
Abstract
Feline interferon omega (IFN-ω) has been proven to have high antiviral activity; however, its in-depth antiviral effects remain unknown. Extracellular vesicles (EVs) have been demonstrated to participate in the regulation of the immune response pathway for the body through various active substances, especially through the microRNA (miRNA) carried by them. In this study, we isolated EVs from feline peripheral blood by differential centrifugation, and further found that the content of IFN-ω in EVs increased continuously within 24 h after IFN-ω treatment, and a large number of miRNAs were significantly downregulated in EVs within 12 h after IFN-ω treatment. These significantly differentially expressed miRNAs were important for regulating changes in antiviral cytokines. This study reveals for the first time the correlation between EVs-mediated miRNA in feline peripheral blood and IFN-ω on antiviral immune response, which may provide strong data support for the development of novel antiviral nanomedicine and the research of the antiviral effects of IFN-ω.
Introduction
Feline interferon omega (IFN-ω) has a broad spectrum and high antiviral activity, including vesicular stomatitis virus (VSV), feline parvovirus, feline calicivirus, feline herpesvirus, etc., and has certain species specificity (Ritz et al., 2007; Yang et al., 2007). Further study on antiviral effects of IFN-ω may provide important theoretical basis for prevention and treatment of viral diseases in feline and other animals. However, since the discovery of IFN-ω in 1992, there has been lack of in-depth research on its mechanism of action.
Extracellular vesicles (Evs) are small vesicles that enclose biomolecules and range in size from 30 to 1,000 nm, and are composed of a double membrane structure derived from the cell membrane (Raposo and Stoorvogel, 2013). The biomolecules in EVs, protected by a phospholipid bilayer, are relatively stable from the influence of various enzymes in body fluids. EVs contain a variety of biomolecules such as lipids, proteins, and nucleic acids, which have the advantages of stability, easy preservation, high abundance, and easy collection (Kalluri and LeBleu, 2020; Raab-Traub and Dittmer, 2017; Raposo and Stoorvogel, 2013; Simons and Raposo, 2009).
Minimal Information for Studies of Extracellular Vesicles was proposed by the International Society for Extracellular Vesicles, including the isolation, properties, and functional studies of extracellular vesicles. It is proposed that EVs prepared by ultracentrifugation should be identified from at least three aspects: morphology, particle size, and specific labeled proteins.
So far, there have been a number of methods to classify EVs subsets, such as classifying EVs as nanovesicles, microvesicles, virus-like particles, EVs-like vesicles, and microparticles based on EVs size. Based on the mode of biogenesis, they are classified as EVs, membrane particles, outer membrane vesicles, and exfoliated membrane vesicles (Edwin van der et al., 2012; Raposo and Stahl, 2019; Tkach and Théry, 2016).
MicroRNA (miRNA) plays a crucial role in regulating cell proliferation, development, differentiation, apoptosis, metabolism, signal transduction, and immune response (Benatti et al., 2014; Lee et al., 2015; Xiao and Rajewsky, 2009). miRNA, a noncoding RNA of ∼22–24 nucleotides in length, is produced by genome coding. It was first identified in the nematode Caenorhabditis elegans in 1993 (Lee et al., 2019b). As an important regulatory factor, miRNA mainly regulates gene expression by complementary binding to the 3′UTR sequence of target genes.
After complementary binding to target genes, miRNA can inhibit translation or degrade target genes, which is related to the degree of complementarity between miRNA and target genes. When miRNA uses its seed region sequence (2–8 nt) to partially complement the 3′UTR of target genes, it induces translation inhibition of miRNA (Fabian and Sonenberg, 2012; Fabian et al., 2011).
MiRNAs contained in EVs regulate the antiviral immune response of host cells by controlling the expression of interferon (IFN) and induced antiviral substances (Sadri Nahand et al., 2020; Wang et al., 2020). It is worth noting that IFN or other biologically active substances in cells can also affect the expression profile of miRNAs in EVs. Host cells can transfer IFN-inducible miRNAs to EVs, thus amplifying the induction signal through EVs. For example, EVs derived from macrophages can transfer interferon alpha (IFN-α)-related miRNAs to hepatitis B virus (HBV)-infected liver cells (Wenyu et al., 2021).
In previous studies, we found significant differences in the expression profile of miRNAs in EVs secreted by cells infected with Seneca valley virus (Xu et al., 2020) and foot-and-mouth disease virus (FMDV; Yang et al., 2020). Moreover, miRNA-136 contained in EVs can inhibit the proliferation of FMDV.
In this study, we isolated EVs from feline peripheral blood treated with IFN-ω, and further found that the content of IFN-ω in EVs increased continuously within 24 h after IFN-ω treatment. In addition, a large number of miRNAs in EVs significantly decreased after IFN-ω treatment for 12 h, further demonstrating that these differentially expressed miRNAs are important regulators of changes in antiviral cytokines. This study for the first time revealed the role of EVs in mediating the antiviral immune response of feline involving IFN-ω and miRNA. This research is of great significance for the development of novel antiviral nanomedicines and the study of the antiviral effects of IFN-ω.
Materials and Methods
Cells, viruses, and IFN
Crandell Reese Feline Kidney (CRFK), F81, and 293t cells were obtained from the China Cell Resource Bank. The cells were maintained in Dulbecco's modified Eagle's medium supplemented with 2 mM
RNA extraction and quantitative polymerase chain reaction experiment
Total RNA was extracted from tissues and cells using the Trizol reagent (16096020; Thermo Fisher Scientific). Then, 5 μg RNA was reverse transcribed into cDNA using the reverse transcription kit (RR036A; Takara, Kyoto, Japan). SYBR Premix Ex Taq kit (Takara) and ABI Step One real-time polymerase chain reaction (RT-PCR) system (Applied Biosystems, Inc., Carlsbad, CA, USA) were used for RT-PCR of mRNA. The relative expressions of genes were calculated using the 2ΔΔCT method with GAPDH and β-actin mRNA as an internal control. The primer sequences are shown in Table 1.
Primers for Real-Time Quantitative Polymerase Chain Reaction Amplification
Extraction of EVs
To further isolate EVs from the feline's peripheral blood, we conducted the following centrifugation experiments. All centrifugation steps were carried out in a 4℃ environment. The peripheral blood was initially collected by centrifuging at 1000 g for 15 min to remove larger fragments and cells. The cell debris was further removed by centrifugation at 2,000 g for 30 min.
The resulting supernatant was centrifuged at 10,000 g for 45 min to remove the cells. The large vesicles were collected and filtered through a 0.22 μm filter. Finally, the collected supernatant was centrifuged at 120,000 g for 2 h using an ultracentrifuge (Thermo Scientific Sorvall WX100). The resulting precipitate was resuspended in 500 μL of PBS.
Nanoparticle tracking analysis
The EVs samples were diluted at a ratio of 1:1000 in PBS containing 0.05% Tween 20 in a total volume of 1.0 ml. Measurements were performed in triplicate using standard settings (refractive index = 1.331, viscosity = 0.89, and temperature = 25°C). Data analysis was performed using nanoparticle tracking analysis 3.2 software (Malvern Panalytical Ltd., Malvern, Worcestershire, UK), and the samples were evaluated using the Nanosight NS300 (Malvern Panalytical Ltd.).
Transmission electron microscopy
Direct morphological observation of the extracellular vesicles is crucial for extracellular vesicles identification. Therefore, we analyzed the extracted extracellular vesicles using transmission electron microscopy (TEM; Hitachi H-7000FA, Tokyo, Japan). After observation, we first extracted the extracellular vesicles using a TEM 200 copper mesh (EMS 80100-Cu US) followed by staining with phosphoric acid for 2 min. After drying under an incandescent lamp, TEM was used to observe the extracted EVs at a voltage of 80 kV.
Western blot analysis
For western blot (WB) analysis, the purified EVs were lysed with radioactive precipitation-free buffer (Santa Cruz Biotechnology, Dallas, TX, USA), and the cleared lysate was collected by centrifugation for protein separation on 12% sodium dodecyl sulfate-polyacrylamide gels. The separated proteins were transferred to 0.45 μm nitrocellulose (NC) filter membranes (Millipore, USA) after electrophoresis. The membranes were blocked for 1 h with Tris-buffered saline containing Tween 20 (TBST) with 5% nonfat milk.
The blots were then incubated with primary antibody at 4°C overnight. The primary antibodies used included rabbit monoclonal anti-CD9 (Abcam, Cambridge, UK) and anti-CD81 (Abcam). After washing three times with TBST, the membranes were incubated with horseradish peroxidase-labeled secondary antibody (Proteintech, Chicago, IL, USA) for 2 h at room temperature. Finally, the proteins were visualized with Clarity Enhanced Chemiluminescence WB substrate (Bio-Rad Laboratories, Hercules, CA, USA).
Laser scanning confocal microscopy
The extracted EVs were stained with the dye PKH67 (Sigma, Aldrich, USA), following the instructions provided. The CRFK and F81 cells were seeded into a 30 mm laser-confocal cell culture dish and when the cells had grown to 80% of the dish's surface area, the EVs were inoculated into the cells. After inoculation with EVs for 12 h, the cells were harvested, fixed with 4% paraformaldehyde, and stained with DiI and DAPI for membrane and nucleus, respectively. The entry of EVs into the cells was observed using a laser confocal microscope (Zeiss LSM 800).
Analysis of differential miRNA target gene functional enrichment in EVs
Gene ontology (GO) enrichment analysis provides all GO terms that significantly enriched in Differential Expression Genes (DEGs) compared with the genome background, and filter the DEGs that correspond to biological functions. First, all DEGs were mapped to GO terms in the Gene Ontology database (
Here N is the number of all genes with GO annotation; n is the number of DEGs in N; M is the number of all genes that are annotated to the certain GO terms; m is the number of DEGs in M. The calculated P-value had passed FDR correction, taking False discovery rate (FDR) ≤0.05 as a threshold. GO terms meeting this condition were defined as significantly enriched GO terms in DEGs. This analysis was able to recognize the main biological functions that DEGs exercise.
Kyoto Encyclopedia of Genes and Genomes (KEGG) is the major public pathway-related database (Xiao and Rajewsky, 2009). Pathway enrichment analysis identified significantly enriched metabolic pathways or signal transduction pathways in DEGs compared with the whole-genome background. The calculated formula is the same as that observed in GO analysis.
Results
Detection of antiviral activity of IFN-ω IFN and its induced antiviral cytokines
Feline IFN-ω has a strong antiviral activity compared with other feline IFNs, and it has certain species specificity. However, the in-depth mechanism of IFN-ω regulating the body's immune response is still unknown. Studies have shown that EVs mediate bioactive substances to participate in the body's immune response. Whether EVs participate in the body's immune response by mediating IFN-ω is still unknown. This study analyzed the antiviral activity units of three IFNs (IFN-ω, IFN-α, and interferon gamma [IFN-γ]) against VSV in feline using the REED MECH method (Xin-Qin et al., 2020).
The results showed that the antiviral activity unit of IFN-ω was 24.43 million units, while the antiviral activity units of IFN-α and IFN-γ in feline were 1.66 million and 5.00 million units, respectively (Supplementary Data S1). 293t cells were inoculated at the same protein concentrations of IFN-ω, IFN-α, and IFN-γ, respectively, and the results showed that IFN-ω had significantly higher induction levels of ISG15 and ISG56 antiviral factors in 293t cells than IFN-α and IFN-γ (Fig. 1A). This study demonstrated that feline IFN-ω has a high antiviral activity and induces significantly higher levels of antiviral cytokines than other feline IFBs. However, the mechanism by which feline IFN-ω significantly upregulates the expression of ISG15 and ISG56 remains unknown.
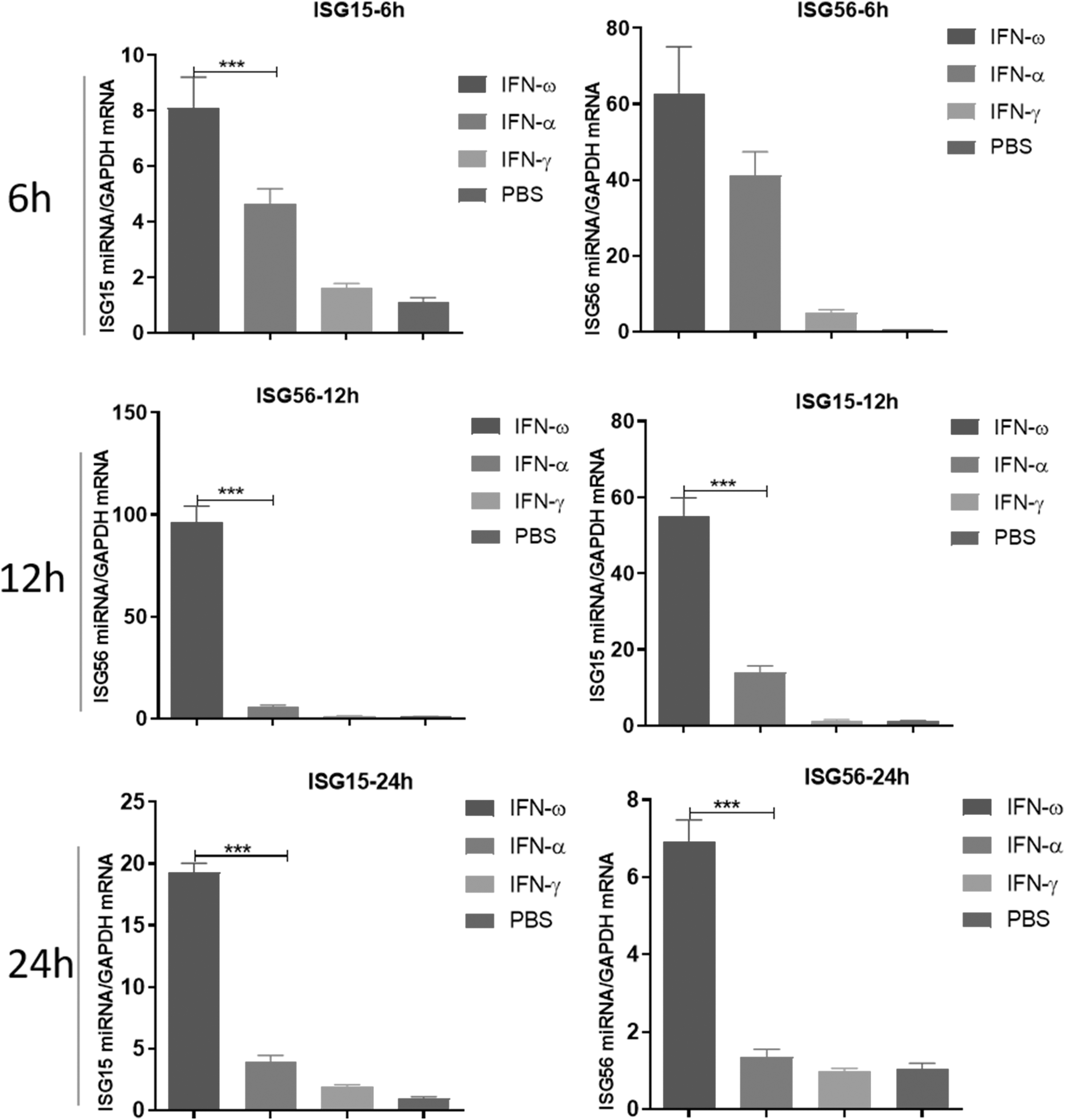
IFN-ω interferon antiviral activity and its induced detection of antiviral cytokines: IFN-ω, FN-α, and IFN-γ with the same protein concentration were inoculated into 293t cells, respectively, and then cultured for 6, 12, and 24 h, collecting cells. Total RNA was extracted by Trizol method, and the expression levels of ISG15 and ISG56 cytokines in cells were detected by RT-qPCR. All data were presented as mean ± SEM, n = 3, for each group. Significant differences were calculated using a two-tailed t-test, and ***P < 0.001 were marked on the graph. RT-qPCR, real-time quantitative polymerase chain reaction; SEM, standard error of the mean.
Extraction and identification of EVs from IFN-ω–treated feline peripheral blood
Recent research has shown that EVs mediate the participation of biologically active substances in the body's immune response and play an important regulatory role. It remains unknown whether EVs facilitate the immune response by mediating the expression of IFN-ω. To this end, we further extracted EVs from peripheral blood of IFN-ω–treated feline and performed identification analysis. The peripheral blood samples were collected from feline at 0, 12, and 24 h after IFN-ω injection.
Total RNA was extracted from serum using Trizol method, and then the expression levels of ISG15, ISG56, and Mx2 in the peripheral blood were detected by RT-qPCR. The results showed that the transcription levels of ISG15, ISG56, and Mx2 in the peripheral blood significantly increased at 12 h after IFN-ω injection, indicating that IFN-ω can induce the highest level of antiviral cytokine expression at 12 h after injection (Fig. 2A).
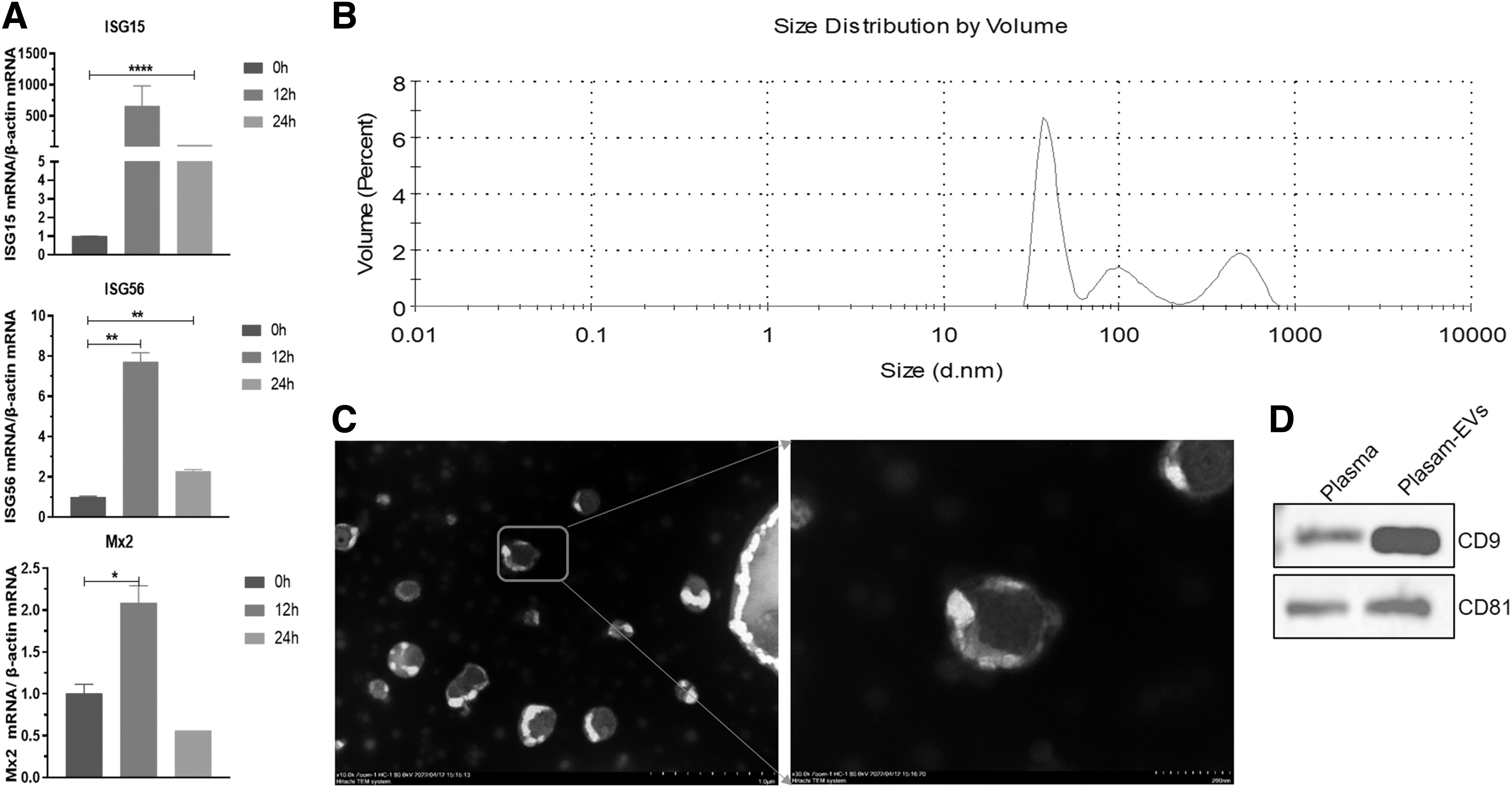
Extraction and identification of feline plasma EVs in IFN-ω–treated feline:
We then further extracted EVs from feline peripheral blood, identified them using nanoparticle size analyzers, transmission electron microscopes, and Western blot experiments, and found that EVs mainly distributed in the range of 50–800 nm in feline peripheral blood (Fig. 2B). Under TEM, numerous cup-shaped vesicle structures of different sizes could be observed (Fig. 3C). The marker proteins CD9 and CD81 of EVs could be specifically detected (Fig. 2D).
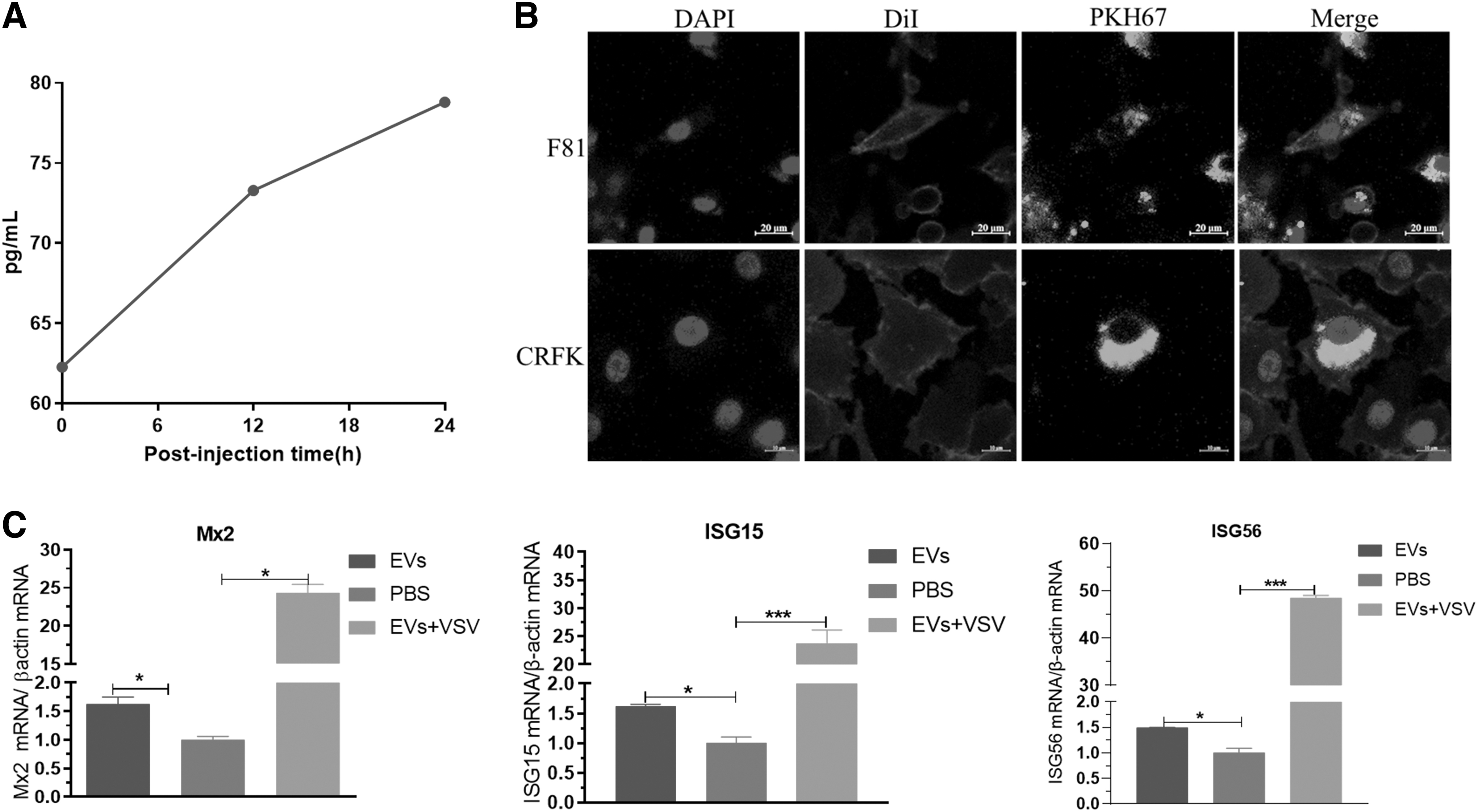
Feline EVs mediate IFN-ω–induced expression of antiviral cytokines:
EVs mediate IFN-ω–induced antiviral cytokine expression
To further analyze whether EVs mediate the spread of IFN-ω and their functional role, this study further detected the content of IFN-ω carried by EVs in the peripheral blood of feline injected with IFN-ω at 0, 12, and 24 h by ELISA. The results showed that the content of IFN-ω carried by EVs in feline injected with IFN-ω increased continuously at 0, 12, and 24 h after injection (Fig. 3A). To further analyze the function of EVs, we extracted EVs and inoculated them into CRFK and F81 cells.
At 12 h after inoculation, the EVs could be seen entering CRFK and F81 cells (Fig. 3B). Interestingly, EVs entering F81 cells significantly upregulated the expression of antiviral cytokines such as ISG15, ISG56, and Mx2. This study suggests that not only does the peripheral blood of feline injected with IFN-ω carry EVs, but also the expression of antiviral cytokines is significantly upregulated (Fig. 3D). However, it is still unknown whether miRNA carried by EVs participates in the regulation of antiviral cytokine expression.
Functional enrichment analysis of differential miRNAs and their target genes in EVs
EVs can carry multiple bioactive substances, including proteins, RNA, and lipid nanoparticles. Among them, miRNA carried by EVs play an important role in the function of EVs. We further analyzed the miRNA expression profile in the peripheral blood EVs of feline injected with IFN-ω, and the results showed that EVs contained a variety of RNAs such as cell genome, tRNA, rRNA, and miRNA, among which the number of miRNAs was the highest (Fig. 4A).
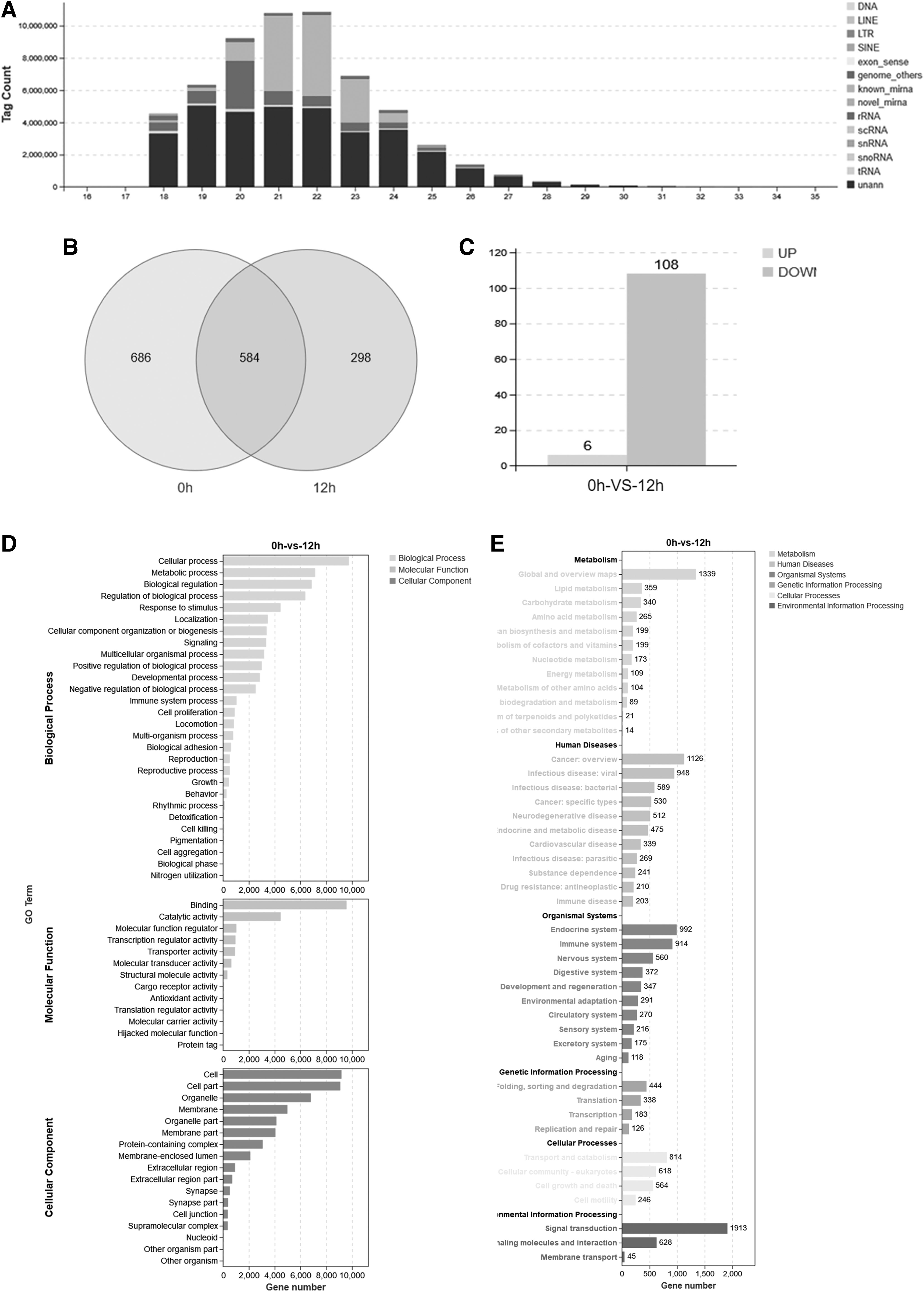
Functional enrichment analysis of miRNA target genes in feline EVs:
After injection of IFN-ω for 12 h, the number of miRNA species in EVs dropped from 1,270 to 882, and 298 of these were unique (Fig. 4B). Among them, 108 miRNAs showed significant downregulation, while 6 miRNAs showed significant upregulation (Fig. 4C). The GO functional analysis of differentially expressed miRNAs revealed that target genes involved in biological processes such as cellular process, metabolic process, biological regulation, and regulation of biological process were enriched.
The main molecular functions of target genes were binding, catalytic activity, molecular function regulator, and transcription regulator activity. Cellular components involved in target genes included cell parts, cells, and organelle parts (Fig. 4D). KEGG pathway analysis revealed that there were 914 immune system targets regulated by differentially expressed miRNAs in organismal systems, which accounted for higher proportion of all regulatory pathways in organismal systems.
In addition, when analyzing the environmental information processing involved in target genes, it was found that there were 1,913 cellular signal transduction pathways involved in target genes, which also accounted for the highest proportion of all regulatory pathways (Fig. 4E). These findings suggest that differentially expressed miRNA target genes play an important role in regulating the immune system and signal transduction in feline.
Functional enrichment analysis of differential miRNAs and their target genes in feline EVs
The above study suggests that miRNAs in EVs have regulatory effects on the immune system. To further screen for miRNAs that regulate the expression of antiviral cytokines such as ISG15, ISG56, and Mx2 from EVs, we analyzed differentially expressed miRNAs. The results showed that there were 20 miRNAs with extremely significant changes (Fig. 5A). We transfected miRNA-122-y, miRNA-127-y, miRNA-128-y, miRNA-32-y, and miRNA-1298-y mimics into F81 cells.

Screening and analysis of miRNAs regulating antiviral cytokine expression in feline peripheral blood EVs:
The results showed that these five miRNAs all regulated the expression of ISG15, ISG56, and Mx2 antiviral cytokines to varying degrees induced by VSV. Among them, miRNA-122-y and miRNA-128-y still significantly upregulated the expression of ISG15 and Mx2 without VSV induction. miRNA-132-y still significantly upregulated the expression of these three antiviral cytokines without VSV induction (Fig. 5B). This study suggests that these five differentially expressed miRNAs have different levels of regulatory effects on the expression of antiviral cytokines.
Discussion
IFNs are classified into three types (Type I, Type II, and Type III) (Niewold, 2020). Type I IFN can be further divided into IFN-α, IFN-ω, and Interferon beta (IFN-β) based on their antigenicity for binding to antibodies. IFN-ω plays an important regulatory role in the antiviral immune response of feline during viral infections (Leal et al., 2015), and it is currently the most commonly used for treating various infectious diseases in feline. In this study, we also demonstrated that the antiviral activity units of feline IFN-ω were significantly higher than those of IFN-α and IFN-γ.
Our research showed that both IFN-ω and IFN-α participate in downstream regulation of antiviral cytokine expression through the classic pathway of type I IFNs (Ivashkiv and Donlin, 2014; Ma et al., 2014; Platanias, 2005). However, our study revealed a significant difference in the antiviral activity units between IFN-ω and IFN-α, suggesting that there may be a significant difference in the regulatory pathways between these two IFNs. Human Type I IFN-α has been shown to induce macrophage secretion of EVs with anti-HBV activity, and EVs mediate the spread of antiviral agents between cells (Li et al., 2013). In this study, we found for the first time that EVs are capable of carrying out this function in feline peripheral blood.
We found that the expression levels of ISG15, ISG56, and Mx2 in peripheral blood increased to their highest levels at 12 h after injection of IFN-ω in feline. It is noteworthy that a large amount of IFN-ω was also present in EVs in peripheral blood at this time point, and the content of IFN-ω in EVs gradually increased over the next 24 h. Further studies showed that EVs could enter cells at 12 h after incubation and significantly induce the expression of ISG15, ISG56, and Mx2.
Unfortunately, due to experimental limitations in this study, we only detected the content of IFN-ω in EVs and did not examine the content of other antiviral cytokines carried by EVs. Our results suggest that EVs may play a role in regulating antiviral immune responses by mediating the delivery of IFN-ω and other antiviral active substances induced by IFN-ω.
IFN-ω plays an important role in regulating the transcription of miRNAs (Cheng et al., 2017; Chiantore et al., 2017; Yang et al., 2015), and EVs can mediate the involvement of various proteins and miRNAs in intercellular signal transmission. It is important to note that they can specifically enrich miRNAs to participate in the regulation of the immune response pathway of the body (Lee et al., 2019a; Liu et al., 2019; Munir et al., 2020). Due to the presence of other bioactive substances in EVs, it is not certain that the regulation of antiviral cytokines by EVs comes solely from IFN-ω in EVs.
Our further analysis of the expression profile of miRNAs in EVs also confirmed this conclusion. In our study, we found that after IFN-ω treatment, there were 108 miRNAs significantly downregulated and only 6 upregulated in EVs, indicating that IFN-ω has a significant downregulation effect on the expression profile of miRNAs in EVs. The target gene functions of these differentially expressed miRNAs are mainly related to signal transduction and immune regulation.
We further demonstrated that five selected differentially expressed miRNA mimics could significantly upregulate the expression of ISG15, ISG56, and Mx2 in F81cells. We speculated that the decrease in the number of miRNAs in EVs may be related to the increase in the usage of miRNAs in cells after IFN-ω activates a certain signaling pathway in cells, resulting in a decrease in the number of miRNAs entering EVs. However, according to our current results, it is still impossible to prove this conclusion.
Although our study still has many limitations, it has proven that IFN-ω not only has an important impact on the expression profile of miRNAs in EVs but also indirectly upregulates the expression of antiviral cytokines through miRNAs. Our study suggests that EVs participate in the regulation of the expression of antiviral cytokines in feline after IFN-ω treatment, and the function of EVs is significantly influenced by the active substances contained in them, such as IFN-ω and miRNA.
Conclusion
In this study, we isolated EVs from feline peripheral blood by differential centrifugation, and further found that the content of IFN-ω in EVs increased continuously within 24 h after IFN-ω treatment, and a large number of miRNAs were significantly downregulated in EVs within 12 h after IFN-ω treatment. These significantly differentially expressed miRNAs were important for regulating changes in antiviral cytokines.
Footnotes
Acknowledgments
The authors thank all the editors and reviewers for their valuable comments and suggestions that helped improve the article. They thank Innovation Laboratory of Chinese Veterinary Medicine, Lanzhou Institute of Animal Husbandry and Veterinary Medicine, Chinese Academy of Agricultural Sciences and State Key Laboratory of Veterinary Etiological Biology, National Foot-and-Mouth Disease Reference Laboratory, Lanzhou Veterinary Research Institute, Chinese Academy of Agriculture Science for their support of this article. They express their sincere gratitude to
Authors' Contributions
X.W. and Y.S. contributed to conceptualization; M.Y. performed methodology; J.Z. provided software; Z.G. performed formal analysis; C.L. conducted investigation; Y.L. provided resources; L.W. performed data curation; G.X. assisted with writing—review and editing; J.L. and T.G. provided supervision; X.W. and G.X. contributed to project administration; X.W. and G.X. performed funding acquisition; all authors read and approved the final article.
Date Availability Statement
The datasets generated from this study are not publicly available as other studies based on these data are ongoing. The data will be made available by the corresponding author on reasonable request.
Institutional Review Board Statement
The ethical policies of the journal, as noted on the journal's author guidelines page, have been adhered to. No ethical approval was required for sampling of the involved animals.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported by the Gansu Province Youth Science and Technology Fund program (Grant No. 21JR7RA035), Lanzhou Talent Innovation Industry Project (Grant No. 2022-RC-46), Fundamental Research Funds of the Chinese Academy of Agricultural Sciences (Grant No. 1610032021017), and Fundamental research fees of Chinese Academy of Agricultural Sciences (Grant No. Y2022XK19).
Supplementary Material
Supplementary Data S1