of LDH from the cancer cells, and delivered the enhanced cytotoxicity to PC-9 and NCL-H460 cells; the H-T and N-T cells possessed the highest cytotoxicity in comparison with others (Fig. 1D–E, P < 0.001).
NKG2D CAR-T cells from peripheral blood of healthy donors were more potent at secreting cytokines TNF-α, IL-10, IL-2, and IFN-γ to kill NSCLC cells than those from other T cell sources
Moreover, cytotoxic T cells secrete cytokines upon killing target cells (Meng et al., 2019). The levels of the cytokines, including TNF-α, IL-10, IL-2, and IFN-γ, were pronouncedly higher in the supernatant of the PC-9 or NCL-H460 cells cocultured with the above CAR-T cells, in comparison with those in the supernatant of the PC-9 or NCL-H460 cells cocultured with NT-T cells (Fig. 2A–H, P < 0.001). The 2 kinds of CAR-T cells with the best anticancer effect (H-T and N-T cells) were chosen to be used in in vivo experiment.
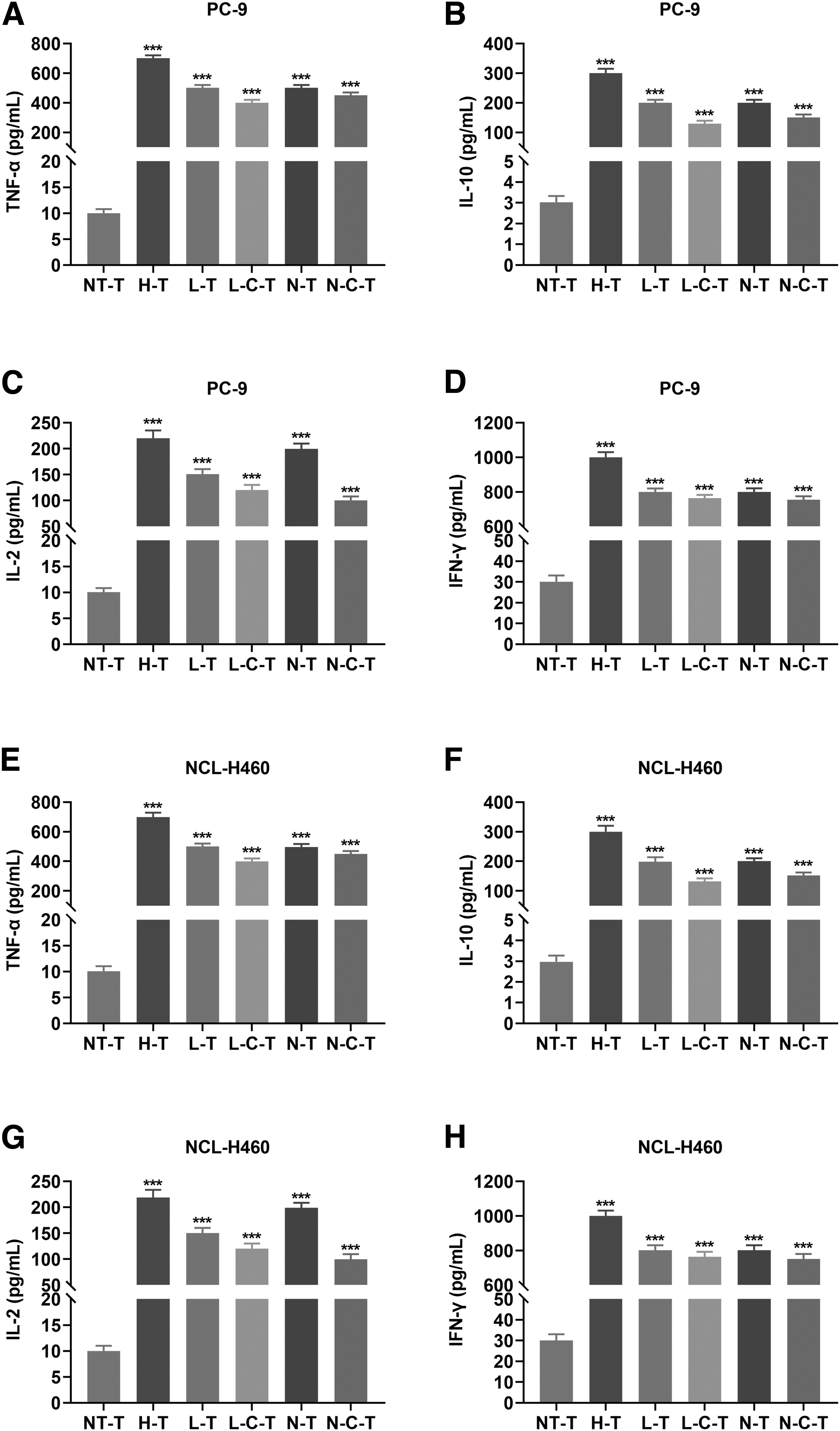
NKG2D CAR-T cells made from peripheral blood of healthy donors were more potent at secreting TNF-α, IL-10, IL-2, and IFN-γ cytokines that help kill NSCLC cells than those from other T cell sources.
NKG2D CAR-T cells from peripheral blood of healthy donors showed potent antitumor activity
In vivo experiment was conducted with xenografts in NOG mice using PC-9 cells to inspect the efficacy of tumor cytotoxicity induced by NKG2D-overexpressing CAR-T cells. Accordingly, PC-9 cells (4 × 106) were subcutaneously injected into right flank of NOG mice, and 14 days later, 2 × 106 of NT-T, H-T, or N-T cells were also injected through intravenous administration. The tumor volume and body weight of mice were measured, and the peripheral blood was collected every 10 days, starting from day 0 until day 40 post injection of NT-T/CART-T cells. The scheme is illustrated in Fig. 3A.

NKG2D CAR-T cells from peripheral blood of healthy donors showed potent antitumor activity.
When compared with NT-T cells, H-T and N-T cells decreased the volume of the transplanted tumors (Fig. 3B) and prolonged the survival time of the mice (Fig. 3C, P = 0.011). A higher serum level of IFN-γ was also detected in the mice at day 10 after the injection of these T cells (Fig. 3D, P < 0.001). In contrast with the treatment using NT-T cells, the treatment with H-T cells led to the weight gain of the mice from day 20 to day 40 post the treatment, and the treatment with N-T cells increased the weight between days 10 and 20 post-treatment (Fig. 3E, P < 0.001).
To further confirm the antitumor effects exerted by NKG2D CAR-T cells, the proportion of CD3+ cells was detected by immunohistochemical test with CD3 antibody at different time points when tumor-bearing mice received the injection of these CAR-T cells. As displayed in Fig. 3F, the significant infiltration of CD3+ cells and reduction of NKG2D+ cells were observed in the tumors at days 3, 5, 7, and 10 after the injection of H-T/N-T cell. Collectively speaking, the results conveyed the idea that NKG2D-overexpressing CAR-T cells deliver high-potential therapeutic efficacy for NSCLC patients.
Discussion
CAR-T cell therapy is a type of immunotherapy where the CAR-T cells can be used to lyse the target cells in a receptor–ligand interaction-dependent manner and be customized according to the ligand expressed on target cells (Sadelain et al., 2003). Previously, NKG2D CAR-T cells have been employed in the annihilation of cervical cancer cells (Zhang et al., 2020). NKG2D CAR-T cells, likewise, were applied in this study to treat NSCLC, and it was demonstrated in the corresponding data that NKG2D CAR-T cells from peripheral blood of healthy donors exerted the most remarkable killing effect on NSCLC cells.
NKG2D is a transmembrane protein serving as an activating receptor for CD8+ T cells (Lanier, 2015). Engagement of NKG2D by NKG2DL potentiates natural T cell surface receptor-dependent cytolytic and cytokine responses for CD8+ T cells (Groh et al., 2001). Moreover, NKG2D–NKG2DL interaction facilitates the immunorecognition of tumor cells, thus abrogating cancer progression (Baragaño Raneros et al., 2014).
CAR-T cells are capable of mediating HLA-unrestricted tumor cell-killing effects by enabling T cells to bind target cell surface antigens through CAR, which contains antibody driving specificity against the antigens (Benmebarek et al., 2019). NKG2D-NKG2DL signaling is frequently activated through CAR-T immunotherapy against cancer cells expressing high levels of NKG2DLs (Baumeister et al., 2019; Meng et al., 2019; Weiss et al., 2018). NKG2DLs are some stress-inducible ligands comprising family of MHC I chain-related molecules (MICA and MICB) and an array of cytomegalovirus UL16-binding proteins (ULBP1–6) (Eagle and Trowsdale, 2007; El-Gazzar et al., 2013).
Existing research has reported that compared with SCLC, NSCLC presented a higher expression level of NKG2DL (19). Similar results were also drawn in our study, as evidenced by the discovery that MICA, MICB, and ULBP-1 levels were unanimously upregulated on the cell surface of those tested NSCLC cell lines. In addition, patients with advanced NSCLC present an obviously low CD3+CD8+NKG2D expression rate in their lymphocyte subgroups than those with early lesions (Yu et al., 2014). The findings above suggested that CAR-T therapy, which resorts to achieving the activation of NKG2D-NKG2DL, may be a promising therapeutic method for NSCLC patients.
The core event before CAR-T therapy is the design of CAR. In our study, NKG2D CAR was designed according to the structure of the second generation of CAR (Golubovskaya and Wu, 2016), and generated as previously described (Zhang et al., 2020). It has been established that T cell sources are an element that must be reconsidered to improve the clinical effects of CAR-T cells (Huang et al., 2020). Therefore, NKG2D CAR was inserted into T cells from different autologous T cell sources, and the killing effects of the different types of NKG2D CAR-T cells were compared as appropriate.
Once CAR T cells are engaged by the ligand on target cells, a nonclassical immune synapse forms on the CAR T cells (Monks et al., 1998), enabling the CAR T cells to rapidly lyse and eliminate the target cells through fulfilling cytolytic effector functions, including the release of perforin and granzyme, the activation of Fas and Fas ligand pathway, and the secretion of cytokines to sensitize the tumor stroma and promote inflammatory (antitumoral) responses (Benmebarek et al., 2019). The release of perforin and granzyme by T cells causes the cytotoxicity (Voskoboinik et al., 2015), which can be reflected by the leakage of LDH (Fotakis and Timbrell, 2006).
Data of our in vitro experiment suggested that compared with NT-T cells, the LDH leakage from NSCLC cells was greatly aggregated by NKG2D CAR-T cells. The secretion of effector differentiation-related cytokines (eg, TNF-α, IL-2, and IFN-γ) by T cells antagonizes the effects of immunosuppressive factors, resulting in improved antitumor function of CAR-T cells, while the activation of IL-10 is associated with a memory phenotype of T cells (Chang et al., 2014; Martinez and Moon, 2019).
In our study, these aforementioned cytokines in the supernatant of these cocultured cells were all found to be secreted at a higher level by the above NKG2D CAR-T cells than by NT-T cells. T cells are sourced from the thymus, where they differentiate to be mature, and then circulate through blood into the thymus-dependent zone and the peripheral immune organs (Owen and Raff, 1970).
These T cells can further invade into tumor lesions to perform the antitumor tasks (Jiang et al., 2019). L-T and L-C-T cells from cancerous T cells show an abnormal immune function (Hosing and Champlin, 2011), and the tumor-derived T cells responsible for producing L-C-T and N-C-T cells exhibit a compromised antitumor ability, since the immunosuppressive tumor microenvironment favors the generation of immune suppressor cells, tumor-derived cytokines, and checkpoint inhibitory ligands, giving rise to decreased antitumor immunity conferred by T cells infiltrating tumor lesions and a weak response to CAR-T cells (Ma et al., 2019; Martinez and Moon, 2019). Therefore, we speculated that L-T, L-C-T, and N-C-T cells are less capable of killing tumor cells than CAR-T cells from T cells derived from normal peripheral immune organs (H-T and N-T).
As expected, our in vitro experiment data denoted that the top 2 most prominent cytotoxic and secretive effects were induced by H-T and N-T cells.
Although CAR-T therapy has made a significant breakthrough in eliminating hematological malignancies at present, obstacles still exist in the treatment of solid tumors with CAR-T cells. It is generally believed to be caused by the physical tumor barriers of solid tumors, which hinder CAR-T cells from trafficking to and infiltrating into the tumor. Meanwhile, the heterogeneity of solid tumors is higher than that of hematological malignancies, resulting in a lower antigen specificity in T cell therapy for solid tumors (Ma et al., 2019; Martinez and Moon, 2019).
Nevertheless, several studies have provided in vivo experimental data displaying that T cells expressing NKG2D CAR are effective in eradicating solid tumor xenografts formed by hepatocellular carcinoma cells (Meng et al., 2019), colorectal cancer cells (Ng et al., 2021), and cervical cancer cells (Zhang et al., 2020). Our study revealed that NKG2D CAR-T cells (H-T and N-T cells) exhibited their therapeutic effect on mice with NSCLC cell-formed xenografts through decreasing the xenograft volume, improving survival and increasing the body weight.
More importantly, after NKG2D CAR-T cell treatment, markedly increased serum level of IFN-γ as well as evident infiltration of the CAR-T cells into the xenografts was observed in these mice, suggesting that the efficacy of NKG2D CAR-T cells in treating NSCLC may be limited to a lesser extent than we had expected. Since patients with cancer are often immunosuppressed with depressed T cell function (Vallejo et al., 2003), the antitumor effect of H-T cells is anticipated to be stronger than that of NT cells, which is consistent with the result from our in vivo experiment. Thus, we considered H-T cells to be more ideal than N-T cells in NKG2D CAR-T therapy.
Conclusion
In conclusion, our study conveyed the idea that immunotherapy with NKG2D CAR-T cells is a promising method to treat NSCLC, and NKG2D CAR-T cells from healthy donors' peripheral blood exert the most potent tumor-killing effect, when compared with those from other T cell sources. However, proteolytic-mediated shedding or exosome-mediated secretion of NKG2DL from tumor cell surface would help tumor cells circumvent the immune surveillance of NKG2D activation, entering an antigen escape resistance pattern as the disease advances (Baragaño Raneros et al., 2014). Therefore, to maximize the efficiency of the therapy, NKG2D CAR-T immunotherapy should be administered as soon as possible after the diagnosis has been made and before the progression of tumor.
Authors' Contribution
J.J. has made substantial contributions to conception and design; Y.L., Y.Z., B.F., and Y.C. contributed to data acquisition, data analysis and interpretation; J.J. and Y.L. drafted the article or critically revised it for important intellectual content; all authors assisted with final approval of the version to be published; J.J., Y.L., Y.Z., B.F., and Y.C. agreed to be accountable for all aspects of the work in ensuring that questions related to the accuracy or integrity of the work are appropriately investigated and resolved.
Footnotes
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported by The Zhejiang Provincial Science and Technology Plan Project (Integrated development of autologous tumor cell vaccines with cell membrane surface anchoring modifications for 4th generation CART cell therapy) [2020ZJZC04].